 Open Journal of Forestry 2012. Vol.2, No.1, 9-16 Published Online January 2012 in SciRes (http://www.SciRP.org/journal/ojf) http://dx.doi.org/10.4236/ojf.2012.21002 Copyright © 2012 SciRes. 9 Analysis of Pseudoreplicants to Evaluate Natural Regeneration after Applying Prescribed Burns in a Temperate Forest of Mexico José Germán F l ore s G arnica Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias, Guadalajara, México Email: flores.german@ inifa p.gob.mx Received August 19th, 2011; revised J an ua ry 3rd, 2012; accepted January 10th, 2012 Although fire is one of the most important disturbing factors of forest in Mexico, little it is known on the effects of fire on the particular Mexican forest ecosystems. This is remarked for the fact that the effects of fires on vegetation vary among different types of forests. This lack of knowledge has constrained the use of fire, as a silvicultural tool. Therefore, the purpose of this project was to evaluate the effects of fire on regeneration, under burns. This work was carried out in a pine forest stand at Tapalpa Saw in Jalisco State, Mexico, dominated by Pinus michoacana and Pinus oocarpa. The study evaluated the effects of two techniques of prescribed burning: 1) backing, and 2) head fire. The sample plots were burned on 25 and 26 March 1991, before the rain season. One month before and 2 years after burning several measurements were made in order to evaluate the effect of fire on regeneration. Due to the limitations to work with “real” replicates (for treatments an control), original sample units (20 × 30 m) were divided into 5 × 5 m smaller sample units, which were considered as pseudoreplicants. Therefore, such analysis did not avoid introducing systematic error (bias) and minimize random error. Nevertheless, the variability within the pseudoreplicants was considerable in order to assume certain significance of the resulting estimations. Therefore, despite that this was a nonreplicated study; the results suggest strong ecological evidence that prescribed fire enhance natural regeneration of Pinus michoacana and Pinus oocarpa. In general, it is concluded that prescribed burning could be a valuable forest management tool in regions with similar conditions to the study area, in order to improve regeneration. However, further research is needed before prescribed fires can be applied with confidence in many Mexican forest conditions. Keywords: Nonreplicated Study; Fire Effect Pinus michoacana; Pinus oocarpa; Forest Fires Introduction The successful mixing of fire and regeneration of some pine species is not just a coincidence (Jenkins et al., 2011; VanLear & Waldrop, 1991). As an example, there are serotinous trees, which release a limited amount of seed year to year that need of fire for general seed release (Vega et al., 2008; Enright & La- mont, 1989). These trees needs to be exposed to temperatures above 50˚C, as during a prescribed fire, in order that resin bonds of cones break and release large quantities of seed for dis- persal and reproductive growth if mineral soil is exposed (Teich, 1970; Givnish, 1981). In this way, serotiny should be a disad- vantage where fires are less frequent (Givnish, 1981) Moreover, some conifers that become established most readily on bare mineral soil reproduce poorly because litter covers favorable seedbeds (Land & Rieske, 2006; Smith, 1986; Johnston, 1971). Thus, eliminating or decreasing litter of forest floor, through prescribed fires, will result in better conditions for natural re- generation (Arno, 1980). Moreover, through prescribed fires, nitrogen and other nutrients of dead vegetation are released into soil, which result in a improving of the seedbed (Haywood, 2007; Wade, 1989; DeBano, 1976). This favorable response of regeneration to such temporal improved conditions corresponds to “the law of population growth with limiting factor” (Ramade, 1984). Therefore, forest managers have tried to emulate the natural advantages of fire to improve natural regeneration (Nesmith et al., 2011; McNabb, 2001). Although the use of fire, as a forest management tool, has been practicing in many countries with similar conditions to Mexico (Wells et al., 1979; Aguirre, 1981; Hudson & Salazar, 1981), the use of prescribed fires in Mexico is very restricted (Flores, 2001; Toledo, 1988). This is due mainly to two major facts: 1) most of the forest managers do not have a real knowledge of the methodology and techniques that exist behind the use of fire; and 2) very little is known about the effects of prescribed fires in the particular conditions of the Mexican forests. Therefore, wi t h th e i dea of contributing to knowledge about fire effects, the purpose of this paper is to evaluate the impact of two tech- niques of prescribed fires on regeneration. As it is known, statistically, we require of some replications in our experimental design, in order to support the significance of the difference among some given treatments (William, 1992). Also they are necessary in order to check that our results have certain consistency and to estimate the experimental error. This improves our precision by reducing the variation (standard deviation) of a treatment mean. Moreover, the use of replica- tions could help us to define if we have to estimate mo re var iable experimental units. With the purpose of avoids introducing sys- tematic error (bias) and minimize random error (types I and II errors) (Heffner et al., 1996; Hurlbert, 1984). However, in this study, the implementation of such replicates was time and cost consuming, and, more relevant, risky. This limited the chance to implement an orthodox experimental design in this study. Therefore, alternatively, I divided out the original sample units into smaller sample units, which were considered as “repli- cants”. However, these were more properly considered as “pseu-  J. G. F. GARNICA do replicants”. Which are used freque ntly in many ecological field experiments, though not always consciously (Hurlbert, 1984). Nevertheless, in this study the interpretation of results was made considering that pseudoreplication is the testing for treatment effects with an error term inappropriate to the hy- pothesis being considered (Hurlbert, 1984). Although we must avoid the use of “pseudoreplicant s”, if t he va riab ility wit hin o ur “small experimental units” is considerable (as in the case of this study) we can assume certain significance of the estimations resulting from “pseudoreplicants”. Despite criticism for pseu- doreplication, nonreplicated studies might nevertheless produce strong ecological evidence (Millar & Anderson, 2004; Hawkins, 1986 [cited by Hargrove & Pickering, 1992]). However, such significance is related not to the treatments, but to the locations where the treatments were applied. Methodology Study Area The study area was located at 5 km to the west of Tapalpa town (Jalisco state), in the west-central region of Mexico (Figure 1). This area is located within the 19˚56' and 19˚58' North latitude; 103˚47' and 103˚51' West longitude (Benavides, 1987). The Tapalpa Saw has the following general characteristics (Martínez et al., 1990): Altitude: 1900 - 2400 m.a.s.l. Mean annual rainfall: 883.1 mm. Mean temperature: 16.6˚C (Minimum mean annual 9.1˚C, Maximum mean annual 24.3˚C). This region corre- sponds to a temperate sub-humid climate (Benavides, 1987), and is dominated by Pinus michoacana, Pinus oocarpa, Quercus spp, and Alnus spp. The study area was mostly on north-facing slopes, at an altitude of 2110 m.a.s.l. In average, the slope varied be- tween 15% and 25%. Experimental Design The set of data, used in this work, resulted from measures taken before and after applying two methods of prescribed fires (March, 1991). These two method s were defined according to the direction of the fire in relation to slope: 1) backing fire, and 2) head fire. The experimental design was based on nine sample plots of 20 × 30 m, where both methods were applied in three sample plots re spectively . The remaining three sa mple plots were used as control. In order to analyze statistically the effect of the treatments on regenerations, the sample plots were divided into 24 smaller “sample units” (5 × 5 m each), which were considered as pseudoreplicants. Seedling Evaluation The following factors were measured for each seedling, within each sample unit: 1) number; 2) height; 3) status [live or dead]; 4) vigor; 5) color; and 6) damage. It was not possible to identify the corresponding species. These measures were made three times: 1) Before burni ng. It had the purpose to evaluate the ori- ginal seedlings in the sample units; 2) After burning. The se- cond measures were made three weeks after burning, where all the individuals per sample unit were evaluated; and 3) After two years. In 1993 every seedling was recorded in all the sam- ple units. Data Analysis The data analysis had two objectives: 1) To find out if there was any significant difference between the two treatments in re- lation to the control; and 2) to determine if there was any signi- ficant difference bet ween the tre at ments. T his was achieve d ba sed on a completely randomized design. Where differences were ana- lyzed considering the number of seedlings resulting from each treatment and the control. For this, each sample unit was con- sidered as a pseudoreplicant. This means a total of 72 sample units (replicants) per each treatment and control. The division of the sample plots allowed us to diminish the variance (Steel & Torrie, 1960). Also, an exploratory statistical analysis was ap- plied, in order to show some characteristic of regeneration. -106 -105-104 -103 -102 -106 -105-102 22 21 20 19 22 21 20 19 -104 -103 Figure 1. Approximated location of the Tapalpa Saw, in Jalisco state, Mexico, were prescribed fires were applied. 1 0 Copyright © 2012 SciRes.  J. G. F. GARNICA Results General Fire Behavior The fire behavior during the burnings was rather unstable in both prescribed methods. There was a considerable variation in the height o f flames, which was due to the di fferences in the spa- ti al distribution of fuel load s over the study , and changes of wind speed and direction. Backing fire (March 25th, 1991). Using this backing fire technique, the speed of spread of fire was approximately 24 m/h, under a windless condition. However, there were some gusts of wind of 3 km/hour (from the east) that increase a lit bit fire rate of spread. The wind had not been blowing in the same di- rection as the down-slope direction in which the fire was se t (north aspect). The average height of the flames was 0.5 m, which means low fire intensity (Fuller, 1991). The maximum height of flames was 2.4 m. Head fire (March 26th, 19 91). The rate of spread of fire was almost ten times faster than in the backing fire (276 m/h). At the beginning there were gusts of wind between 3 and 4.5 km/hour (from the east). In general, the average wind speed was 5.5 km/ hour. However, wind had not been blowing in the same direc- ti on. The average height of flame was 1.5 m, whi ch corresponds to a fire intensity of more than 300 kW/m (Luke & McArthur, 1977). The rising of temperature resulted in an increase of wind speed up t o 7.5 and 9 km/hour, which, at the same time, increased the speed of spread of fire. This caused that flames sweep more quickly over the ground level, resulting in a decrease in soil temperature (Whitaker, 1961), which reduces the risk of im- pairment of seedling. However, fire intensity varies according to the amount, moisture content, and structure of the fuel (Chandler et al., 1983a). Original Seedlings before Burning Seedlings that originally were only in the treated plots were evaluated. Table 1 shows the average number of seedlings, and mean height that were found within the treated plots, before burning. T he general mean number of indivi duals (within 72 m2) was 7.2, which is eq uivalent to 995 seedlings per hectare, within a range from 417 to 4028 individuals. Although, on average, there were more individuals in the head fire plots (HFPs) (1157/ ha, SE = 662) than in the backing fire plots (BFPs) (833/ha, SE = 263), this difference was not significant (F = 0.189; FCrit. = 7.708 [P < 0.05]). Table 1. Number and mean height of seedlings per plot, before burning, for both treatments. Also the corresponding number of seedlings per hectare is shown. Treatment Plot Number of seedlings Mean of number of seedlings/haMean height 1 9.5 1319 31.5 2 3 417 26.7 3 5.5 764 26.2 Backing fire Mean 6 833 28.1 4 1.5 208 23.0 5 6 833 19.0 6 17.5 2430 24.5 Head fire Mean 8.3 1157 22.2 Total mean 7.2 995.2 25.2 The mean height was 25.2 cm, within a range from 15.1 to 43 cm. The mean height of individuals in the BFPs was 28.1 cm (SE = 2.69), while for HFPs it was 22.2 (SE = 1.64). The difference between the BFPs and HFPs was not significant (F = 6.46; FCrit. = 7.708). Number of Seedlings after Burning Immediately after burning 93.6% of the existing seedlings and root sprouts were dead. The mean number of seedlings per plot (600 m2) was 34 and the standard error was 21.1. This is equivalent to 561 seedlings per hectare, a quite lower value than that recorded in the original regeneration. As was expected, the mean height of the seedlings was very similar to that before burning ( 26.8 cm). The ra nge in heig ht was 10 - 160 cm, wit h 20 - 30 cm being the modal height class. Only 10 of the 202 seed- lings recorded in the burnt plots survived to fire. All of them were located in plot 1, within the backing fire plot. Two years later a census of all the plots was carried out (Fe- bruary 12-24, 1993). Both live and dead individuals were re- corded. Table 2 shows a summary of the results of this survey. On average the BFPs had a mean of 86 seedlings per plot. The head fire plots had a very similar mean of 88 seedlings per plot, while the control plots had an average of only 34 seedlings per plot. However, plot 9 (control) showed a high density of seed- lings (88). This can be explained because this plot had suffered certain perturbations, such as trees harvesting, which moved litter off the forest floor, allowing seeds to come into touch with mineral soil. This did not occur in the other plots; therefore this plot (9) was excluded from other comparisons. Comparison of Seedlings before and after Burning Comparison between treatments. Figure 2 shows a graphi- cal comparison be tween the original regeneration (1991) and the re generation produced two y ears after burning (1993). There was a notably higher mean density of regeneration after two years. Both treatments show almost the same production of regenera- tion. The density of seedlings was very much less in the control plots. Table 3 summarizes the corresponding averages of rege- neration before and after burning, per plot. No data were collec- ted from the control plots before burning. However, it is expec- ted that the number of seedlings was very similar than the esti- mated two years after burning the treated plots. Table 2. Number of seedlings per plot and per treatment, two years after burn- ing. Backing fire Head fire Control Plot 1 2 3 4 5 6 7 8 9 Total767111347142 75 13 1 88 Mean 86 88 34 SE 13.2 28.2 27.3 S/ha 1267 1183 1883783 2367 1250 217 171467 Mean 1433 1467 567 SE = Standard error; S/ha = Seedlings per hectare. Copyright © 2012 SciRes. 11 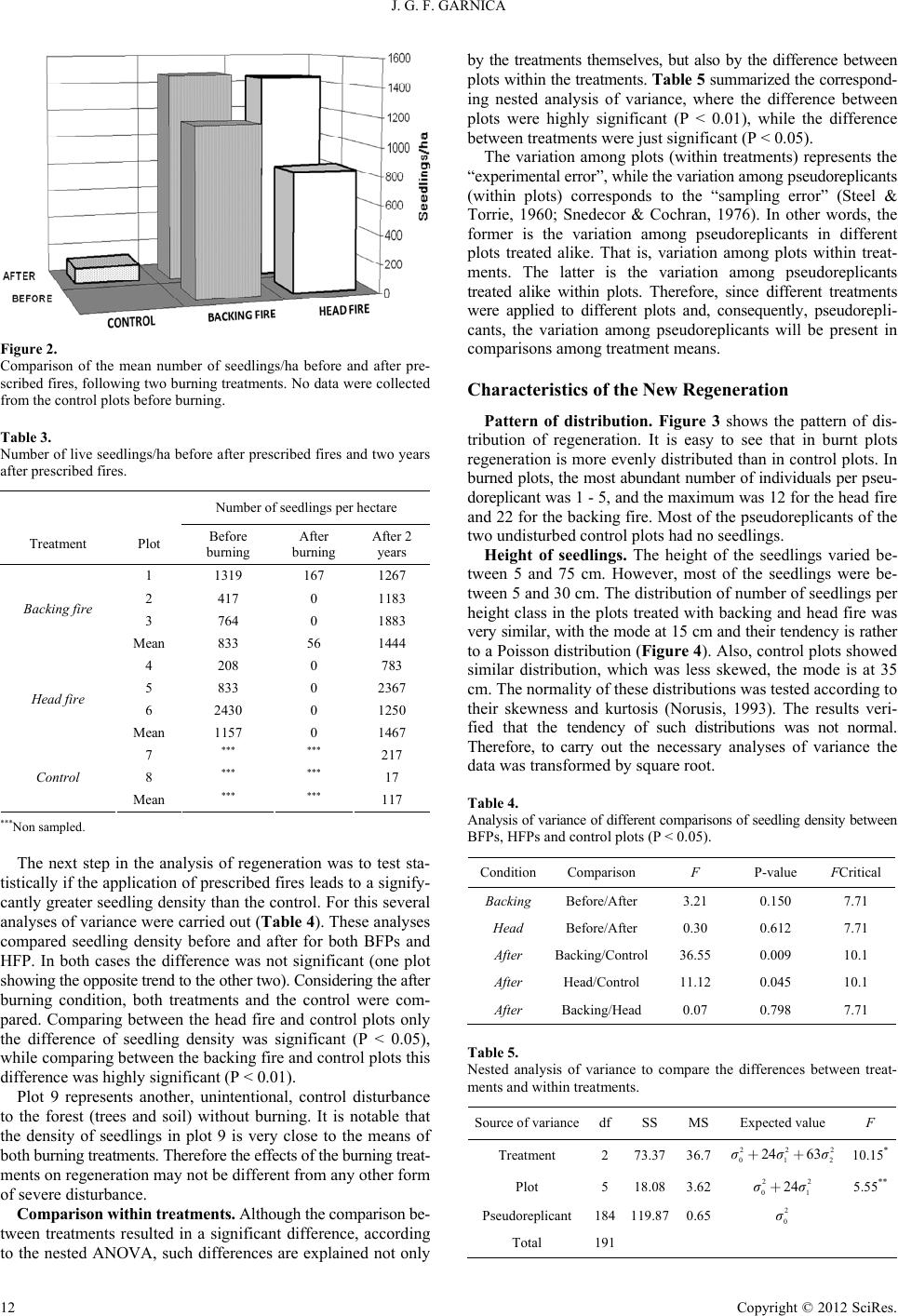 J. G. F. GARNICA Figure 2. Comparison of the mean number of seedlings/ha before and after pre- scribed fires, following two burning treatments. No data were collected from the control plots befor e burning. Table 3. Number of live seedlings/ha before after prescribed fires and two years after prescribed fires. Number of seedlings per hectare Treatment Plot Before burning After burning After 2 years 1 1319 167 1267 2 417 0 1183 3 764 0 1883 Backing fire Mean 833 56 1444 4 208 0 783 5 833 0 2367 6 2430 0 1250 Head fire Mean 1157 0 1467 7 *** *** 217 8 *** *** 17 Control Mean *** *** 117 ***Non sampled. The next step in the analysis of regeneration was to test sta- tistically if the application of prescribed fires leads to a signify- cantly greater seedling density than the control. For this several analyses of variance were carried out (Table 4). These analyses compared seedling density before and after for both BFPs and HFP. In both cases the difference was not significant (one plot showing the opposite trend to the other two). Considering the after burning condition, both treatments and the control were com- pared. Comparing between the head fire and control plots only the difference of seedling density was significant (P < 0.05), while comparing between the backing fire and control plots this difference was highly significant (P < 0.01). Plot 9 represents another, unintentional, control disturbance to the forest (trees and soil) without burning. It is notable that the density of seedlings in plot 9 is very close to the means of both burning treatments. Therefore the effects of the burning treat- ments on regeneration may not be different from any other form of severe disturbance. Comparison within treatments. Although the comparison be- tween treatments resulted in a significant difference, according to the nested ANOVA, such differences are explained not only by the treatments themselves, but also by the difference between plots withi n the treatments. Table 5 summarized the correspond- ing nested analysis of variance, where the difference between plots were highly significant (P < 0.01), while the difference between treatments were just significant (P < 0.05). The variation among plots (within treatments) represents the “experi mental error”, while the v a riation among pseudoreplicants (within plots) corresponds to the “sampling error” (Steel & Torrie, 1960; Snedecor & Cochran, 1976). In other words, the former is the variation among pseudoreplicants in different plots treated alike. That is, variation among plots within treat- ments. The latter is the variation among pseudoreplicants treated alike within plots. Therefore, since different treatments were applied to different plots and, consequently, pseudorepli- cants, the variation among pseudoreplicants will be present in comparisons among treatment means. Characteristics of the New Regeneration Pattern of distribution. Figure 3 shows the pattern of dis- tribution of regeneration. It is easy to see that in burnt plots regeneration is more evenly distributed than in control plots. In burned plots, the most abundant number of individuals per pseu- doreplicant was 1 - 5, and the maximum was 12 for the head fi re and 22 for the backing fire. Most of the pseudoreplicants of the two undisturbed control plots had no seedlings. Height of seedlings. The height of the seedlings varied be- tween 5 and 75 cm. However, most of the seedlings were be- tween 5 and 30 cm. The distribution of number of seedlings per height class in the plots treated with backing and head fire was very similar, with the mode at 15 cm and t heir tendency is rather to a Poisson distribution (Figure 4). Also, control plots showed similar distribution, which was less skewed, the mode is at 35 cm. The normality of the se distributions was teste d according to their skewness and kurtosis (Norusis, 1993). The results veri- fied that the tendency of such distributions was not normal. Therefore, to carry out the necessary analyses of variance the data was transformed by square root. Table 4. Analysis of variance of different comparisons of seedling density between BFPs, HFPs and control plots (P < 0.05). ConditionComparison F P-value FCritical Backing Before/After 3.21 0.150 7.71 Head Before/After 0.30 0.612 7.71 After Backing/Control36.55 0.009 10.1 After Head/Control 11.12 0.045 10.1 After Backing/Head0.07 0.798 7.71 Table 5. Nested analysis of variance to compare the differences between treat- ments and within treatments. Source of variancedfSS MS Expected value F Treatment 2 73.3736.7 22 01 24 63σσ++ 2 2 σ 10.15* Plot 5 18.083.62 22 01 24σσ+ 5.55** Pseudoreplicant 184119.870.65 2 0 σ Total 191 1 2 Copyright © 2012 SciRes. 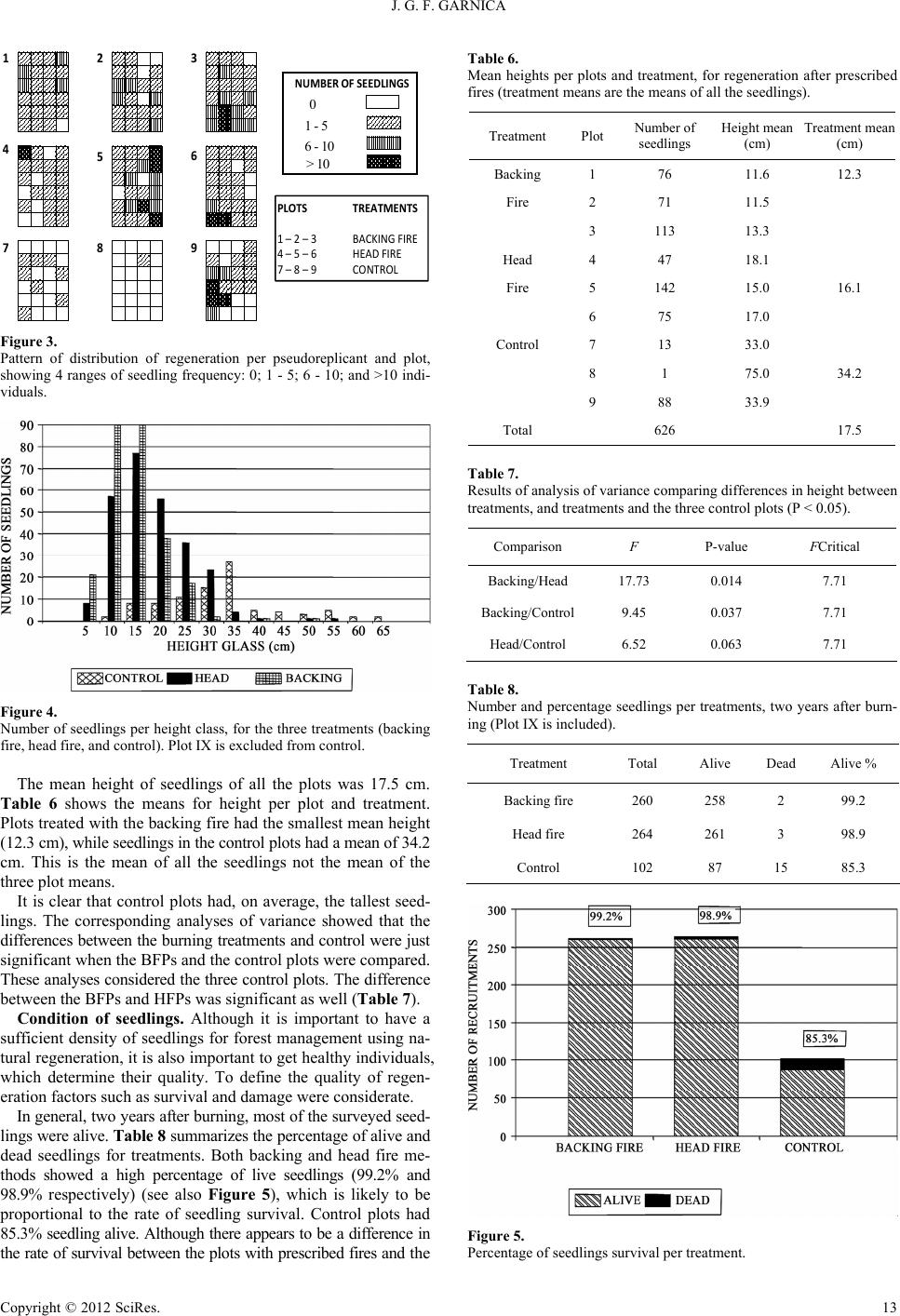 J. G. F. GARNICA UMBER O F SEEDLINGS 0 1 - 5 6 - 10 > 10 TREATMENTS PLOTS 1 - 2 - 3 4 - 5 - 6 7 - 8 - 9 BACK ING FIRE HEAD FIRE CONTROL 3 4 5 6 7 8 9 1 2 PLOTS TREATMENTS 1 –2 –3BACKING FIRE 4 –5 –6HEAD FIRE 7 –8 –9CONTROL NUMBER OF SEEDLINGS 1 4 2 3 7 56 8 9 Figure 3. Pattern of distribution of regeneration per pseudoreplicant and plot, showing 4 ranges of seedling frequency: 0; 1 - 5; 6 - 10; and >10 indi- viduals. Figure 4. Number of seedlings per height class, for the three treatments (backing fire, head fire, and contr ol). Plot IX is excluded from control. The mean height of seedlings of all the plots was 17.5 cm. Table 6 shows the means for height per plot and treatment. Plots treated with the backing fire had the smallest mean height (12.3 cm), while seedlings in the control plot s had a mean of 34.2 cm. This is the mean of all the seedlings not the mean of the three plot means. It is clear that control plots had, on average, the tallest seed- lings. The corresponding analyses of variance showed that the differences between the burning t reatments and control were just significant when the BFPs and the control plots were compared. These analyses considered the three control plots. The difference between the BFPs and HFPs was significant as well (Table 7). Condition of seedlings. Although it is important to have a sufficient density of seedlings for forest management using na- tural regeneration, it is also important to get healthy individuals, which determine their quality. To define the quality of regen- eration factors such as survival and damage were considerate. In general, two years after burning, most of the survey ed seed- lings were alive. Table 8 summarizes the percentage of alive and dead seedlings for treatments. Both backing and head fire me- thods showed a high percentage of live seedlings (99.2% and 98.9% respectively) (see also Figure 5), which is likely to be proportional to the rate of seedling survival. Control plots had 85.3% seedling alive. Although there appe ars to be a difference in Table 6. hts per plots and treatment, for regeneration after prescribed Mean heig fires (treatment means ar e the means of all the seedlings). the rate of sur vival between the plo ts with pres cribed fires and t h e Treatment Plot Number of Height mean Tr eatment mean seedlings (cm) (cm) Backing 1 76 11.6 12.3 Fire 2 71 11.5 3 113 13.3 Hd 4 1 Control 7 3 Tl 1 ea47 18.1 Fire 5 142 15.0 6.1 6 75 17.0 13 33.0 8 1 75.0 4.2 9 88 33.9 ota626 7.5 able 7. analysis of variance comparing differences in height between Comparison F P-value FCritical T Results of treatments, and treatments and the three contr ol pl ot s (P < 0.05). Backing/Head 17.73 0.014 7.71 Blacking/Contro9.45 0.037 7.71 Head/Control 6.52 0.063 7.71 able 8. centage seedlings per treatments, two years after burn- Treatment Total Alive Dead Alive % T Number and per ing (Plot IX is included) . Backing fire 260 258 2 99.2 Head fire 264 261 3 98.9 15 Control 102 87 85.3 Figure 5. of seedlings survival per treatment. Percentage Copyright © 2012 SciRes. 13  J. G. F. GARNICA control plot s, the corresponding analysis of variance did not cor- damage, these on A Due to thants, used in st ods en able 9. damage to seedlings that were found and the sections of the Section of the plant Type of dam age trees of such species recoloniz rapidly in recently roborate this. However, since plot 8 [control] had only a single seedling, this was not a powerful statistical test. Although seedlings showed different types of ly affected a very low proportion of the seedlings (6.4% on average). Table 9 shows a list of the types of damage that were found, as well as the affected part of the plant. These types of damage were n ot exclusive to cert ain treatment s. Although it wa s not the purpose of this study to evaluate the type of damage and its frequency, it would be important to analyze whether there is any relationship between certain types of damage and the prac- tice of prescribed fires. Discussion and Conclusion e variability within the pseudoreplic udy, was considerable it could be assumed certain significance of the resulting estimations. Therefore, despite that this was a nonreplicated study; the results suggest strong ecological evi- dence that prescribed fire enhance natural regeneration of Pinus michoacana and Pinus oocarpa. Comparing between the rege- neration before and after burning, in the treated plots, it is clear that there was an improvement in the density of the natural re- generation. The mean density was nearly doubled after burning. However, this difference was not statistically significant, which could reflect the high variance within the treated and the control plots. Added to this, it is possible to consider that, according to Keeley (1987), although opportunities for population expansion increase after fire for some species, these opportunities increase in the long absence of fire for others. For example, Acer ru- brum seedlings respond negatively to fire, both in terms of survival and reproduction (Reich & Abrams, 1990). In some species, such as Pinus halepensis, a fter a fire the regeneration is retarded during the first 2 - 3 years (Moravec, 1990). Therefore, to estimate with more accuracy the fire effects on regeneration, according to Bradstock and Myersough (1981) and Sirois and Payette (1991), in future studies it will be necessary to record other aspects, such as seed bearer density and damage or de- struction of the seed supply [including the soil seed bank]. Compared with the undi sturbed control, both bur ning meth hanced the mean density of natural regeneration. These results are similar to the obtained in other studies. For example, St Pierre et al. (1992) described that after fire the regeneration of jack pine (Pinus banksiana) was increased. The reason of this is because obligate post-fire seeding species tend to produce a great proportion of viable seeds following fire (Bell et al., 1993). Fire is the necessary trigger for general seed release for many species (Teich, 1970; Enright & Lamont, 1989). Moreover, T Types of plant affected. Whole Direct damage (ca e and grow burnt sites (Savill & Evans, 1986). Van Lear and Waldrop (1991) suggest that southern pines in the USA are not only fire tolerant, but also they require fire for their self-perpetuation. Nevertheless, I did not found any specific information on the fire-dependency of the regeneration of P. michoacana and P. oocarpa. Due to the frequency of fire in the study area, the study of the fire-dependency of such species would be useful to support the decision making process of forest managers. Before treatments, the plots of both firing methods did not dif fer significantly in their number of seedlings. Also, their mean heights were not significantly different. This meant that the two firing methods were applied to plots that were similar in their regeneration. Due to the small mean size of the seedlings and the similarity between P. michoacana and P. oocarpa at such size, it was not possible to separate the two species. Neverthe- less, because some of the enumerated “seedlings” were shoot sprouts, it would be possible to define to which species they be- long. Therefore, in future studies it would be better to different- tiate whether the stems are seedlings or root sprouts. Further- more, their distribution has to be considered. This information will help to evaluate the original conditions of a forest stand, and therefore to understand better the changes caused by fire. Although it was considered that the prescribed fires were of low intensity, they were sufficie ntly intense to kill almost all the original regene ration. Hence, it is important that forest managers determine if the regeneration is already sufficient, in which case it could be bette r not to apply any prescribed fire. If more abund- ant regeneration will come after a prescribed burning, the use of fire may be justified. After burning, plot 9 (control) showed a high density of seed- li ngs, which did not corresp onded to the results in the other con- trol plots. This can be explained because this plot was subjected to certain perturbations, such as trees harvesting, which remo- ved litter from the forest floor, allowing seeds to be in touch with mineral soil. This favorably affected the establishment of natu ral re ge nerat io n. Al tho ugh p resc ribe d fi re may resul t in abu nd- ant seedlings of certain species (Pase & Lindenmuth, 1971), this also depends on other factors, such as rainfall patterns (Bradstock & Myerscough, 1981). In this study only the re- generation after the two firing methods and the control were considered. Although the difference in the number of seedlings between treatments was significant, there wa s also a high variation within them. This may be because the conditions (light, moisture, seed- bank etc.) varied from plot to plot (Steel & Torrie, 1960). Hence, although in this study every effort was made to locate the experimental plots in the same conditions (slope, amount of fuel, number of trees, etc.), in future stud ies it wi ll be ne cess ary to consider more factors in order to ensure homogeneity of experimental plots. If we can defined an enough number of replicates; a blocked design could be used in order to group sample units based on a given criteria, trying that sample units be selected randomly. Although the intensity of fires has been recognized as an im- po used by cattle) Curled Shriveled Needles Roots d Fungus Dried Dried Forked Stem Deforme rtant factor in subsequent regeneration (Whittaker, 1961), no difference was found between the two firing methods in this study. Thus the use of one or other methods may depend on other criteria, such as safety, labor or cost. Some species have the same regeneration response regardless of fire intensity (Ma- lanson & O’Leary, 1982). However, excessively high soil tem- peratures, produced by fire, are important with regard to the 1 4 Copyright © 2012 SciRes.  J. G. F. GARNICA death or survival of seeds or other plant organs (Beadle, 1940). The distribution of seedlings along the plots area was affec- ted by the abundance of root sprouts. In some species, resprout- in e individuals of the or live. In contrast, the control pl ce or in REFERENCES Arno, S. F. (1980). Forethern Rockies. Journal of Forestry, 78, 460-4 g plants contribute most to post-fire recovery (Malanson & O’Leary, 1982). This is more likely if fires that are too frequent for seed production (Smith, 1986). Also, the spatial distribution of seedli ngs is a ff e cted b y t h e loc ation of the old trees, an d seed ling density tends to be higher at a distance from the burned pine can- opy and lower near the burned pine trunk (Ne’eman et al., 1992). Furthermore, regeneration is much denser and better distributed on areas with a good environment for germination, for example where slash is burned (Johnston, 1971; Whipple, 1978; Tweed, 1987). Thus I this study prescribed fire was effective in the preparation of seedbeds for regeneration. The rege neration of some specie s can recover from fire ( Perera, 1989), such as in this study, where som iginal regeneration survived the fire. This explains why some ind ivi du als, recorded after the burning, were up to 75 cm height. The tallest individuals were root sprouts, which were located predominantly in the control plots. In bot h backin g fire an d head fi re plot s there wa s a very hig h percentage of seedlings that were a ots showed a notable proportion of dead individuals. This can be explained because the control plots had a thicker layer of litter on the forest floor. Thus, although some seeds germinate successfully on a litter layer, when it loses moisture the seed- lings start to dry and as a consequence some of them die (Hodgkinson & Oxley, 1990). Therefore, the amount and type of fuel on the forest floor influences the percentage of seeds, which germinated, and/or seedlings that survived. This is more noticeable where fine fuels are abundant, which lost moisture faster (Blackmar, 1971; Simard & Main, 1982). Finally, the results produced in this work could support a re- commendation to use prescribed fires, in order to enhan crement forest regeneration. However, it is important to con- sider that thi s recommendation co uld only apply for regions with similar conditions to the study area. This is remarkably impor- tant due to the fact that in Mexico we can find around 50 spe- cies of pine. st fire history in the Nor 66. Beadle, N. C. W. (1940). Soil temperatures during forest fires and their effect on the survival of vegetation. Journal of Ecology, 28, 180-192. doi:10.2307/2256168 Bell, D. T., Plummer, J. A., & Taylor, S. K. (1993). Seed germination ecology in Southwestern West Australia. The Botanical Review, 59, 24-73. doi:10.1007/BF02856612 Benavides, S. J. D. (1987). Estimación de la calidad de sitio mediante indices de sitio del Pinus michoacana cornuta Martínez y Pinus oocarpa Schiede, para el A.D.F. Tapalpa, Estado de Jalisco. Bachellor Thesis, Chapingo: División de Ciencias Forestales. U.A. Chapingo. lackmar, W. H. (1971). Equilibrium moisture content of common fire fuels found in southeastern forests. Forest Service Research Paper, B No. SE-74. Southwestern Forest Experiment Station. USA. Bradstock, R. A., & Myerscough, P. J. (1981). Fire effects on seed release and the emergence and establishment of seedlings in Banksia ericifolia L.f. Australian Journal of Botany, 29, 521-531. doi:10.1071/BT9810521 ain, M. D. (1985). Prescribed winter burns can reduce the growth of nine-year-old Loblolly pine. Research Note SO-312. Souther Cn Forest C aviour and effects (p. D roceedings of the Federal D sites and seedling recruitment in five co-occurring Banksia species. Experimental Stati o n . USDA, Forest Service. handler, C., Cheney, P., Thomas, P., Trabaud, L., & Williams, D. (1983a). Fire in the forestry. Forest fire beh 450). New York: John Wiley & Sons, Inc. eBano, L. F. (1976). Nutrients lost in debris and runoff water from a burned chaparral watershed. Conference P Inter-Agency Sediment, 3, 13-27. avis, K. P. (1959). Forest fire control and use. New York: McGraw- Hill. Enright, N. J., & Lamont, B. B. (1989). Seed banks, fire season, safe Journal of Ecology, 77, 1111-1112. doi:10.2307/2260826 uller, M. (1991). Forest fires. An introduction to wildland fire behav- iour, management, fire fighting and prevention. New Y Fork: John G volution, 35, 101-123. doi:10.2307/2407945 Wiley & Sons. ivnish, T. J. (1981). Serotiny, geography, and fire in the pine barrens of New Jersey. E growth Haywood, J. (2007). Influence of herbicides and felling, fertilization, and prescribed fire on longleaf pine establishment and through six growing seasons. New Forests, 33, 257-279. doi:10.1007/s11056-006-9026-3 effner, R. A., Butler IV, M. J., & Reilly, C. K. (1996). Pseud revisited. Ecology, 77, 2558-2562 Horeplication . doi:10.2307/2265754 odgkinson, K. C., & Oxley, R. E. (1 factors on germination H990). Influence of fire and edaphic of the arid zone shrubs Acacia aneura, Cassia nemophila and Dodonaea viscosa. Australian Journal of Botany, 38, 269-279. doi:10.1071/BT9900269 oule, G., & Payatte, S. (1991). Seed dynamics of Abies balsamea and Acer saccharum in a deciduous fore Hst of Northeastern North America. American Journal of Botany, 78, 895-905. doi:10.2307/2445168 udson, J., & Salazar, M. (1981). Prescribed Honduras. Miscellane Hfires in the pine forests of ous Serie No. 1. Siguatepeque: National H Monographs, 54, 187 -211. School of Forest Science s . urlbert, S. H. (1984). Pseudoreplication and the design of ecological field experiments. Ecological doi:10.2307/1942661 nkins, M. A., Klein, R. N., & McDaniel, V. L. (2011). Y regeneration as a funct Je ellow pine ion of fire severity and post-burn stand struc- ture in the southern Appalachian Mountains. Forest Ecology and Management, 22, 681-691. doi:10.1016/j.foreco.2011.05.001 hnston, W. F. (1971). Broadcast burning slash favors black spruce reproduction on organic soil in Minnesota. The Forest Chroni Jo cle, 47, Lre pressure and shortleaf pine (Pinus echinata) regeneration 33-35. and, A. D., & Rieske, L. K. (2006). Interactions among prescribed fire, herbivo following southern pine beetle (Dendroctonus frontalis) mortality Original. Forest Ecology and Management, 235, 260-269. doi:10.1016/j.foreco.2006.08.336 uke, R. H., & McArthur, A. G. (1978). Bushfires in Aust partment of Primary Industry For Lralia. De- estry and Timber Bureau, Com- Mrubs. Oecologia, 53, 355-358. monwealth of Australia. alanson, G. P., & O’Leary, J. F. (1982). Post-fire regeneration of californian coastal sage sh doi:10.1007/BF00389013 artínez, M. A., Flores, G. J. G., & de D. Benavides S Índices de riesgo de ince M. J., (1990). ndio en la sierra de Tapalpa, Estado de MJalisco. Revista Ciencia Forestal en México, 15, 3-34. cNabb, K. L. (2001). Prescribed burning in Alabama forests. Alabama: Alabama A & M and Alburn Universities. Millar, R. B., & Anderson, M. J. (2004). Remedies for ps eudoreplication. Fisheries Research, 70, 397-40 7. doi:10.1016/j.fishres.2004.08.016 oravec, J. (1990). Regeneration o forests following fire. Vegetatio, 87 Mf N. W. African Pinus halepensis , 29-36. doi:10.1007/BF00045652 e’eman, G., Lahav, H., & Izhaki, I. (1992). lings 1 year after fire in a N Spatial pattern of seed- Mediterranean pine forest. Oecologia, 91, 365-370. doi:10.1007/BF00317625 Copyright © 2012 SciRes. 15  J. G. F. GARNICA N of effects from prescribed fires esmith, J. C. B., Caprio, A. C., Pfaff, A. H., McGinnis, T. W., & Keeley, J. E. (2011). A comparison and wildfires for resource objectives in Sequoia and Kings Canyon National Parks. Forest Ecology and Management, 261, 1275-1282. doi:10.1016/j.foreco.2011.01.006 orusis, M. P. (1993). SPSS for Windows. Base System. Chicago: SPS Inc. NS l of Forestry, 69, 800-805. n Sri Lanka. Forest Ecology and Pase, C. P., & Lindenmuth Jr., A. W. (1971). Effects of prescribed fire. Journa Perera, A. (1989). Post-fire recovery of 10-year-old Pinus caribaea var. hondurensis in a hilly watershed i Management, 28, 309-313. doi:10.1016/0378-1127(89)90010-8 amade, F. (1984). Ecology of natural resources. New York: John Wiley & Sons. R amics of Central Wisconsin oak forest regeneration. Reich, P. B., & Abrams, M. D. (1990). Fire affects ecophysiology and community dyn Ecology, 71, 2179-2190. doi:10.2307/1938631 avill, P. S., & Evans, J. (1986). Plantation silviculture in temperate regions. Oxford: Claredon Press. S n Journal of Forest Research, 12, Simard, A. J., & Main, W. A. (1982). Comparing methods of predicting jack pine slash moisture. Canadia 793-802. doi:10.1139/x82-119 irois, L., & Payette, S. (1991). Reduced postfire tree regeneration along a boreal forest-forest-tu Sndra transect in Northern Quebec. Ecology, 72, 619-627. doi:10.2307/2937202 mith, D. A. (1986). The practice of silviculture (8th ed.). New York: John Wiley & Sons. S Iowa State University Press. inus banksiana) Snedecor, G. W., & Cochran, W. A. (1976). Statistical methods (6th ed.). Ames, Iowa: The St-Pierre, H., Gagnon, R., & Bellefleur, P. (1992). Post-fire regenera- tion of black spruce (Picea mariana) and Jack p ine (P in the boreal forest, Quebec. Canadian Journal of Forest Research, 22, 474-481. doi:10.1139/x92-062 teel, R. G. D., & Torrie, J. H. (1960). Principles and procedures of statistics. New York: McGra w-Hill SBook Company, Inc. Topulation eich, A. H. (1970). Cone serotiny and inbreeding in natural p of Pinus banksiana and Pinus contorta. Canadian Journal of Botany, 48, 1805-1809. doi:10.1139/b70-265 oledo, M. R. (1988). Levels of forest Tfire risk. Michoacan: Regional T. Born of fire. National Parks, 61, 22. burning for Center for Reaserches in Forestry, Agricultural, and Animal Hus- bandry. weed, W. (1987) Van Lear, D. H., & Waldrop, T. A. (1991). Prescribed regeneration. In M. L. Duryea, & P. M. Dougherty (Eds.), Klumer forest regeneration manual (pp. 235-249). Netherlands: Academic Publishers. doi:10.1007/978-94-011-3800-0_12 ega, J. A., Fernández, C., Pérez, G. P., & FontuVrbel, T. (2008). The influence of fire severity, seronity, and post-fire management on Pinus pinaster ait. Recruitment I three burnt areas in Galicia (NW Spain). Forest Ecology and Management, 256, 1596-1603. doi:10.1016/j.foreco.2008.07.005 ose, J. M., & White, A. S. (1987).V Processes of understory seedling recruitment 1 year after prescribed fire in an Arizona ponderosa pine community. Canadian Journal of Botany, 65, 2280-2290. doi:10.1139/b87-311 ade, D. D. (1989). A Wguide for prescribed fire in southern forests. Wmpbell, R. E., de Bano, L. F., Lewis, C. E., Fred riksen , W (1978). The relationship of buried, germinating seeds to Technical Publication R8-TP 11. Southern Region, Forest Service, USDA. ells, C. G., Ca R. L, Franklin, E. C., Froelich, R. C., & Dunn, P. H. (1979). Effects of fire on soil. General Technical Report WO-7. Forest Service, USDA. hipple, S. A. vegetation in an old-growth Colorado supalpine forest. Canadian Journal of Botany, 56, 1505-1509. doi:10.1139/b78-176 hittaker, E. (1961). Temperatures in heath fires. Journal Wof Ecology, 49, 709-715. doi:10.2307/2257233 illiam, W., Hargrove, W. W., & Pickering, J. (1992). Pseudoreplica-Wtion: A sine qua non for regional ecology. Landscape Ecology, 6, 251-258. doi:10.1007/BF00129703 1 6 Copyright © 2012 SciRes.
|