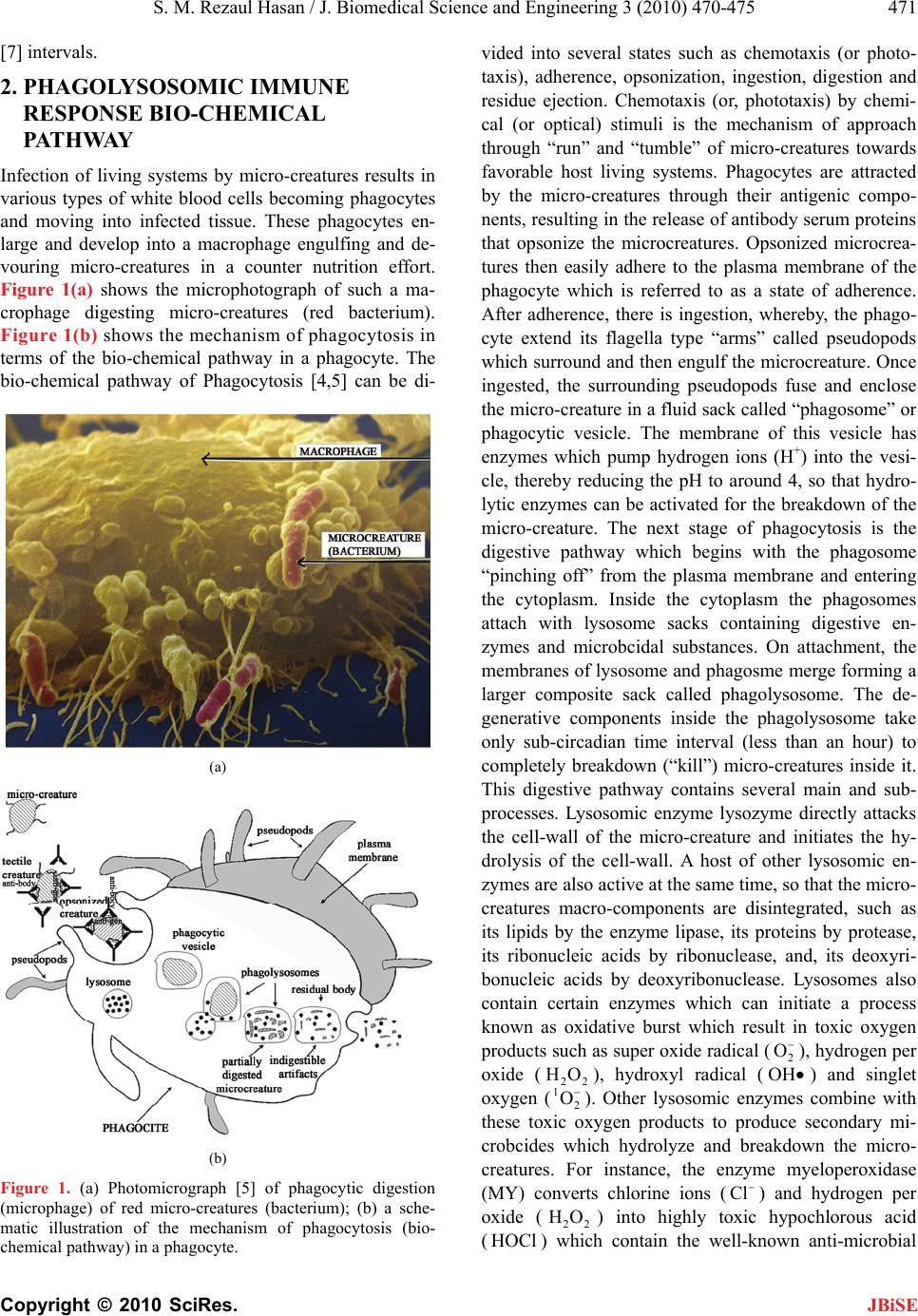
S. M. Rezaul Hasan / J. Biomedical Science and Engineering 3 (2010) 470-475 475
Copyright © 2010 SciRes. JBiSE
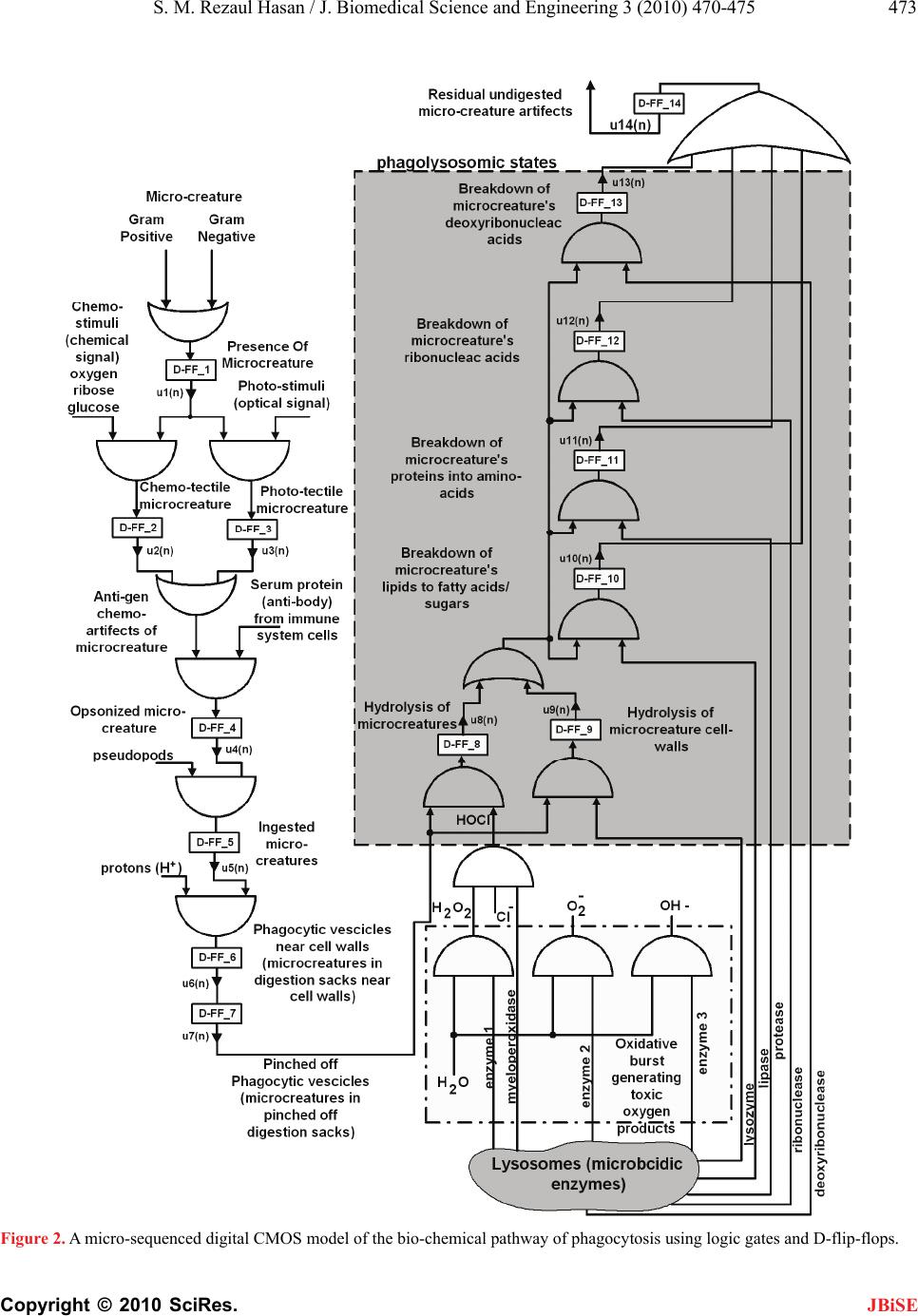
causes the immune response pathway to activate. This
results in the presence of either chemo-tectile (D-FF_2)
or photo-tectile (D-FF_3) micro-creatures during the
circadian interval <2,9>. Also, opsonized microcreatures
are present during the interval <3,10>, microcreatures
are ingested during the interval <4,11>, digestive vesi-
cles are formed near the phagocyte membrane during the
interval <5,12>, vesicles are detached and moves into
the cytoplasm during the interval <6,13>, phagolyso-
some is formed and the microcreature is hydrolyzed
(state in D-FF_8) and microcreature cell-wall is hydro-
lyzed (state in D-FF_9) during the interval <7,14>. The
breakdown of lipids (state in D-FF_10), proteins (state in
D-FF_11), ribonucleic acid (state in D-FF-12) and de-
oxyribonucleic acid (state in D-FF_13) take place simul-
taneously in the interval <8,15>. The undigested mi-
cro-creature artifacts (state in D-FF_14) are removed
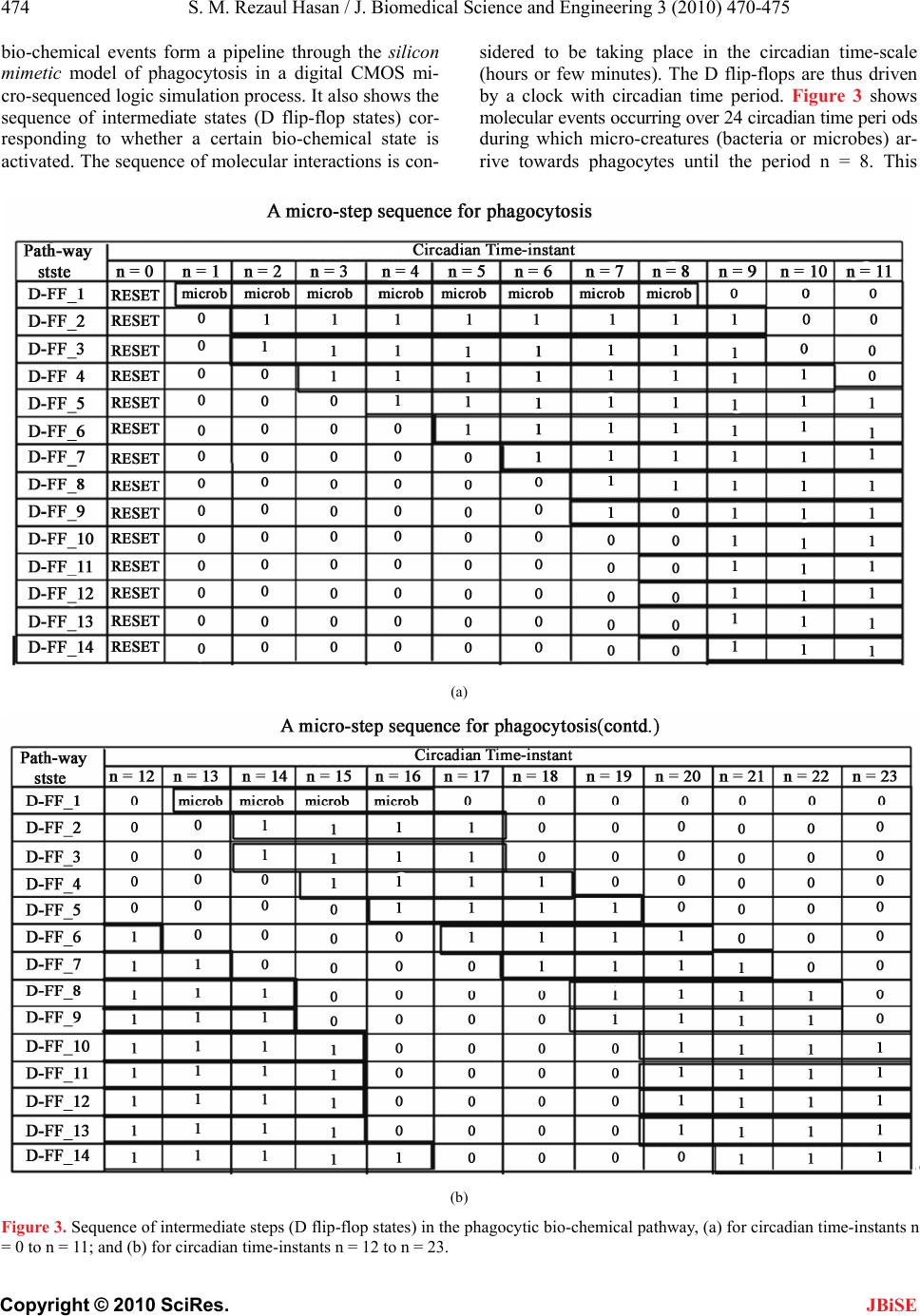
during the interval <9,16>. As the micro-creatures are is
not present during the interval <9,12>, there is a short
pause in the digestive bio-chemical process which is
evident from the diagonal zero states in Figure 3(b).
Also, beginning the interval <13,16> the presence of a
short burst of microcreatures results in a short pipeline
of digestive pathway activities (reactions and interac-
tions) that is evident through the diagonal array of “1” as
shown in Figure 3(b).
5. CONCLUSIONS
A digital circuit based model for the phagocytic bio-
chemical digestive pathway in living systems has been
developed and discussed in detail. The model corre-
sponds quite well with the immune response phenome-
non depicting striking resemblance of CMOS logic cir-
cuit (with states in D flip-flops) to states in bio-cellular
phenomenon. Compared to mathematical modeling,
model derived from analogies with integrated circuit
allows VLSI CAD circuit and logic simulators to be con-
veniently used as a biological simulation program.
Hence this work provides an alternative route for further
systems biological investigation into more comprehen-
sive integrated circuit models for more extensive bio-
chemical pathways in living systems. This investigation
will thus contribute to the desired manipulation of bio-
logical processes at the cellular level leading to electrical
circuit modeling of diseases and “nano-medicine”.
REFERENCES
[1] Dhar, P.K., Zhu, H. and Mishra, S.K. (2004) Computa-
tional approach to systems biology: From fraction to in-
tegration and beyond. IEEE Transactions on Nanobio-
science, 3(3), 144-152.
[2] Khammash, M. and El-Samad, H. (2004) Systems boi-
logy: From physiology to gene regulation. IEEE Control
Systems Magazine, 24(4), 62-76.
[3] Vaidyanathan, P.P. (2004) Genomics and proteomics: A
signal processor’s tour. IEEE Circuits and Systems Mag-
azine, 4(4), 6-29.
[4] Campbell, N.A. and Reece, J.B. (2005) Biology. Pearson,
San Francisco.
[5] Tortora, G.J., Funke, B.R. and Case, C.L. (2007) Micro-
biology: An introduction. Pearson Benjamin Cummings,
San Francisco.
[6] Watson, J.D. and Crick, F.H.C. (1953) A structure for
DNA. Nature, 171(4356), 737-738.
[7] Goldbeter, A. (1995) A model for circadian oscillations in
the drosophila period protein. Proceedings of Biological
Sciences, 261(1362), 319-324.
[8] Chen, L. and Wang, R. (2006) Designing gene regulatory
networks with specified functions. IEEE Transactions on
Circuits and Systems-I: Regular Papers, 53(11), 2444-
2450.
[9] Hasan, S.M.R. (2005) A novel CMOS integrated circuit
model for cellular DNA-protein regulatory mRNA tran-
scription process. Proceedings of 12th International Con-
ference on Biomedical Engineering, International Federa-
tion for Medical and Biological Engineering CD proceed-
ings, Singapore.
[10] Rezaul Hasan, S.M. (2005) A novel integrated circuit
model for mRNA transcription in bio-cellular processes,”
Proceedings of 12th Electronics New Zealand Confer-
ence, Manukau, 7-12.
[11] Hasan, S.M.R. (2008) A novel mixed-signal integrated
circuit model for DNA-protein regulatory genetic circuits
and genetic state machines. IEEE Transactions on Cir-
cuits and Systems-I: Regular Papers, 55(5), 1185-1196.
[12] Hasty, J., McMillen, D. and Collins, J.J. (2002) Engi-
neered gene circuits. Nature, 420(14), 224-230.
[13] Simpson, M.L., Cox, C.D., Peterson, G.D. and Sayler,
G.S. (2004) Engineering in the biological substrate: In-
formation processing in genetic circuits. Proceedings of
the IEEE, 92(5), 848-863.
[14] Hasan, S.M.R. and Ula, N. (2008) Analog CMOS charge
model for molecular redox electron-transfer reactions
and bio-chemical pathways. Proceedings of International
Symposium on Circuits and Systems, Geneva.
[15] Rezaul Hasan, S.M. (2008) A Micro-sequenced CMOS
model for cell signaling pathway using G-protein and
phosphorylation cascade. International Journal of Intelligent
Systems Technologies and Applications, 19(12) 57-62.
[16] Schiek, R.L. and May, E.E. (2003) Development of a
massively-parallel, biological circuit simulator. Proceed-
ings of the Computational Systems Bioinformatics, Cam-
bridge, 620-622.
[17] K. Martin, (2000) Digital integrated circuit design. Ox-
ford University Press, Oxford.