Food and Nutrition Sciences
Vol.4 No.8A(2013), Article ID:35289,8 pages DOI:10.4236/fns.2013.48A017
Antioxidant Effect of Roasted Barley (Hordeum vulgare L.) Grain Extract towards Oxidative Stress in Vitro and in Vivo*
![]()
1College of Food Science and Technology, Nanjing Agricultural University, Nanjing, China; 2Department of Food Science and Technology, Egerton University, Egerton, Kenya; 3Department of Nutrition and Food Hygiene, School of Public Health, Southeast University, Nanjing, China.
Email: #marynyambeki@yahoo.com
Copyright © 2013 Mary Omwamba et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received May 6th, 2013; revised June 6th, 2013; accepted June 14th, 2013
Keywords: Oxidative Stress; Roasted Barley Grain; Antioxidant Activity; Aged Mouse Model
ABSTRACT
The antioxidant activity of extract from roasted barley grain was evaluated by various methods in vitro and in vivo. Results showed that the extract exhibited high antioxidant activities in vitro and in vivo, evidenced by its ability to chelate ferrous ions, scavenge hydroxyl and superoxide radicals, and prevent lipid peroxidation of liver homogenate. The extract significantly increased the total antioxidant capability (T-AOC) in aged mice (P < 0.05). The activities of antioxidant enzymes superoxide dismutase (SOD) and glutathione peroxidase (GSH-Px) increased while levels of malondialodehyde (MDA) and manoamine oxidase (MAO) decreased in both the liver and brain of aged mice treated with the extract compared to the control (untreated mice). The results demonstrate potential antioxidant activities and antiaging effect of roasted barley grain. This provides scientific support for the use of roasted barley grain as an antioxidant against oxidative stress.
1. Introduction
The recent growth in knowledge of reactive oxygen species (ROS) and antioxidants in biology is producing a medical revolution that promises a new age of health. ROS are increasingly recognized as the pathogenesis of certain human diseases, including cancer, ageing and chronic arterial disease [1]. Living tissues are endowed with indigenous antioxidant defense mechanisms, including the enzymes catalase, superoxide dismutase, glutathione peroxidase and glutathione-S-transferase. A reduction in the activity of these enzymes is associated with the accumulation of highly reactive free radicals, leading to deleterious effects such as loss of integrity and function of cell membranes. The role of an antioxidant is to remove free radicals. The mechanism for antioxidants to remove free radical involves donating hydrogen to a free radical and hence its reduction to a non reactive species through removing the odd electron feature which is responsible for radical reactivity [2].
Oxidative stress is a process where the physiological balance between pro-oxidants and antioxidants is disrupted in favors of the former, ensuing in potential damage for the organism [3]. Dietary antioxidant intake may be an important strategy for inhibiting or delaying the oxidation of susceptible cellular substrates, and is thus relevant to disease prevention in many paradigms. There is growing interest in barley products because of their high content of phenolic compounds such as phenolic acids (benzoic and cinnamic acid derivatives), proanthocyanidins, tannins, flavonols, chalcones, flavones, flavanones, and amino phenolic compounds [4]. In our previous study, we reported that roasting had significant influences on total phenolic contents as well as antioxidant activity in barley. The contents of phenolic compounds and antioxidant activity as measured by DPPH radical scavenging activity, ferric thiocyanate method and reducing power assay, increased significantly in RBG. Phenolic acids were identified as the major phenol compounds in the extract [5,6].
Phenolic compounds such as flavonoids, phenolic acids, diterpenes and tannins have received attention for their high antioxidative activity [7]. Preliminary results suggest that these phenolic acids are absorbed in humans [8,9] and that their antioxidant activity may reduce the risk of coronary heart diseases, cancers, and the aging processes [10]. It has been reported that barley including many phenol compounds such as vanillic acid, p-coumaric acid, p-hydroxylbenzoic acid, p-hydroxybenzal-dehyde, quercetin and 3,4-dihydroxybenzaldehyde has an effect on scavenging ROS [11].
Although in vitro antioxidant activity of RBG has been studied [5,11,12], no literature documenting the antioxidant role of hot air RBG in vivo is available. The objecttive of the present study was to evaluate the antioxidant effect of extract from RBG towards oxidative stress in vitro and in vivo. The in vitro antioxidant activity of RBG was determined by measuring the scavenging effects on hydroxyl and superoxide radicals as well as chelation of Fe2+ and lipid peroxidation of liver homogenate. In vivo evaluation was based on the aged mice model.
2. Materials and Methods
2.1. Chemicals and Reagents
The total antioxidant capacity (T-AOC), superoxide dismutase (SOD), malondialdehyde (MDA), maloamine oxide (MAO) and glutathione peroxidase (GSH-Px) comercial kits were purchased from Jiancheng institute of biotechnology, China. Folin-Ciocalteu phenol reagent, gallic acid, 300 g/l hydrogen peroxide, sodium salicylate, pyrogallol, 2-thiobarbituric acid and other chemicals used were of high quality and commercially obtained from local suppliers in Nanjing, China.
2.2. Sample Preparation
Roasted barley grains were prepared according to the condition that optimizes antioxidant activity as reported in our earlier study [5]. Flour from RBG was extracted in 800 g/kg ethanol at a ratio of 1:10 w/v. After 2 h of extraction in a shaking water bath at 45˚C under dark conditions, the supernatant and the residue were separated by vacuum filtration. The residue was then re-extracted as in the first extraction and the extraction solutions combined. The obtained extraction solutions were concentrated in a rotary vacuum evaporator at 45˚C, lyophilized and stored at −20˚C. Before the assays, the extract was dissolved at required concentrations.
2.3. Animal Treatment
Thirty six aged (24 months) and 12 young (3 months) male Kunming mice were purchased from the experimental animal center of Shanghai. The mice were housed in a temperature (25˚C ± 2˚C) and humidity (50% ± 5%) controlled room with a 12 h light-dark cycle and free access to standard pellet diet and drinking water. Animals were maintained per national guidelines and protocols approved by the Institutional Animal Ethical Committee. After an adaptation period of 6 days, the aged mice were randomly divided into four groups of 12 mice each. Control group (group AC) mice were introgastric gavage (IG) administered with 0.2 mL of 9 g/l saline only; the RBG extract treatment group (group RBG 100 and 200) mice were IG administrated RBG extract (100 and 200 mg/kg, dispersed in 9 g/l saline respectively). The young mice were used as the normal control group (group NC) and were IG administered with 0.2 mL of 9 g/l saline only [13]. The IG administration in each group was done daily for 60 days. Body weights were recorded weekly during the 60 day experiment. At the end of the experimental period, all the animals were sacrificed by decapitation. From each animal, 5ml of blood samples were collected and serum separated after centrifuging at 10,000 rpm for 20 min at 4˚C. The liver and total brain were excised. The serum, total brain and liver samples were stored at −80˚C until analysis.
2.4. Determination of Total Phenol Content (TPC)
Total phenolic content of the extracts was determined according to the Folin-Ciocalteu spectrophotometric method [14] with slight modifications. Briefly, 0.5 mL diluted barley extract solution was mixed with 2.5 ml of tenfold diluted Folin-Ciocalteu’s phenol reagent and allowed to react for 5 min. Then 2 mL of 75 g/l sodium carbonate (Na2CO3) solution was added, and the final volume made up to 10 mL with deionized water. After reaction for 1 h at room temperature, the absorbance at 760 nm was determined. The calibration curve was obtained with gallic acid, and the results expressed as mg of gallic acid equivalents per gram of dry weight (mg GAE/g dry weight (DW)).
2.5. Fe2+-Chelating Activity Assay
The extract was assessed for its ability to compete with ferrozine for iron (II) ions in free solution. The Fe2+- chelating activity was measured according to a literature procedure by Hsu et al. [15] with a few modifications. The reaction mixture (2.15 ml) contained 500 μl roasted barley grain (0 - 10 g/L), 50 μl FeCl2 (2 mmol/l) and 1.6 ml deionised water. The mixture was shaken vigorously and left at room temperature for 5 min; 100 μl of ferrozine (5 mmol/l in methanol) were then added, mixed and left for another 5 min to complex the residual Fe2+. The absorbance of the Fe2+-ferrozine complex was measured at 562 nm against a blank performed in the same way using FeCl2 and water. EDTA was used as the standard solution. All tests were run in triplicate and averaged. The chelating activity of the extract for Fe2+ was calculated as;
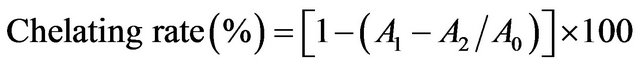
where A0 was the absorbance of the control (without extract) and A1 was the absorbance in the presence of the extract or standard solution. A2 was the absorbance without ferrozine.
2.6. Hydroxyl Radical Scavenging Assay
Hydroxyl radical (OH·) scavenging effect was measured according to the method of Wang et al. [2] OH radicals were generated by the Fenton reaction in the system from FeSO4 and H2O2, and detected by their ability to hydroxylate salicylate. The reaction mixture contained 0.5 ml FeSO4 (8 mM), 0.8 ml H2O2 (6 mM), 0.5 ml distilled water, 1.0 ml sample of various concentrations and 0.2 ml sodium salicylate (20 mM). After reaction for 30 min at 37˚C, the absorbance of the hydroxylated salicylate complex was measured at 562 nm. Trolox was used as the positive control. The percent scavenging rate was obtained according to the formula:
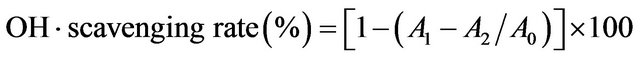
where A0 was the absorbance of the control (without extract), A1 was the absorbance with extract addition and A2 was the absorbance without sodium salicylate.
2.7. Superoxide Radical Scavenging Assay
Superoxide radical 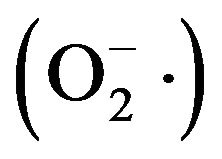 scavenging effect of RBG extract was examined by a pyrogallol autoxidation system as reported by Su et al. [16] with minor modifications. The reaction mixture contained 0.2 mL of different concentrations of extract added to 5.7 mL of 50 mM Tris-HCl buffer (pH 8.2). The mixture was incubated at 25˚C for 10 min and then 0.1 ml of 6 mM pyrogallol (25˚C) was added. The absorbance of the reaction mixture was measured at 320 nm every 30 s until the reaction proceeded to 5 min. Trolox at the same concentration as sample was used as the positive control. The scavenging rate was obtained according to the following equation
scavenging effect of RBG extract was examined by a pyrogallol autoxidation system as reported by Su et al. [16] with minor modifications. The reaction mixture contained 0.2 mL of different concentrations of extract added to 5.7 mL of 50 mM Tris-HCl buffer (pH 8.2). The mixture was incubated at 25˚C for 10 min and then 0.1 ml of 6 mM pyrogallol (25˚C) was added. The absorbance of the reaction mixture was measured at 320 nm every 30 s until the reaction proceeded to 5 min. Trolox at the same concentration as sample was used as the positive control. The scavenging rate was obtained according to the following equation

where A0 was the absorbance of the supernatant without extract, A1 was the absorbance of the extract addition and A2 was the absorbance of extract solution.
2.8. Inhibition of Lipid Peroxidation in Mice Liver Homogenate
The inhibition effect of RBG on lipid peroxidation of mice liver homogenate was measured according to the thiobabituric acid method. FeCl2-H2O2 was used to induce liver homogenate peroxidation. One percent liver homogenate was prepared and the lipid peroxidation assay was conducted according to Yen and Hsieh [17] with slight modification. Briefly, 0.2 ml of different concentrations of RBG extract (0 - 0.5 mg/ml) was mixed with 1.0 ml 10 g/kg liver homogenate, 50 µl FeCl2 (0.5 mM) and H2O2 (0.5 mM). The mixture was incubated at 37˚C for 1 h. Then 1.0 mL TCA (1.5 g/kg) and thiobabituric acid (6.7 g/kg) were added. The mixture was heated in boiling water for 15 min. The absorbance was recorded at 532 nm. Trolox was used as a positive control. The percentage inhibition was calculated as follows;
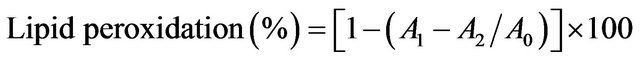
where A0 was absorbance of control (without extract), A1 was absorbance of extract addition and A2 was absorbance without liver homogenate.
2.9. Measurement of T-AOC in Mice Blood
At the end of feeding treatment, mice were decapitated. Blood was collected and centrifuged at 10,000 rpm for 20 min at 4˚C and the serum separated. The total antioxidant capability (T-AOC) was examined by a commercial kit (Nanjing Jiancheng Bioengineering Institute, Nanjing City, PR China) following the manufacturer’s protocol. In the reaction mixture ferric ion was reduced by antioxidant reducing agents and blue complex Fe2+-TPTZ (2,4,6-tri(2-pyridyl)-s-triazine) was produced. The optical density was measured at 520 nm. One unit (U) of TAOC was defined as the amount that increased the absorbance by 0.01 at 37˚C. Data were expressed as U/ml blood.
2.10. Measurement of SOD, GSH-Px, MDA and MAO Activity in the Brain and Liver
After mice were sacrificed, their brain and liver was removed rapidly and homogenized in ice-cold physiological saline solution (9 g/l). The homogenate was centrifuged at 10,000 rpm at 4˚C for 20 min, and the supernatant collected for analysis. The determination of the activity of superoxide dismutase (SOD), glutathione peroxide (GSH-Px), malondialdehyde (MDA) and monoamine oxidase (MAO) were performed using commercial kits (Nanjing Jiancheng Bioengineering Institute, Nanjing City, PR China).The assay for total SOD was based on its ability to inhibit the oxidation of oxymine by the xanthine-xanthine oxidase system. One unit (U) of SOD activity was defined as the amount that reduced the absorbance at 550 nm by 50%, and data were expressed as U/mg protein. The activity of GSH-Px was determined by quantifying the catalyzed reaction rate of GSH per minute on the base of its catalysis. One unit of GPx activity was defined as 1.0 µM GSH oxidized to GSSG per milligram of proterin per 5 min at 37˚C after exclusion of non enzymatic reaction. MDA content was determined by thiobarbituric acid reaction (TBAR) method and expressed as nanomoles per milligram of brain or liver protein. Ultraviolet chromatometry was used for MAO activity determination. One unit (U) of MAO activity was defined as the amount that increased the absorbance by 0.01 at 37˚C, and the MAO activity was expressed as U/h/mg protein.
2.11. Protein Assay
Protein concentration was measured by the Lowry method [18]. Bovine serum albumin (BSA) was used as the standard. The principle behind the Lowry method of determining protein concentrations lies in the reactivity of the peptide nitrogen(s) with copper (II) ions under alkaline conditions and the subsequent reduction of the FolinCiocalteau phosphomolybdic phosphotungstic acid to heteropolymolybdenum blue by the copper-catalyzed oxidation of aromatic acids.
2.12. Statistical Analysis
The experimental results were expressed as means ± standard deviation (SD). SAS version 9 was used to carry out the analysis of variance (ANOVA), P < 0.05 values were regarded as significant.
3. Results
3.1. Fe2+-Chelating Activity Assay
To better estimate the antioxidant potential of roasted barley grain extract, its chelating activity was evaluated against Fe2+. The chelating effects of the extract and EDTA on ferrous ions increased with increasing concentrations (Figure 1). In concentrations of 10 mg/ml, the barley grain extract as well as EDTA exhibited chelating effects of 52.6% and 80.5%, respectively. The concentration of the extract and EDTA needed for 50% chelation of ferrous ions (EC50) was 7.3 and 3.9 mg/ml, respectively.
3.2. OH· Scavenging Activity
In this study the hydroxyl radical scavenging effect of roasted barley grain extract was concentration-dependent and at concentration of 3 mg/ml the scavenging effect was found to be 69.56% (Figure 2). Trolox served as a positive control was found to have 93.01% scavenging effect at the same concentration. Although the scavenging effect of roasted barley grain extract was lower than that of trolox, the result showed that the extract had strong hydroxyl radical scavenging effect with the half inhibition concentration (EC50) of 0.56 ± 0.05 mg/ml. Trolox had an excellent scavenging activity on OH· with the EC50 of 0.03 ± 0.01 mg/ml.
3.3. Superoxide Radical Scavenging Activity
Figure 3 shows the percentage inhibition rate of superoxide radical generation by ethanolic extract of roasted barley grain. The decrease in absorbance at 560 nm with antioxidants indicates the consumption of superoxide anion in the reaction mixture. The inhibition activity of roasted barley grain extract was higher with EC50 of 3.26 ± 0.15 mg/ml compared to trolox with EC50 of 3.59 ± 0.13 mg/mL which was significantly different at P < 0.05. At the concentration of above 3 mg/mL, trolox had a
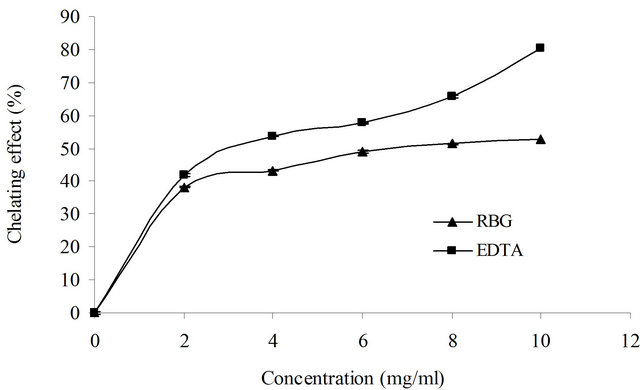
Figure 1. Chelating effect of roasted barley grain (RBG) extract on ferrous ions compared to that of ethylene diaminetetra acetic acid (EDTA). Values are means ± SD (n = 3).
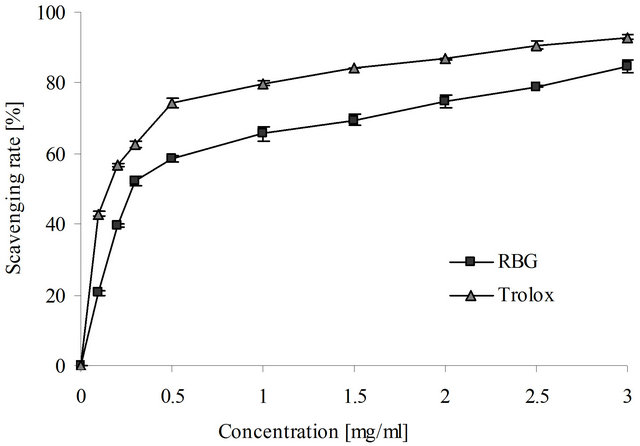
Figure 2. The hydroxyl radical scavenging capacity of roasted barley grain (RBG) extract and Trolox. Values are means ± SD (n = 3).

Figure 3. The superoxide radical scavenging capacity of roasted barley grain extract and Trolox. Values are means ± SD of three determinations.
better inhibition activity compared to barley extract.
3.4. Inhibition of Lipid Peroxidation
In this study, we measured the potential of roasted barley grain extract to inhibit lipid peroxidation in mice liver homogenate, induced by the FeCl2-H2O2 system. The end products of the process were measured in terms of thiobarbituric acid-reactive substances (TBARS) formed. Both roasted barley grain extract and trolox inhibited lipid peroxidation in a concentration-dependent manner (Figure 4). At a concentration of 500 μg/ml the inhibition of lipid peroxidation by roasted barley grain extract was 66.3% while that of trolox was 94.8%.
3.5. Antioxidant Effect of Roasted Barley Grain Extract in Aged Mice
Compared to AC group, total antioxidant capacity (TAOC) was found to be significantly higher for RBG 200 group (P < 0.01) and RBG 100 group (P < 0.05) mice. The increase was by 49.0% and 13.8% respectively. The mean activity of antioxidant enzymes SOD and GSH-Px was significantly higher in the liver and brain tissues homogenate of mice treated with 200 mg/kg roasted barley grain extract compared to AC mice group (P < 0.01) (Table 1). In both the brain and liver of RBG 100 mice group the activity of SOD increased significantly (P < 0.01). The mean concentration of MDA and MAO levels in the brain and liver homogenate decreased significantly in both RBG 200 and RBG 100 mice group (P < 0.01) except for MAO in the liver of RBG 100 mice group which was significantly lower at P < 0.05 when compared to AC group.
When the brain and liver of AC group was compared with the NC group, the activity of antioxidant enzymes SOD and GSH-Px were significantly lower (P < 0.01) in AC group while the levels of MDA and MAO were significantly higher (P < 0.01) except for the MAO level in the brain which was significantly higher at P < 0.05 (Table 1).
4. Discussion
The investigations on antiradical and antioxidant activeties of phenolic compounds, including flavonoids and phenolic acids have been reported [19]. Studies have shown that the polyphenols found in dietary and medicinal plants could inhibit oxidative stress by antioxidant mechanisms [20]. Iron and copper are essential transition metal elements in the human body required for the activity of a large range of enzymes and for some proteins involved in cellular respiration, O2 transport and redox reactions. Unfortunately, they contain unpaired electrons that enable them to participate in one-electron transfer reactions. Hence, they are powerful catalysts of autoxidation reactions [21]. Roasted barley grain chelated Fe2+ by 52.6% at 10 mg/ml. The chelating effect of the extract can inhibit generation of radicals, consequently reducing free radical-induced damage. 3,4-dihydroxybenzaldehyde purified from barley grain was found to chelated Fe2+ by 7% at 0.32 μg/ml, 11% at 1.6 μg/ml, 12% at 8 μg/ml, 56% at 40 μg/ml and 70% at 200 μg/ml, respectively [22].
Hydroxyl radical scavenging is an important antioxidant activity because of very high reactivity of the hydroxyl radical which enables it to react with a wide range of molecules found in living cells such as sugars, amino acids, lipids and nucleotides. Although OH formation can occur in several ways, by far the most important mechanism in vivo is the Fenton reaction, where a transition metal is involved as a pro-oxidant in the catalyzed decomposition of superoxide and hydrogen peroxide [23].
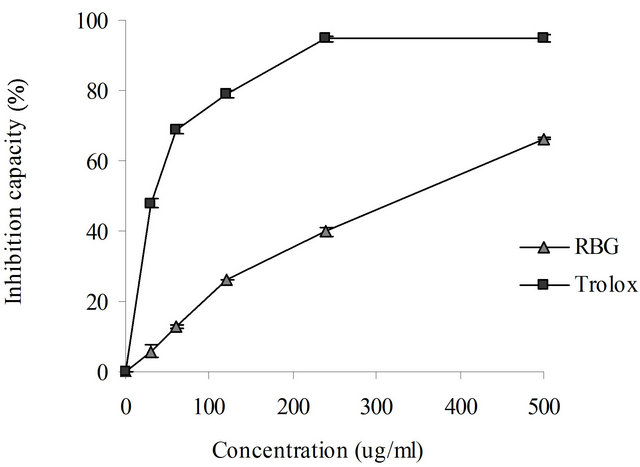
Figure 4. Inhibition of lipid peroxidation by roasted barley grain extract and Trolox in rat liver homogenate. Each value represents mean ± SD (n = 6).
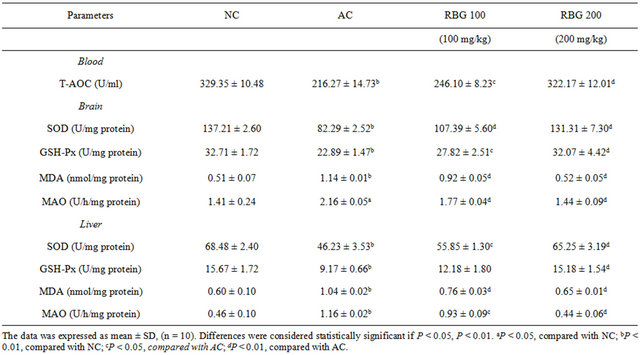
Table 1. Effect of roasted barley grain (RBG) extract on activities of SOD, GSH-Px, MAO, T-AOC and level of MDA in aged mice.
The high hydroxyl radical scavenging activity of the extract may be attributed to the phenol compounds in the extract especially p-hydroxybenzaldehyde and 3,4-dihydroxybenzaldehyde which contain phenolic hydroxyl group with the ability to accept electrons. These electrons can combine with free radicals competitively to decrease hydroxyl radical and intracellular ROS [23,24].
Superoxide radical is the most common free radical generated in vivo. Pyrogallic acid can autooxidise in slightly alkaline conditions to produce superoxide radical. The constant rate of this autooxidation reaction is dependent on the pyrogallic acid concentration. Antioxidants can inhibit the spontaneity oxidation of pyrogallol but it has been found that the reaction is stable in 1 - 3 min. Repetitious tests with roasted barley grain extract showed the reaction was stable in 4 - 5 min and conesquently the data at 5 min was adopted to evaluate the inhibition activity. At lower concentrations roasted barley grain extract inhibited superoxide radical in the same way as trolox given the low values of EC50 obtained. Superoxide radical is known to be very harmful to cellular components as a precursor of the more reactive oxygen species, contributing to tissue damage and various diseases [25].
Lipid peroxidation, a process induced by free radicals, leads to oxidative deterioration of polyunsaturated lipids. In biological systems, lipid peroxidation inactivates cellular components and therein plays a key role in oxidative stress in biological systems by cell membrane disruption and cell damage [26]. Due to depletion in antioxidant levels, the free radicals are not neutralized and aged organs show enhanced susceptibility to lipid peroxidation. Decrease in lipid peroxidation by roasted barley grain may be a result of it scavenging hydroxyl radical produced by FeCl2-H2O2 and H2O2 in the reaction system. On the other hand, polyphenols in roasted barley grain may contain phenolic hydroxyl group, with the ability to accept electrons, which can combine with free radical competitively to decrease lipid peroxidation induced by free radicals.
Barley polyphenol, and grain in general, provides a myriad of antioxidative mechanism which may be partially explained by the fact that it acts as electron donors reducing the oxidized intermediates of lipid peroxidation processes [27,28], the ability to scavenge superoxide anion [28], a common action among other form of polyphenols [29] and the ability to chelate ferrous ions [27]. 3,4-dihydroxybenzaldehyde, a natural phenolic compound purified from barley seeds, may exert the inhibitory effect on H2O2-induced tumor development by blocking H2O2-induced oxidative DNA damage, cell death and apoptosis [22].
A vast number of evidence implicates that aging is associated with a decrease in antioxidant status and that age-dependent increases in lipid peroxidation are a consequence of diminished antioxidant protection [30], being in agreement with our current study. The major antioxidant enzymes, including SOD, GPX and CAT, are regarded as the first line of the antioxidant defense system against reactive oxygen species generated in vivo during oxidative stress. SOD dismutates superoxide radicals to form hydrogen peroxide, which in turn is decomposed to water and oxygen by GSH-Px and CAT, thereby preventing the formation of hydroxyl radicals [31].
Research shows that diets rich in polyphenols increased the antioxidative capabilities of serum GPx and SOD [32,33]. This study reveals that roasted barley grain extract significantly increased the antioxidant activity of enzymes SOD and GSH-Px in liver and brain homogenates of aged mice. This is presumed to be the potential effect of the antioxidative nature of barley’s phenolic compounds. The antioxidative effect action of roasted barley grain was either by having a more direct effect in reducing ROS or by maintaining enzymatic activities. In both liver and brain, significant increase in SOD and GSH-Px activity and low levels of MDA and MAO were found in all barley grain extract treated groups compared to the AC group. Roasted barley grain extract could thus increase the activity of SOD and GSH-Px and decrease the level of MDA and MAO (Table 1). Change in enzymatic activity with aging has previously been reported [34]. Glutathione protects vulnerable cells from oxidative and other metabolite stress due to the metabolism of endogenous as well as exogenous substrates. Therefore the change in GSH-Px activity may play a significant role in the aging process. It has been proposed in literature that any significant increase in SOD activity must be accompanied with comparable increase in GSH-Px activities to prevent excessive buildup of hydrogen peroxide [35].
REFERENCES
- J. Moskovitz, M. B. Yim and P. B. Chock, “Free Radicals and Disease,” Archives of Biochemistry and Biophysics, Vol. 397, No. 2, 2002, pp. 354-359. doi:10.1006/abbi.2001.2692
- H. Wang, X. D. Gao, G. C. Zhou, L. Cai and W. B. Yao, “In Vitro and in Vivo Antioxidant Activity of Aqueous Extract from Choerospondias axillaris Fruit,” Food Chemistry, Vol. 106, No. 3, 2008, pp. 888-895. doi:10.1016/j.foodchem.2007.05.068
- B. Halliwell and J. M. C. Gutteridge, “Role of Free Radicals and Catalytic Metal Ions in Human Disease: An Overview,” Methods of Enzymology, Vol. 186, 1990, pp. 1-85. doi:10.1016/0076-6879(90)86093-B
- A. K. Holtekjolen, A. B. Baevere, M. Rodbotten, H. Berg and S. H. Knutsen, “Antioxidant Properties and Sensory Profiles of Breads Containing Barley Flour,” Food Chemistry, Vol. 110, No. 2, 2008, pp. 414-421. doi:10.1016/j.foodchem.2008.02.054
- M. Omwamba and Q. Hu, “Antioxidant Capacity and Antioxidative Compounds in Barley (Hordeum vulgare L.) Grain Optimized Using Response Surface Methodology in Hot Air Roasting,” European Food Research and Technology, Vol. 229, No. 6, 2009, pp. 907-914. doi:10.1007/s00217-009-1128-7
- M. Omwamba and Q. Hu, “Antioxidant Activity in Barley (Hordeum Vulgare L.) Grains Roasted in a Microwave Oven under Conditions Optimized Using Response Surface Methodology,” Journal of Food Science, Vol. 75, No. 1, 2010, pp. C66-C77. doi:10.1111/j.1750-3841.2009.01426.x
- C. A. Rice-Evans, N. J. Miller and G. Paganga, “StructureAntioxidant Activity Relationships of Flavonoids and Phenolic Acids,” Free Radical Biology and Medicine, Vol. 20, No. 7, 1996, pp. 933-956. doi:10.1016/0891-5849(95)02227-9
- A. R. Rechner, G. Kuhnle, P. Bremner, G. P. Hubbard, K. P. Moore and C. A. Rice-Evans, “The Metabolic Fate of Dietary Polyphenols in Humans,” Free Radical Biology and Medicine, Vol. 33, No. 2, 2002, pp. 220-235.
- A. R. Rechner, M. A. Smith, G. Kuhnle, G. R. Gibson, E. S. Debnam, S. K. S. Srai, K. P. Moore and C. A. RiceEvans, “Colonic Metabolism of Dietary Polyphenols: Influence of Structure on Microbial Fermentation Products,” Free Radical Biology and Medicine, Vol. 36, No. 2, 2004, pp. 212-225.
- L. Yu and K. Zhou, “Antioxidant Properties of Bran Extracts from ‘Platte’ Wheat Grown at Different Locations,” Food Chemistry, Vol. 90, No. 1-2, 2005, pp. 311-316. doi:10.1016/j.foodchem.2004.04.007
- H. Etoh, K. Murakami, T. Yogoh, H. Ishikawa, Y. Fukuyama and H. Tanaka, “Anti-Oxidative Compounds in Barley Tea,” Bioscience, Biotechnology, and Biochemistry, Vol. 68, No. 12, 2004, pp. 2616-2618. doi:10.1271/bbb.68.2616
- P. Duh, G. Yen, W. Yen and L. Chang, “Antioxidant Effect of Water Extracts from Barley Prepared under Different Roasting Temperatures,” Journal of Agriculture and Food Chemistry, Vol. 49, No. 3, 2001, pp. 1455-1462. doi:10.1021/jf000882l
- X. Song, M.M. Bao, D.D. Li and Y.M Li, “Advanced Glycation in D-Galactose Induced Mouse Aging Model,” Mechanisms of Aging and Development, Vol. 108, No. 3, 1999, pp. 239-251. doi:10.1016/S0047-6374(99)00022-6
- V. L. Singleton and J. A. Rossi, “Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents,” American Journal of Enology and Viticulture, Vol. 16, No. 3, 1965, pp. 144-158.
- B. Hsu, I. M. Coupar and K. Ng, “Antioxidant Activity of Hot Water Extract from the Fruit of the Doum Palm, Hyphaene Thebaica,” Food Chemistry, Vol. 98, No. 2, 2006, pp. 317-328. doi:10.1016/j.foodchem.2005.05.077
- X.-Y. Su, Z.-Y. Wang and J.-R. Liu, “In Vitro and in Vivo Antioxidant Activity of Pinus koraiensis Seed Extract Containing Phenolic Compounds,” Food Chemisty, Vol. 117, No. 4, 2009, pp. 681-686. doi:10.1016/j.foodchem.2009.04.076
- G. C. Yen and C. L. Hsieh, “Antioxidant Activity of Extracts from Du-Zhong (Eucommia urmoides) towards Various Peroxidation Models in Vitro,” Journal of Agriculture and Food Chemistry, Vol. 46, No. 10, 1998, pp. 3952-3957. doi:10.1021/jf9800458
- O. H. Lowry, N. J. Rosebrough, A. L. Farr and R. J. Randall, “Protein Measurement with the Folin Phenol Reagent,” Journal of Biology and Chemistry, Vol. 193, No. 1, 1951, pp. 265-275.
- R. Manian, N. Anusuya, P. Siddhuraju and S. Manian, “The Antioxidant Activity and Free Radical Scavenging Potential of Two Different Solvent Extracts of Camellia sinensis (L.) O. Kuntz, Ficus bengalensis L. and Ficus racemosa L.,” Food Chemistry, Vol. 107, No. 3, 2008, pp. 1000-1007. doi:10.1016/j.foodchem.2007.09.008
- C. Manach, A. Scalbert, C. Morand, C. Remesy and L. Jimenez, “Polyphenols: Food Sources and Bioavailability,” American Journal of Clinical Nutrition, Vol. 79, No. 5, 2004, pp. 727-747.
- R. V. Lloyd, P. M. Hanna and R. P. Mason, “The Origin of the Hydroxyl Radical Oxygen in the Fenton Reaction,” Free Radical Biology and Medicine, Vol. 22, No. 5, 1997, pp. 885-888. doi:10.1016/S0891-5849(96)00432-7
- J. B. Jeong, S. C. Hong and H. J. Jeong, “3,4-Dihydroxybenzaldehyde Purified from the Barley Seeds (Hordeum vulgare) Inhibits Oxidative DNA Damage and Apoptosis via Its Antioxidant Activity,” Phytomedicine, Vol. 16, No. 1, 2009, pp. 85-94. doi:10.1016/j.phymed.2008.09.013
- S. J. Stohs and D. Bagchi, “Oxidative Mechanism in the Toxicity of Metal Ions,” Free Radical Biology and Medicine, Vol. 18, No. 2, 1995, pp. 321-336. doi:10.1016/0891-5849(94)00159-H
- S. S. Thomas, H. G. Michaelh and M. A. Jennife, “Antioxidant Properties of Malt Model Systems,” Journal of Agriculture and Food Chemistry, Vol. 53, No. 12, 2005, pp. 4938-4945. doi:10.1021/jf0501600
- B. Halliwell and J. M. C. Gutteridge, “Free Radicals in Biology and Medicine,” Oxford University Press, Oxford, 1999.
- G. Barrera, S. Pizzimenti and M. U. Dianzani, “Lipid Peroxidation: Control of Cell Proliferation, Cell Differentiation and Cell Death,” Molecular Aspects of Medicine, Vol. 29, No. 1-2, 2008, pp. 1-8. doi:10.1016/j.mam.2007.09.012
- G. C. Yen and H. Y. Chen, “Antioxidant Activity of Various Tea Extracts in Relation to Their Antimutagenicity,” Journal of Agriculture and Food Chemistry, Vol. 43, No. 1, 1995, pp. 27-32. doi:10.1021/jf00049a007
- H. Zhao, W. Fan, J. Dong, J. Lu, J. Chin, L. Shan, Y. Lin and W. Kong, “Evaluation of Antioxidant Activities and Total Phenolic Contents of Typical Malting Barley Varieties,” Food Chemistry, Vol. 107, No. 1, 2008, pp. 296- 304. doi:10.1016/j.foodchem.2007.08.018
- A. Kasdallah-Grissa, B. Mornagui, E. Aouani, M. Hammami, M. El May, N. Gharbi, A. Kamoun and S. El-Fazaa, “Resveratrol, a Red Wine Polyphenol, Attenuates Ethanol-Induced Oxidative Stress in Rat Liver,” Life Sciences, Vol. 80, No. 11, 2006, pp. 1033-1039. doi:10.1016/j.lfs.2006.11.044
- S. Casani, R. Gómez-Pastor and E. Matallana, “Antioxidant Compound Supplementation Prevents Oxidative Damage in a Drosophila Model of Parkinson’s Disease,” Free Radical Biology and Medicine, Vol. 61C, 2013, pp. 151-160. doi:10.1016/j.freeradbiomed.2013.03.021
- D. C. Yao, W. B. Shi, Y. L. Gou, X. R. Zhou, Y. A. Tak and Y. K. Zhou, “Fatty Acid-Mediated Intracellular Iron Translocation: A Synergistic Mechanism of Oxidative Injury,” Free Radical Biology and Medicine, Vol. 39, No. 10, 2005, pp. 1385-1398. doi:10.1016/j.freeradbiomed.2005.07.015
- G. Arteel, L. Marsano, C. Mendez, F. Bentley and C. J. McClain, “Advances in Alcoholic Liver Disease,” Best Practice and Research in Clinical Gastroenterology, Vol. 17, No. 4, 2003, pp. 625-647. doi:10.1016/S1521-6918(03)00053-2
- C. Y. Chen, P. E. Milbury, H. K. Kwak, F. W. Collins, P. Samuel and J. B. Blumberg, “Avenanthramides and Phenolic Acids from Oats Are Bioavailable and Act Synergistically with Vitamin C to Enhance Hamster and Human LDL Resistance to Oxidation,” Journal of Nutrition, Vol. 134, No. 6, 2004, pp. 1459-1466.
- H. R. Warner, “Superoxide Dismutase, Aging and Degenerative Disease,” Free Radical Biology and Medicine, Vol. 17, No. 3, 1994, pp. 249-258. doi:10.1016/0891-5849(94)90080-9
- S. Samuhasaneeto, D. Thong-Ngam, O. Kulaputana, D. Suyasunanont and N. Klaikeaw, “Curcumin Decreased Oxidative Stress, Inhibited NF-jB Activation, and Improved Liver Pathology in Ethanol-Induced Liver Injury in Rats,” Journal of Biomedical Biotechnology, Vol. 2009, 2009, Article ID: 981963. doi:10.1155/2009/981963
Abbreviations
EDTA: ethylene diaminetetra acetic acidGSH-Px: glutathione peroxidaseMAO: manoamine oxidaseMDA: malondialodehydeRBG: roasted barley grainROS: reactive oxygen speciesSOD: superoxide dismutaseT-AOC: total antioxidant capacity.
NOTES
*On behalf of my co-authors, I declare no conflicts of interest about the manuscript and no financial arrangement between an author and any company.
#Corresponding author.