Applied Mathematics
Vol.05 No.17(2014), Article ID:50913,7 pages
10.4236/am.2014.517268
A Mathematical Model for Schistosomiasis Japonicum with Harmless Delay
Huahua Cao, Shujing Gao*, Xiangyu Zhang, Youquan Luo
College of Mathematics and Computer Science, Gannan Normal University, Ganzhou, China
Email: 870577346@qq.com, *gaosjmath@126.com, xyzhang5@163.com
Copyright © 2014 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 12 August 2014; revised 28 August 2014; accepted 16 September 2014
ABSTRACT
From the lifecycle of schistosome, the phenomenon of time delay is widespread. In this paper, a two-dimensional system is studied that incorporates two time delays which are the incubation period of human and snail, respectively. Our purpose is to demonstrate that the time delays are harmless for stability of equilibria of the system. Further, sufficient conditions of stability of equilibria are obtained.
Keywords:
Stability, Schistosomiasis Japonicum, Time Delay

1. Introduction
Mathematical models ([1] -[7] , etc.) have been used to study the transmission and control of schistosomiasis since the first model that has been given by MacDonald in [8] . MacDonald’s model consists of two differential equations in two state variables that correspond to average parasite burden in the definitive hosts and the prevalence of infection in snails. DAS et al. [5] added a layer of biological realism to these early models to study the delay effect on schistosomiasis transmission with control measures. The model is given by
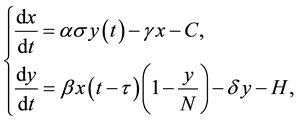 (1)
(1)
where 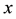 is the current number of egg laying schistosomes in the human host population and
is the current number of egg laying schistosomes in the human host population and 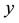 is the current number of infected snails in the environment. Here,
is the current number of infected snails in the environment. Here, 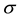 is the human population density per unit accessible water area;
is the human population density per unit accessible water area; 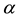 is the multiplication rate due to the infected snail population;
is the multiplication rate due to the infected snail population; 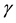 and
and 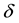 are the intrinsic death rates of two populations
are the intrinsic death rates of two populations 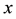 and
and 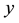 respectively;
respectively; 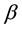 is the simple contact rate;
is the simple contact rate; 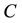 is the constant decay rate due to chemotherapy; and
is the constant decay rate due to chemotherapy; and 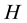 is the constant decay rate by predation or harvesting. Further
is the constant decay rate by predation or harvesting. Further 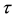 is the incubation period for becoming
is the incubation period for becoming 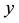 to be infectious. For simplicity, it is assumed that
to be infectious. For simplicity, it is assumed that 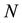 is the constant total population of snails.
is the constant total population of snails.
In [5] , for the sake of mathematical simplicity, they assumed the development of schistosoma is instantaneous. In fact, the developmental time of schistosome is not short. Under normal circumstances, the transit time from parasite eggs to miracidia to infect snail is about 21 days, cercariae are produced about 44 - 159 days after the miracidium penetration in snail hosts. In this paper, we also assume that 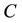 and
and  are the proportions of chemotherapy and predation or harvesting, respectively. Based on the above description, a schistosomiasis model with two time delays is proposed:
are the proportions of chemotherapy and predation or harvesting, respectively. Based on the above description, a schistosomiasis model with two time delays is proposed:
 (2)
(2)
where  is the incubation period for becoming infected human host population and
is the incubation period for becoming infected human host population and  is the transit time from parasite eggs to miracidia to infect snail. We assume that all parameters are positive.
is the transit time from parasite eggs to miracidia to infect snail. We assume that all parameters are positive.
From biological view, we assume that system (2) holds for the time  with given nonnegative initial conditions:
with given nonnegative initial conditions:
 (3)
(3)
where , the Banach space of continuous functions mapping the interval
, the Banach space of continuous functions mapping the interval 
into , where
, where .
.
In the following, we focus on dynamics of system (2) in a nonnegative cone

It is well known by the fundamental theory of functional differential equations [9] that system (2) has a unique solution  satisfying initial conditions (3). It is easy to show that all solutions of system (2) corresponding to initial conditions (3) are defined on
satisfying initial conditions (3). It is easy to show that all solutions of system (2) corresponding to initial conditions (3) are defined on  and remain positive for all
and remain positive for all .
.
The remainder of the paper is organized as follows. In the next section, the stability of the disease-free equilibrium of system (2) is obtained. In Section 3, we investigate the stability of the endemic equilibrium. Some dynamical behaviors are given by numerical simulations in Section 4. This paper is ended with a brief discussion.
2. Stability Analysis of the Disease-Free Equilibrium
In this section, the stability of the disease-free equilibrium of system (2) is investigated.
Using standard methods, it is easy to see that the disease-free equilibrium  always exists.
always exists.
Define the basic reproductive number by

Then for system (2), it is easy to obtain the following result:
(i) If , system (2) has a unique disease-free equilibrium
, system (2) has a unique disease-free equilibrium ;
;
(ii) If , system (1.2) has two equilibria, the disease-free equilibrium
, system (1.2) has two equilibria, the disease-free equilibrium  and the unique endemic equilibrium
and the unique endemic equilibrium , where
, where

In the following, we study the global stability of the disease-free equilibrium  of system (2).
of system (2).
Theorem 2.1. If , the disease-free equilibrium
, the disease-free equilibrium  of system (2) is locally asymptotically stable.
of system (2) is locally asymptotically stable.
Proof. First, according to [9] , the Jacobian matrix at  of system (2) can be written as
of system (2) can be written as
 (4)
(4)
Then the characteristic equation of system (2) at 
 (5)
(5)
where ,
,  ,
,  ,
, .
.
When , (5) becomes into
, (5) becomes into
 (6)
(6)
If , the roots of the equation (6) have negative real parts. Note that
, the roots of the equation (6) have negative real parts. Note that  is equivalent to
is equivalent to . Therefore, if
. Therefore, if  and
and ,
,  is locally asymptotically stable.
is locally asymptotically stable.
Assume that there exists a  such that (5) has pure imaginary roots
such that (5) has pure imaginary roots . Then we have from (5) that
. Then we have from (5) that

Separating real and image parts:

Adding up the squares of both equations, we obtain that
 (7)
(7)
Note that

and  Thus,
Thus,  , which implies that (7) has no positive roots, i.e.,
, which implies that (7) has no positive roots, i.e.,  does not exist. This yields that all roots of (5) have negative real parts if
does not exist. This yields that all roots of (5) have negative real parts if .
.
Next, the global stability of the disease-free equilibrium of system (2) is analyzed. And the strategy of proof is to use Lyapunov functionals and the LaSalle invariance principle.
Theorem 2.2. If , the disease-free equilibrium
, the disease-free equilibrium  is globally asymptotically stable in
is globally asymptotically stable in  for all
for all .
.
Proof. Let  be any positive solution of system (2) with initial conditions (3).
be any positive solution of system (2) with initial conditions (3).
Define
 ,
,
where .
.
Calculating the derivative of  along positive solutions of system (2), it follows that
along positive solutions of system (2), it follows that
 (8)
(8)
Define
 (9)
(9)
We derive from (8) and (9) that
 (10)
(10)
Define
 . (11)
. (11)
It follows from (10) and (11) that
 (12)
(12)
On substituting  and
and  into (12), we obtain that
into (12), we obtain that
 (13)
(13)
If , that is
, that is , it then follows from (13) that
, it then follows from (13) that . By Theorem
. By Theorem
 , the largest invariant subset of
, the largest invariant subset of . Clearly, we see from (13) that
. Clearly, we see from (13) that  if and only if
if and only if . Noting that
. Noting that  is invariant, for each element in
is invariant, for each element in , we have
, we have . It therefore follows from the second equation of system (2) that
. It therefore follows from the second equation of system (2) that

which yields . Hence,
. Hence,  if and only if
if and only if . Accordingly, the global asymptotic stability of
. Accordingly, the global asymptotic stability of  follows from LaSalle’s invariance principle.
follows from LaSalle’s invariance principle.
3. Stability Analysis of the Endemic Equilibrium
It is obtained that the endemic equilibrium  of system (2) is local stable in this section. Further, the global stability of
of system (2) is local stable in this section. Further, the global stability of  is shown if
is shown if .
.
Similar to the proof of Theorem 2.1, the following result is obtained.
Theorem 3.1. If , the endemic equilibrium
, the endemic equilibrium  of system (2) is locally asymptotically stable.
of system (2) is locally asymptotically stable.
Proof. First, according to [9] , the Jacobian matrix at  can be written as
can be written as
 (14)
(14)
Then the characteristic equation of system (2) at :
:
 , (15)
, (15)
where .
.
When , (15) becomes into
, (15) becomes into
 , (16)
, (16)
If , then
, then . It is shown that all the roots of the Equation (16) have negative real parts, suggesting
. It is shown that all the roots of the Equation (16) have negative real parts, suggesting  is locally asymptotically stable.
is locally asymptotically stable.
Assume that there exists a  such that (15) has pure imaginary roots
such that (15) has pure imaginary roots  Then we have from (15) that
Then we have from (15) that

Separating real and image parts:

Adding up the squares of both equations, we obtain that
 (17)
(17)
We know that  if
if , so (17) has no positive roots, i.e.,
, so (17) has no positive roots, i.e.,  does not exist. This yields that all roots of (17) have negative real parts if
does not exist. This yields that all roots of (17) have negative real parts if .
.
Now, we are interested in the global stability of . Then its global stability is investigated by means of Bendixson theorem.
. Then its global stability is investigated by means of Bendixson theorem.
Theorem 3.2. If , the endemic equilibrium
, the endemic equilibrium  is globally asymptotically stable in
is globally asymptotically stable in  when
when .
.
Proof. It is easy to check that equilibrium  of system (2) is unstable if
of system (2) is unstable if . By the above discussion, we know that equilibrium
. By the above discussion, we know that equilibrium  is locally stable if
is locally stable if  and all solutions of system (2) are ultimately bounded in
and all solutions of system (2) are ultimately bounded in . To prove the second assertion, we only prove that system (2) has not periodic orbits in the interior of
. To prove the second assertion, we only prove that system (2) has not periodic orbits in the interior of  if
if .
.
When ,
,

It follows that

which leads to the nonexistence of periodic orbits by Bendixson theorem, therefore,  is globally asymptotically stable.
is globally asymptotically stable.
4. Numerical Simulations
It is reported that cercariae are produced about 44 - 159 days after the miracidium penetration in snails. And the time from parasite eggs to miracidia to infect snail is about 21 days. Therefore, we choose  and
and  in this paper. Further, in this section, we perform some numerical simulations and sensitivity analysis using the following value of parameters:
in this paper. Further, in this section, we perform some numerical simulations and sensitivity analysis using the following value of parameters: ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  ,
, .
.
Thus, we can obtain , the disease-free equilibrium
, the disease-free equilibrium  is asymptotically stable (Figure 1(a)). When
is asymptotically stable (Figure 1(a)). When , the value of other parameters is fixed, we can obtain
, the value of other parameters is fixed, we can obtain  and the unique endemic equilibrium is asymptotically stable (Figure 1(b)). In addition, fixing
and the unique endemic equilibrium is asymptotically stable (Figure 1(b)). In addition, fixing  in simulations, we find that the number of parasite eggs and infectious snails increases as
in simulations, we find that the number of parasite eggs and infectious snails increases as  decreases, respectively (Figure 2).
decreases, respectively (Figure 2).
From the above theorems, we know that the two time delays are harmless. According to the expression of ,
,


Figure 1. The figure (a) shows that a numerical solution of system (2) tends to the disease-free equilibrium as time tends to infinity, where  The figure (b) illustrates that a numerical solution of system (2) tends to the endemic equilibrium as time tends to infinity, where
The figure (b) illustrates that a numerical solution of system (2) tends to the endemic equilibrium as time tends to infinity, where .
.


Figure 2. Simulation results: , 0.042, 0.044, 0.046, 0.048, 0.050 from top to base, respectively. We can find that the smaller of values of
, 0.042, 0.044, 0.046, 0.048, 0.050 from top to base, respectively. We can find that the smaller of values of , the higher of values of parasite eggs and infected snails.
, the higher of values of parasite eggs and infected snails.
the impact of C and H on schistosomiasis transmission is discussed. Fixing , we can see that when
, we can see that when , the endemic equilibrium exists and is stable, when
, the endemic equilibrium exists and is stable, when , the endemic equilibrium doesn’t exist. But the disease-free equilibrium is stable (Figure 3(a)). Analogously, fixing
, the endemic equilibrium doesn’t exist. But the disease-free equilibrium is stable (Figure 3(a)). Analogously, fixing , from Figure 3(b), it is obvious that the disease-free equilibrium is stable when
, from Figure 3(b), it is obvious that the disease-free equilibrium is stable when .
.
From the formula of the basic reproductive number, we know that the basic reproductive number is a decrease function of the rates of chemotherapy and predation or harvesting. This means chemotherapy and predation or harvesting can influence the system.
However, to find out the most influential control measure, we need sensitivity analysis. Now we carry out the sensitivity analysis by calculating the derivation of  on
on  and
and . The derivation is respectively
. The derivation is respectively

From Figure 4(a), we can see that when ,
,  decreases rapidly with the increase of
decreases rapidly with the increase of , the decline of
, the decline of  is not obvious. Similarly,
is not obvious. Similarly,  decreases rapidly with the increase of
decreases rapidly with the increase of  when
when  (Figure 4(b)).
(Figure 4(b)).


Figure 3. Forward bifurcation diagrams for the parasite eggs population.


Figure 4. Sensitivity analysis of  on
on  and
and , respectively.
, respectively.
In brief, the basic reproductive number  is more sensitive when
is more sensitive when  and
and  are small.
are small.
By sensitivity analysis of the basic reproductive number on the rates of chemotherapy and predation or harvesting, we know that the basic reproductive number is a decrease function of the rates of chemotherapy and predation or harvesting. In numerical simulations, we also find that the smaller of values of the rate of chemotherapy, the more sensitive of the basic reproductive number .
.
Although the two time delays are harmless, all of these results imply that the rates of chemotherapy and predation or harvesting can influence the dynamic behaviors. Furthermore, to reduce the prevalence of schistosomiasis infection, to some extent, increasing the rate of predation or harvesting by some measures could achieve better results than increasing the rate of chemotherapy.
5. Conclusions
In this paper, we propose a system of delayed differential equations for schistosomiasis japonicum transmission and obtain sufficient conditions for the existence and local stability of equilibria. Further, global asymptotic stability of the disease-free equilibrium is also studied by constructing suitable Lyapunov functions. When , the disease-free equilibrium is globally asymptotically stable (Figure 5(a)); when
, the disease-free equilibrium is globally asymptotically stable (Figure 5(a)); when , the endemic-free equilibrium is locally asymptotically stable and globally asymptotically stable if
, the endemic-free equilibrium is locally asymptotically stable and globally asymptotically stable if . Thus,
. Thus,  plays an important part in controlling schistosomiasis.
plays an important part in controlling schistosomiasis.
Finally, we guess that the endemic equilibrium should be global asymptotic stable when . And this guess is verified by numerical simulations (Figure 5(b)). This issue will be addressed in future studies.
. And this guess is verified by numerical simulations (Figure 5(b)). This issue will be addressed in future studies.


Figure 5. Phase diagrams: (a) ; (b)
; (b) .
.
Acknowledgements
The research has been partially supported by The Natural Science Foundation of China (No. 11261004), China Postdoctoral Science Foundation funded project (No.
Cite this paper
HuahuaCao,ShujingGao,XiangyuZhang,YouquanLuo, (2014) A Mathematical Model for Schistosomiasis Japonicum with Harmless Delay. Applied Mathematics,05,2807-2814. doi: 10.4236/am.2014.517268
References
- 1. Anderson, R.M. and May, R.M. (1985) Helminth Infections of Humans Mathematical Models, Population Dynamics, and Control. Advances in Parasitology, 24, 1-101.
http://dx.doi.org/10.1016/S0065-308X(08)60561-8 - 2. Barbour, A.D. (1996) Modeling the Transmission of Schistosomiasis: An Introductory View. The American Journal of Tropical Medicine and Hygiene, 55, 135-143.
- 3. Woolhouse, M.E. (1991) On the Application of Mathematical Models of Schistosome Transmission Dynamics. I. Natural Transmission. Acta Tropica, 49, 241-270.
http://dx.doi.org/10.1016/0001-706X(91)90077-W - 4. Yang, H.M. (2003) Comparison between Schistosomiasis Transmission Modeling Considering Acquired Immunity and Age-Structured Contact Pattern with Infested Water. Mathematical Biosciences, 184, 1-26.
http://dx.doi.org/10.1016/S0025-5564(03)00045-2 - 5. Das, P., Mukherjee, D. and Sarkar, A.K. (2006) A Study of Schistosome Transmission Dynamics and Its Control. Journal of Biological Systems, 14, 295-302.
http://dx.doi.org/10.1142/S0218339006001799 - 6. Chiyaka, E.T. and Garira, W. (2009) Mathematical Analysis of the Transmission Dynamics of Schistosomiasis in the Human-Snail Host. Journal of Biological Systems, 17, 397-423.
http://dx.doi.org/10.1142/S0218339009002910 - 7. Qi, L.X. and Cui, J. (2012) Modeling the Schistosomiasis on the Islets in Nanjing. International Journal of Biomathematics, 5, 189-205.
- 8. Macdonald, G. (1965) The Dynamics of Helminth Infections, with Special Reference to Schistosomes. Transactions of the Royal Society of Tropical Medicine and Hygiene, 59, 489-506.
http://dx.doi.org/10.1016/0035-9203(65)90152-5 - 9. Hale, J.K. (1976) Theory of Functional Differential Equations. Springer, New York.
NOTES

*Corresponding author.