Applied Mathematics
Vol.07 No.10(2016), Article ID:67882,24 pages
10.4236/am.2016.710105
The Bistability Theorem in a Model of Metastatic Cancer
Jens Christian Larsen
Vanløse Alle 50 2 mf tv, 2720 Vanløse, Copenhagen, Denmark

Copyright © 2016 by author and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 2 May 2016; accepted 27 June 2016; published 30 June 2016
ABSTRACT
The main theorem of the present paper is the bistability theorem for a four dimensional cancer model, in the variables 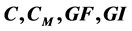 representing primary cancer C, metastatic cancer
representing primary cancer C, metastatic cancer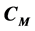 , growth factor GF and growth inhibitor GI, respectively. It says that for some values of the para- meters this system is bistable, in the sense that there are exactly two positive singular points of this vector field. And one is stable and the other unstable. We also find an expression for
, growth factor GF and growth inhibitor GI, respectively. It says that for some values of the para- meters this system is bistable, in the sense that there are exactly two positive singular points of this vector field. And one is stable and the other unstable. We also find an expression for 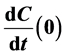 for the discrete model T of the introduction, with variables
for the discrete model T of the introduction, with variables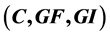 , where C is cancer,
, where C is cancer, 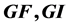 are growth factors and growth inhibitors respectively. We find an affine vector field Y whose time one map is T2 and then compute
are growth factors and growth inhibitors respectively. We find an affine vector field Y whose time one map is T2 and then compute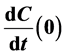 , where
, where 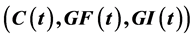 is an integral curve of Y through
is an integral curve of Y through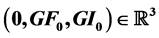 . We also find a formula for the first escape time for the vector field associated to T, see section four.
. We also find a formula for the first escape time for the vector field associated to T, see section four.
Keywords:
Bistability, Cancer, Mass Action Kinetic System, Discrete Dynamical System

1. Introduction
1.1. Summary of the Paper
We continue the study of the cancer model from Larsen (2016) [1] . The model is
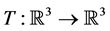
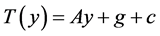
where
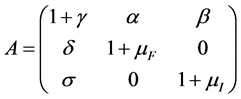
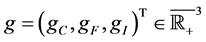 are birth rates and T denotes transpose. Here
are birth rates and T denotes transpose. Here 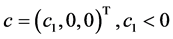 is chemotherapy
is chemotherapy
and 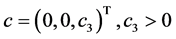 is immune therapy. The parameters
is immune therapy. The parameters ,
,  ,
,  ,
, . We have shown previously Larsen (2016) [1] , that there are affine vector fields on
. We have shown previously Larsen (2016) [1] , that there are affine vector fields on , such that their time one map is T, when the eigenvalues of A have positive real part. This enables you to find a formula for the rate of change of cancer growth in
, such that their time one map is T, when the eigenvalues of A have positive real part. This enables you to find a formula for the rate of change of cancer growth in . The characteristic polynomial of A is
. The characteristic polynomial of A is

when  The discriminant of this polynomial is
The discriminant of this polynomial is

The eigenvalues are

In section two we prove the Bistability Theorem for a mass action kinetic system of metastatic cancer  and primary cancer C. The model also has
and primary cancer C. The model also has  growth factors and
growth factors and  growth inhibitors. We show that for some values of the parameters there are exactly two positive singular points
growth inhibitors. We show that for some values of the parameters there are exactly two positive singular points 
 where
where  We prove that
We prove that  is unstable and
is unstable and  is stable, when one of the rate constants is small.
is stable, when one of the rate constants is small.
For  we have: if the eigenvalue
we have: if the eigenvalue  of A has
of A has  then one can find an affine vector field, whose time one map is
then one can find an affine vector field, whose time one map is . Similarly, when
. Similarly, when  and the eigenvalues
and the eigenvalues  of the cha- racteristic polynomial of A are nonzero, then one can find an affine vector field on
of the cha- racteristic polynomial of A are nonzero, then one can find an affine vector field on , whose time one map is
, whose time one map is . This enables us to find a formula for the rate of change of cancer growth in
. This enables us to find a formula for the rate of change of cancer growth in  This is the subject of Section 3.
This is the subject of Section 3.
The phase space of our model T is . In section four we show, that when
. In section four we show, that when ,
,  ,
,  ,
,  orbits of the vector field associated to T will escape phase space for both
orbits of the vector field associated to T will escape phase space for both  and
and . We obtain a formula for the first escape time. There is a similar treatment for
. We obtain a formula for the first escape time. There is a similar treatment for 
1.2. The Litterature
uPAR (urokinase plasminogen activator receptor) is a cell surface protein, that is associated with invasion and metastasis of cancer cells. In Liu et al. (2014) [2] a cytoplasmic protein Sprouty1 (SPRY1) an inhibitor of the (Ras-mitogen activated protein kinase) MAPK pathway is shown to interact with uPAR and cause it to be degraded. Overexpression of SPRY1 in HCT116 or A549 xenograft in athymic nude mice, led to great suppression of tumor growth. SPRY1 is an inhibitor of the MAPK pathway. Several cancer cells have a low basal expression of SPRY1, e.g. breast, prostate and liver cancer. SPRY1 promotes the lysosomal mediated degradation of uPAR. SPRY1 overexpression results in a decreased expression of uPAR protein. This paper suggests that SPRY1 regulates cell adhesion through an uPAR dependant mechanism. SPRY1 inhibits proliferation via two distinct pathways: 1) SPRY1 is an intrinsic inhibitor of the Raf/MEK/ERK pathway; 2) SPRY1 promotes degradation of uPAR, which leads to inhibition of FAK and ERK activation.
According to Luo and Fu (2014), [3] EGFR (endoplasmic growth factor receptor) tyrosine kinase inhibitors (TKIs) are very efficient against tumors with EGFR activating mutations in the EGFR intracytoplasmic tyrosin kinase domain and cell apoptosis was the result. However some patients developed resistance and this reference aimed to elucidate molecular events involved in the resistance to EGFR-TKIs. The first EGFR-TKI s to be approved by the FDA (Food and Drug Administration, USA) for treatment of NSCLC (non small cell lung cancer) were gefitinib and erlotinib. The mode of action is known. These drugs bind to the ATP binding site of EGFR preventing autophosphorylation and then blocking downstream signalling cascades of pathways RAS/ RAF/MEK/ERK and PI3K/AKT with the results, proliferation inhibition, cell cycle progression delay and cell apoptosis.
There are several important monographs relevant to the present paper, see Adam & Bellomo (1997), [4] , Geha & Notarangelo (2012), [5] , Murphy (2012), [6] , Marks (2009), [7] , Molina (2011), [8] .
2. A mass Action Kinetic Model of Metastatic Cancer
The main result of this section is Theorem 1 below that proves the bistability of the mass action kinetic system (1) to (8). Consider then the mass action kinetic system from Larsen (2016), [9] , in the species  primary cancer cells, metastatic cancer cells, growth factor, growth inhibitor respectively.
primary cancer cells, metastatic cancer cells, growth factor, growth inhibitor respectively.
 (1)
(1)
 (2)
(2)
 (3)
(3)
 (4)
(4)
 (5)
(5)
 (6)
(6)
 (7)
(7)
 (8)
(8)
The complexes are 
 And this defines the rate constants
And this defines the rate constants . With mass action kinetics the ODE s become
. With mass action kinetics the ODE s become




all  We shall now find the singular points of this vector field denoted
We shall now find the singular points of this vector field denoted

But first we state a theorem, we shall next prove. A positive (nonnegative) singular point 
of f is a singular point of f, such that 
 Define
Define

Theorem 1 Assume  When
When 
 there are exactly two positive singular points
there are exactly two positive singular points

where 
 is unstable. Given
is unstable. Given  such that
such that
 and,
and,  then there exists
then there exists  such that
such that 
is stable when 
Consider a singular point  of f and linearize
of f and linearize

Setting the last coordinate of f equal to zero gives

when  Now insert this into the first and second coordinates of f to get
Now insert this into the first and second coordinates of f to get
 (9)
(9)
and
 (10)
(10)
When  we get from (9)
we get from (9)

and from (10) we get

This means that B simplifies to

Let  denote the matrix you obtain by deleting row three and column three in B. Then
denote the matrix you obtain by deleting row three and column three in B. Then

Also

The characteristic polynomial of  is denoted
is denoted

Finally

In Larsen (2016) [9] , we found two cubic polynomials  such that
such that

whenever  is a nonnegative singular point of f. We shall need the following lemma.
is a nonnegative singular point of f. We shall need the following lemma.
Lemma 1 Assume  Then
Then

where



Proof. The coefficient to  is according to Larsen (2016), [9]
is according to Larsen (2016), [9]

 and
and 
 The coefficient to
The coefficient to  is according to Larsen (2016), [9]
is according to Larsen (2016), [9]

Everything cancels out and leaves a zero. The coefficient to  is according to Larsen (2016), [9]
is according to Larsen (2016), [9]

Square  and multiply
and multiply  to get
to get

Everything cancels out except

The coefficient to  is according to Larsen (2016), [9]
is according to Larsen (2016), [9]

Multiply

Everything cancels out except

Finally the constant term is

The lemma follows.
Theorem 2 Assume  When
When 
 there are exactly two positive singular points of f
there are exactly two positive singular points of f

where

Proof. We have

where



and



due to symmetry of  When
When  P and
P and  have two positive roots
have two positive roots

in P and

in , see (15) and (16) below. We are going to verify that
, see (15) and (16) below. We are going to verify that
 (11)
(11)
are singular points of f and that
 (12)
(12)
are not singular points of f. Here

and

Also
 (13)
(13)
 (14)
(14)
We have
 (15)
(15)
and logically equivalent
 (16)
(16)
where  To see (15) compute
To see (15) compute

So

and from this the formula follows. And (16) is a similar computation.
We shall insert (15), (16) in the first coordinate of f, multiplied with 

Now abbreviate  and find
and find

Multiply with  to get
to get

But this amounts to

and this vanishes due to the formula for roots of quadratic polynomials. That the second coordinate vanishes is logically equivalent. So (11) are singular points of f.
We shall now argue, that

is not a singular point of f. To this end define

Insert the formulas (15), (16) for  in the first coordinate of f multiplied with
in the first coordinate of f multiplied with  to get
to get

Multiply with  to find
to find
 (17)
(17)
 (18)
(18)
But (17) is zero by the above and (18) is nonzero. So  is not a singular point. That
is not a singular point. That  is not a singular of f is logically equivalent. The theorem follows.
is not a singular of f is logically equivalent. The theorem follows.
In the remainder of the proof of Theorem 1, we assume, that

We shall now verify that  is unstable. We shall show that
is unstable. We shall show that 
But we have

Simply insert (15) and (16) in the numerator

Now we use that

so

is equivalent to

The right hand side here is negative and the left hand side is positive. Thus  has a positive eigenvalue. So
has a positive eigenvalue. So  is unstable.
is unstable.
We shall now show that  is stable, when
is stable, when  is small. We shall use the Routh Hurwitz criterion. So we start by showing, that
is small. We shall use the Routh Hurwitz criterion. So we start by showing, that  But similarly to the above
But similarly to the above

But this amounts to

which is equivalent to

and this again is equivalent to

and from this it follows that  We have the following formula for
We have the following formula for 

And a formula for 

Define

so that

Now introduce these two formulas in the formulas for 


Notice that  for small
for small  Also
Also

is negative for small  The Routh Hurwitz criterion says in our framework, that
The Routh Hurwitz criterion says in our framework, that 
 is equivalent to stability of
is equivalent to stability of  But
But  is equivalent to
is equivalent to
 because our assumptions imply
because our assumptions imply  So
So  is equivalent to
is equivalent to

This equation holds for small . So
. So  is stable for small
is stable for small . This follows by writing
. This follows by writing

where  and h is smooth. This is the standard trick from singularity theory. Then
and h is smooth. This is the standard trick from singularity theory. Then

And from this it follows that  is stable for small
is stable for small . To be precise, given
. To be precise, given 
 such that
such that  and,
and,  then there exists
then there exists  such that
such that  is stable when
is stable when  Theorem 2 follows.
Theorem 2 follows.
Consider the mass action kinetic system in the species  cancer cells, growth factor, growth inhibitor and a protein, respectively.
cancer cells, growth factor, growth inhibitor and a protein, respectively.
 (19)
(19)
 (20)
(20)
 (21)
(21)
 (22)
(22)
 (23)
(23)
 (24)
(24)
 (25)
(25)
The complexes are 

 And this defines the rate constants
And this defines the rate constants . With mass action kinetics the ODE s become
. With mass action kinetics the ODE s become




see Horn and Jackson (1972), [10] . Notice that (24), (25) are the Brusselator, which is known to have oscillating solutions for some values of the parameters, see Sarmah et al. (2015), [11] . Subtracting  on both sides of (25) gives the reaction
on both sides of (25) gives the reaction  Let
Let 

 With these parameter values and initial conditions
With these parameter values and initial conditions 
 the system oscillates, see Figure 1.
the system oscillates, see Figure 1.
3. Eigenvalues with Negative Real Part
In this section  in the discrete model T of the introduction. The purpose of this section is to find a formula for the rate of change of cancer growth
in the discrete model T of the introduction. The purpose of this section is to find a formula for the rate of change of cancer growth

Figure 1. The oscillating mass action kinetic system. I have plotted P versus C.

on the hyperplane  Here
Here  is an integral curve of the vector field Y, defined below. There are four cases to consider. First assume, that
is an integral curve of the vector field Y, defined below. There are four cases to consider. First assume, that  Let
Let  We shall assume that
We shall assume that  Define
Define

and compute, when 

If  has negative real part we might be able to find an affine vector field whose time one map is
has negative real part we might be able to find an affine vector field whose time one map is . Notice that
. Notice that

By Larsen (2016), [1] ,

Then

Define the vector field
 (26)
(26)
 and let
and let

where  The flow of X is
The flow of X is
 (27)
(27)
 (28)
(28)
where  Also
Also

If


then

Assume that  Then we can let
Then we can let

But this means that

because we have

 So we get
So we get

i.e.  Consider first the immune therapy model
Consider first the immune therapy model

So assuming 

We want to have

and

such that

Here  denotes the time one map of X and
denotes the time one map of X and  Define
Define

Then

Thus

Now

Define

Let  denote the first row in U. Compute letting
denote the first row in U. Compute letting


where  is an integral curve of Y through
is an integral curve of Y through  And, because
And, because 
 this is equal to
this is equal to

Now suppose  and
and  distinct and define
distinct and define

Then

when  because the columns of D are eigenvectors of A corresponding to eigenvalues
because the columns of D are eigenvectors of A corresponding to eigenvalues  respectively. Compute, when
respectively. Compute, when  the inverse
the inverse

Then

Define the vector field
 (29)
(29)
 X has flow
X has flow
 (30)
(30)
and the time one map is

and we want this to be

Then define the vector field

This vector field has time one map

Then arguing as before

and

We can now find

Next consider the chemo therapy model

and initially, that  Define the vector field X by (26). It has flow (27), (28). Define the vector field
Define the vector field X by (26). It has flow (27), (28). Define the vector field

We want this vector field to have time one map

Then we find

Now compute arguing as above

Finally we can find

and this becomes

Now consider the chemo therapy model, when  and
and  distinct. Define the vector field X by (29). It has flow (30). Here
distinct. Define the vector field X by (29). It has flow (30). Here

The second coordinate here should be equal to

while the third coordinate should be equal to

in order that the time one map of  is
is . Now we can find
. Now we can find

and this is simplified to

Remark 1 When  then
then  that is
that is  So
So
by the above you can find an affine vector field whose time one map is . Similarly when
. Similarly when
 then
then  and
and  So by the above, you have a formula for
So by the above, you have a formula for  on
on 
4. Escaping Phase Space
In this section  The phase space of our model T of the introduction is
The phase space of our model T of the introduction is . When
. When  integral curves of B from theorem 1 in Larsen (2016), [1] , starting in
integral curves of B from theorem 1 in Larsen (2016), [1] , starting in  will always escape phase space for both
will always escape phase space for both  and
and  Here
Here

and  where
where

U as in section 3. This vector field, B, has time one map T, see Larsen (2016), [1] , or argue as in Section 3.
The purpose of this section is to prove, that there exists a first escape time , i.e. the existence of a smallest
, i.e. the existence of a smallest , such that
, such that

When  we prove, that either
we prove, that either

or there exists a smallest  such that
such that

Proposition 3 Suppose  Given
Given  then there exists
then there exists 
 such that
such that


Proof. We have the following formula for the flow of B

Here


and

Define


Since  we can define
we can define  by
by

It follows that we have the following formula

Since  the proposition follows.
the proposition follows.
Remark 2 By the proof we have

implies  Here
Here . Let
. Let  denote the smallest positive solution to
denote the smallest positive solution to 
When  we have the following proposition using the definitions
we have the following proposition using the definitions


These formulas are explained in the proof of Proposition 4.
Let  where
where

D as in section 3. B has time one map T, see Larsen (2016), [1] , or argue as in section three.
Proposition 4 Suppose  Let
Let  be given. (i) If
be given. (i) If  then there exists a unique
then there exists a unique  such that
such that

If  then
then

for all .
.
(ii) If  then there exists a unique
then there exists a unique  such that
such that

If  then
then

for all .
.
Proof. First of all the flow of F is


We have the following formula

where  is the first row of D. From this equation, (i) follows. For (ii) write
is the first row of D. From this equation, (i) follows. For (ii) write

From this formula, (ii) follows.
Remark 3 In case (i) of the proposition, if  we have
we have

implies

In case (ii) of the proposition, if  we have
we have

implies

We shall now derive a formula for the first escape time  To start with, assume that
To start with, assume that 
 Notice that
Notice that

and

where



i.e.

Compute


where

If  let
let  If
If  define
define  by
by

Then we have the following formulas
 (31)
(31)
 (32)
(32)
Assume that  Then there exists
Then there exists  such that
such that

for  If there exists
If there exists  such that
such that

we claim that there are atmost finitely many such solutions and hence that there exists a smallest  such that
such that

Assume for contradiction, that there are infinitely many solutions to

By (31) there are exactly  solutions to
solutions to

Since there are infinitely many solutions to  there exist
there exist

in  such that
such that

By the mean value theorem, there exists  such that
such that

 Hence
Hence

 A contradiction and there are only finitely many solutions to
A contradiction and there are only finitely many solutions to  If there exists a
If there exists a  such that
such that  let
let  denote the smallest such number, and otherwise let
denote the smallest such number, and otherwise let 
If  then
then

Since  then
then  Define
Define  by
by
 (33)
(33)
so

By  denote the smallest positive solution to
denote the smallest positive solution to  Suppose
Suppose  and
and  if
if  let
let  otherwise write (33). If
otherwise write (33). If

let  otherwise let
otherwise let

 so that
so that

By  denote the smallest positive
denote the smallest positive . Here
. Here

Suppose  If
If  let
let  otherwise write (33). Then there exists
otherwise write (33). Then there exists  such that
such that  By
By  denote the smallest positive solution to
denote the smallest positive solution to  arguing as above.
arguing as above.
If  for all
for all  let
let  otherwise denote by
otherwise denote by  the smallest positive solution to
the smallest positive solution to  Now define the first escape time
Now define the first escape time  by
by

We shall now find the first escape time when  Then we have
Then we have

and

where



i.e.

Assume in the notation of Proposition 4, that  and let
and let

If  let
let  Now compute
Now compute

and

There are atmost two solutions to  If there exists
If there exists  such that
such that  let
let  denote the smallest such solution, otherwise let
denote the smallest such solution, otherwise let  If there exists
If there exists  such that
such that 
 let
let  denote the smallest such solution, otherwise let
denote the smallest such solution, otherwise let  Now define the first escape time, when
Now define the first escape time, when 

5. Summary and Discussion
In this paper we proved that the model of primary and metastatic cancer in Section 2 is bistable, in the sense, that there are exactly two positive singular points. One of them is unstable, and when one of the rate constants is small the other is stable. Then we found formulas for the rate of change of cancer growth for the model T of the introduction, when for  the eigenvalues
the eigenvalues  are nonzero and for
are nonzero and for  when
when  In section four we proved that there is a first escape time for the flow of the affine vector field associated to T when
In section four we proved that there is a first escape time for the flow of the affine vector field associated to T when  A similar result when
A similar result when  was also treated.
was also treated.
It would be interesting to figure out what happens if the polynomials  of section 2 are cubic polynomials and not quadratic as in Theorem 1.
of section 2 are cubic polynomials and not quadratic as in Theorem 1.
About the References
How do cancer cells coordinate glycolysis and biosynthesis. They do that with the aid of an enzyme called Phosphoglycerate Mutase 1. In the reference [12] , the authors suggest a dynamical system for their findings in a figure at the end of the paper. In the reference [13] , A. K. Laird showed that solid tumors do not grow exponentially, but rather like a Gompertz function. The publications of the author are concerned with semi Riemannian dynamical systems, e.g. Lorentzian Geodesic Flows, see [14] and electrical network theory of countable graphs, see [15] , [16] .
Cite this paper
Jens Christian Larsen, (2016) The Bistability Theorem in a Model of Metastatic Cancer. Applied Mathematics,07,1183-1206. doi: 10.4236/am.2016.710105
References
- 1. Larsen, J.C. (2016) Models of Cancer Growth. Journal of Applied Mathematics and Computing.
- 2. Liu, X., Lan, Y., Zhang, D., Wang, K. and Hua, Z.-C. (2014) SPRY1 Promotes the Degradation of uPAR and Inhibits uPAR-Mediated Cell Adhesion and Proliferation. American Journal of Cancer Research, 4, 683-697.
- 3. Luo, M. and Fu, L.-W. (2014) Redundant Kinase Activation and Resistance of EGFR-Tyrosine Kinase Inhibitors. American Journal of Cancer Research, 4, 608-628.
- 4. John, A. and Bellomo, N. (1997) A Survey of Models for Tumor-Induced Immune System Dynamics. Birkhauser, Boston.
- 5. Geha, R. and Notarangelo, L. (2012) Case Studies in Immunology. Garland Science, Hamden.
- 6. Murphy, K. (2012) Immunobiology. 8th Edition, Garland Science, Hamden.
- 7. Marks, F., Klingmüller, U. and Müller-Decker, K. (2009) Cellular Signal Processing. Garland Science, Hamden.
- 8. Molina-Paris, C. and Lythe, G. (2011) Mathematical Models and Immune Cell Biology. Springer Verlag, New York.
http://dx.doi.org/10.1007/978-1-4419-7725-0 - 9. Larsen, J.C. The Bistability Theorem in a Cancer Model.
- 10. Horn, F. and Jackson, R. (1972) General Mass Action Kinetics. Archive for Rational Mechanics and Analysis, 47, 81- 116.
http://dx.doi.org/10.1007/BF00251225 - 11. Sarmah, H.K., Das, M.C. and Baishya, T.K. (2015) HOPF Bifurcation in a Chemical Model. International Journal for Innovative Research in Science and Technology, 1, 23-33.
- 12. Hitosugi, T., et al. (2012) Phosphoglycerate Mutase 1 Coordinates Glycolysis and Biosynthesis to Promote Tumor Growth. Cancer Cell, 22, 585-600.
- 13. Laird, A.K. (1964) Dynamics of Tumor Growth. British Journal of Cancer, 18, 490-502.
http://dx.doi.org/10.1038/bjc.1964.55 - 14. Larsen, J.C. (1996) Lorentzian Geodesic Flows. Journal of Differential Geometry, 43, 119-170.
- 15. Larsen, J.C. (1991) Dynamical Systems and Semi Riemannian Geometry. Ph.D. Thesis, the Technical University of Denmark, Lyngby.
- 16. Larsen, J.C. (1997) Electrical Network Theory of Countable Graphs. IEEE Transactions, Curcuits and Systems: Fundamental Theory and Applications, 44, 1045-1055.