 Vol.2, No.3, 262-267 (2010) doi:10.4236/health.2010.23037 Copyright © 2010 SciRes Openly accessible at http://www.scirp.org/journal/HEALTH/ Health Role of Asp37 in metal-binding and conformational change of ciliate Euplotes octocarinatus centrin Wen Liu1, Lian Duan1, Bing Zhao1, Ya-Qin Zhao1, Ai-Hua Liang2, Bin-Sheng Yang1* 1Key Laboratory of Chemical Biology and Molecular Engineering of Ministry of Education, Institute of Molecular Science, Shanxi University, Taiyuan, China; yangbs@sxu.edu.cn 2Key Laboratory of Chemical Biology and Molecular Engineering of Ministry of Education, Institute of Biotechnology, Shanxi Uni- versity, Taiyuan, China Received 24 December 2009; revised 18 January 2010; accepted 21 January 2010. ABSTRACT Centrin is a member of the EF-hand super family that plays critical role in the centrosome dupli- cation and separation. To investigate the role of Asp37 in the process of metal-binding and conformational change of ciliate Euplotes oc- tocarinatus centrin (EoCen), the mutant D37K, in which aspartic acid 37 had been replaced by lysine, was first obtained by the site-directed mutagenesis. Then 2-p-toluidinylnaphthalene-6- sulfonate (TNS) was used as a fluorescence probe to detect the conformational change of the protein. The results show that the metal- binding capability of the site I of EoCen was lost by the mutation of Asp37→Lys. In comparison the Tb3+-saturated EoCen, the hydrophobic surface of D37K, which is exposed by the bind- ing of Tb3+, has shrunk sharply, suggesting that Asp37 plays an important role in maintaining the proper conformation of EoCen in the presence of Tb3+. Meanwhile, the conditional binding constants of TNS with Tb3+-loaded EoCen and D37K were obtained, K(Tb3+-EoCen-TNS)=(7.38 ±0.25)×105 M -1 and K(Tb3+-D37K-TNS)=(1.16± 0.05)×106 M-1. Keywords: Centrin; Aspartic Acid; Tb3+; TNS 1. INTRODUCTION Centrin is a calcium-binding protein of ~20 kDa, and present in both the pericentriolar material and the centri- oles of centrosome. Genetic studies show that centrin is essential to normal cell cycle-dependent duplication and segregation of the microtubule organizing center (MTOC) [1]. The microtubule organizing center (MTOC) is cyto- plasmic organelles, encountered in almost all the eu- karyotic cells and having an important role in the nu- cleation of the microtubules and the regulation of their dynamics [2]. It is fundamental to many cellular proc- esses, including chromosomal segregation, cytokinesis, fertilization, cellular morphogenesis, cell motility, and intracellular trafficking [3]. Much research has focused on these functions, because abnormal centrosome dupli- cation may lead to chromosomal instability and then cancer, an idea supported by discovery of supernumerary abnormal centrosomes in different human tumor cells [4-6]. In addition, centrin forms part of the human het- erotrimetric DNA damage recognition complex required for global genome nucleotide excision repair [7]. Centrin seems to act as a Ca2+ sensor, i.e., in its Ca2+-load form, centrin interacts with specific target protein to modulate the cellular activity. In general, the binding of Ca2+ in- volves a structural rearrangement of the α-helices of the EF-hand pair domain with the consequent exposure of a hydrophobic cleft [8,9]. Conformational changes are intrinsic to the function of a variety of proteins. 2-p-Toluidinylnaphthalene-6-sulfonate (TNS) has been extensively used in the conformational change of centrin induced by metal ions [10-12]. Ciliate Euplotes octocarinatus centrin (EoCen) is a protein of 168 residues, which shares about 60, 62 and 66% sequence identity with human centrin 1, human centrin 2 and human centrin 3, respectively, and shares approximately 50% sequence identity with the well studied EF-hand protein calmodulin (CaM). Like CaM, centrin consists of two independent domains tethered by a flexible linker, each domain comprising a pair of EF-hand motifs of helix-loop-helix that can potentially bind two calcium ions [13]. As show in Figure 1 [14], the first amino acid of Ca2+-binding 12-residue loop is aspartic acid which is highly conserved. Thus, the Asp at the first position of the loop would be expected to be important to the proper conformation and metal binding characteristics of centrin. Lanthanides have been known for their diversity in biological effects, and the application of lanthanides in medicine has high potential. In agriculture, lanthanides  W. Liu et al. / HEALTH 2 (2010) 262-267 Copyright © 2010 SciRes Openly accessible at http://www.scirp.org/journal/HEALTH/ 263 Figure 1. The sequence logo of the EF-hand domain [14]. The residues are colored as follows: D, E, N, red; K, R, blue; S, T, purple; G, green; hydrophobic, yellow. have been used to increase the production of crops and to promote the growth of livestock in China for many years [15]. The molecular mechanism of the biological effects of lanthanides is not totally understood so far. Lanthanide ions (Ln3+) have similar ionic radii and similar coordina- tion properties to Ca2+ [16]. Hence, Tb3+ was usually used to sense properties of Ca2+-binding proteins [17]. EoCen is the first reported by our laboratory [18] (gene register Y18899), which is cloned from Euplotes octoca- rinatus, and the detailed biological function is unclear. In this paper, in order to investigate the importance of the first amino acid (Asp37) of site I of EoCen to the metal- binding and conformation characteristics of this protein, the mutant protein (D37K), with the mutation of Asp37 to Lys, was obtained by the method of site direct mutation. Using TNS as fluorescence probe, the characterization for the binding of Tb3+ to EoCen and the mutant D37K were investigated. 2. MATERIALS AND METHODS 2.1. Reagents and Stock Solutions N-2-hydroxyethylpiperazine-N-2-ethanesulfonic acid (Hepes) (analytical reagent), ampicillin (ultra pure grade), isopro- pyl-β-D-thiogalactoside (IPTG) (ultra pure grade), tryp- tone, and yeast extract were purchased from Bio. Basic Inc. TNS was bought from Sigma. Glutathione Sepharose TM 4B (GST) was purchased from Pharmacia Ltd. Ter- bium oxide was 99.99% and purchased from Hunan in China. The terbium stock solution was prepared by dis- solving weighed Tb4O7 in concentrated hydrochloric acid. The concentration of Tb3+ was standardized by com- plexometric titration with EDTA using xylenol orange as indicator in HAc/NaAc buffer at pH 5.5. The solution of TNS prepared by dissolving weighed samples. 2.2. Protein Expression and Purification Two proteins were used in this study, namely, EoCen (full-length of the wild type EoCen 1M-168Y), D37K (mutant EoCen with Asp37 changing to be Lys 1M- 168Y). The D37K was acquired by polymerase chain reaction technique with p1 (5’-TATTTAAGACCAAC AAAACTGG-3’) and p2 (5’-AATCAAAGGCTTCTTT GATCTC-3’) used as primers. Proteins of EoCen were over-expressed off a PGEX-6p-1 plasmid construct in Escherichia coli BL21 (DE3) induced with isopro- pyl-Dthiogalactopyranoside (IPTG) to yield milligram quantities of the desired protein as reported previously [18]. Briefly, transformed E. coli cells were grown in LB media containing 100 µg/mL ampicillin and incubated at 37 ◦C while monitoring its growth via optical density (OD) measurements at 600 nm. Once OD600 reaches 0.6, a final concentration of 0.5 mM IPTG was added to the culture, and 3 h later, the cells were harvested and frozen. Frozen cells were thawed in PBS buffer and sonicated with a macro probe at mediate power on ice. This solution was centrifuged at 15,000 g for 25 min at 4 ◦C. The su- pernatant was applied to a Glutathione-Sepharose TM 4B column which has been equilibrated with PBS buffer. After the initial purification by washing the supernatant with PBS buffer, prescissor proteinase was added at 4 ◦C for reacting about one night. Primary target proteins were washed out and eluted with PBS buffer. Then the proteins were applied to a superdex 75 column to be further puri- fied. Fractions containing centrin were identified via 15% SDS-PAGE (sodium dodecyl sulfate-polyacrylamide gel electrophoresis). After purification, the proteins were kept in -80 ◦C. The stock protein solutions were con- served in 10 mM Hepes. 2.3. Metal Lon Removal and Protein Concentration To remove contaminating bound cation, the protein sam- ples were ultrafiltrated extensively against 10 mM Hepes, pH 7.4 containing 1 mM EDTA. The protein concentration was measured spectrophotometrically using molar extinc- tion coefficient at 280 nm of 5600 L·mol−1·cm−1. The ex- tinction coefficient of centrin was estimated from the ty- rosine content as previously reported [19]. 2.4. Native- and SDS-PAGE Assays Each protein sample with the same concentration was prepared in Hepes (10 mM, pH 7.4) for this experiment. Polyacrylamide gels contained 390 mM Tris (pH 8.8), 10% ammonium persulfate, 15% acrylamide/bis (29:1), and 0.1% TEMED. Tris-glycine electrophoresis run- ning buffer contained 25 mM Tris (pH 8.3) and 250 mM glycine. All electrophoreses were run at room temperature. Gels were run at a constant current of 11-12 mA for 2 h. Gels were fixed in 50% methanol, 7% acetic acid for 1 h, washed in distilled water for 1 h, stained with Coomassie blue R-250 for 2 h, washed in distilled water for 2 h. For SDS-PAGE, keeping same experimental condi- tions as above described except that 10% sodium dode- cyl sulfate (SDS) was added and running buffer con- tained 25 mM Tris (pH 8.3), 250 mM glycine, 10% SDS was used. 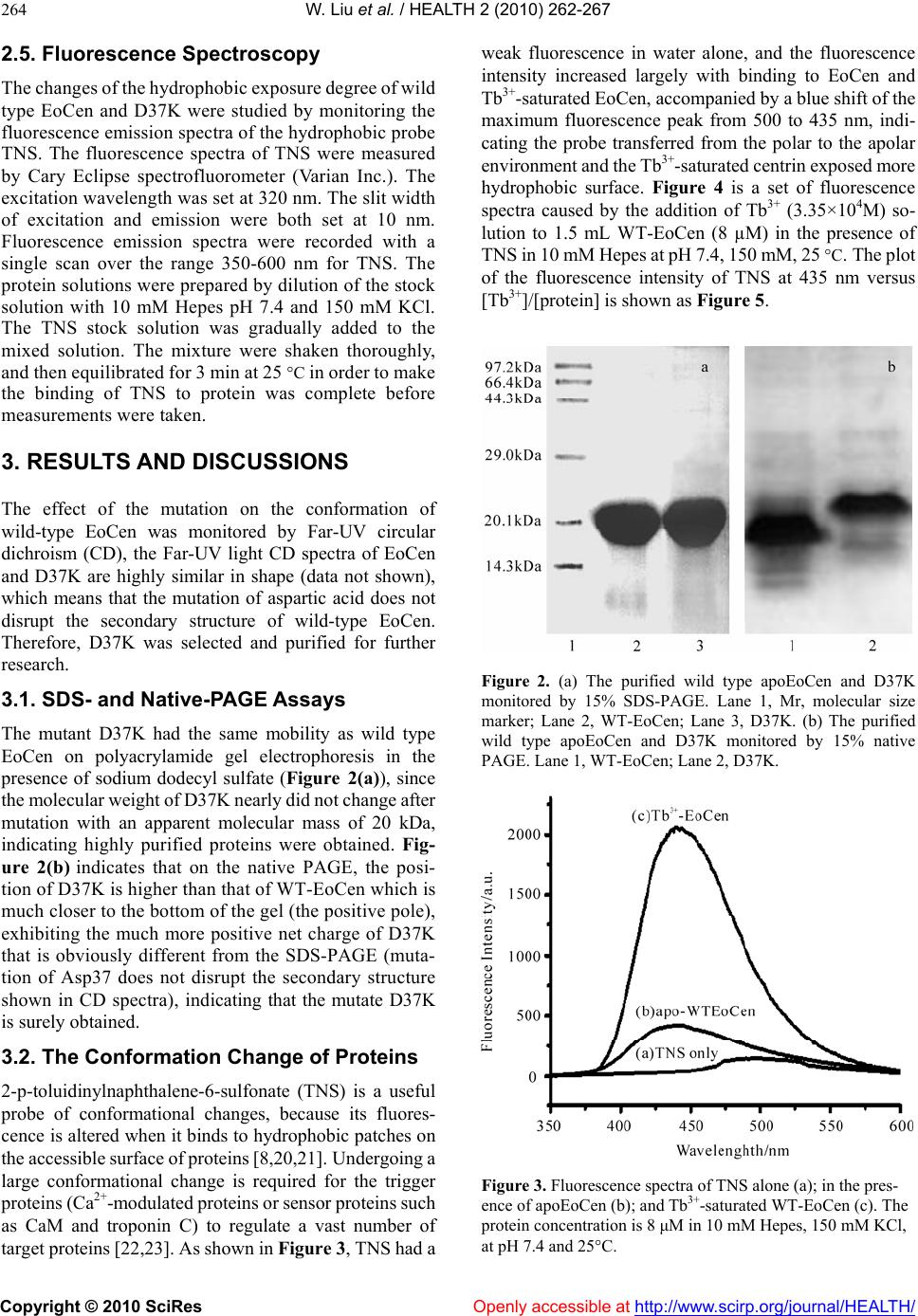 W. Liu et al. / HEALTH 2 (2010) 262-267 Copyright © 2010 SciRes Openly accessible at http://www.scirp.org/journal/HEALTH/ 264 2.5. Fluorescence Spectroscopy The changes of the hydrophobic exposure degree of wild type EoCen and D37K were studied by monitoring the fluorescence emission spectra of the hydrophobic probe TNS. The fluorescence spectra of TNS were measured by Cary Eclipse spectrofluorometer (Varian Inc.). The excitation wavelength was set at 320 nm. The slit width of excitation and emission were both set at 10 nm. Fluorescence emission spectra were recorded with a single scan over the range 350-600 nm for TNS. The protein solutions were prepared by dilution of the stock solution with 10 mM Hepes pH 7.4 and 150 mM KCl. The TNS stock solution was gradually added to the mixed solution. The mixture were shaken thoroughly, and then equilibrated for 3 min at 25 C in order to make the binding of TNS to protein was complete before measurements were taken. 3. RESULTS AND DISCUSSIONS The effect of the mutation on the conformation of wild-type EoCen was monitored by Far-UV circular dichroism (CD), the Far-UV light CD spectra of EoCen and D37K are highly similar in shape (data not shown), which means that the mutation of aspartic acid does not disrupt the secondary structure of wild-type EoCen. Therefore, D37K was selected and purified for further research. 3.1. SDS- and Native-PAGE Assays The mutant D37K had the same mobility as wild type EoCen on polyacrylamide gel electrophoresis in the presence of sodium dodecyl sulfate (Figure 2(a)), since the molecular weight of D37K nearly did not change after mutation with an apparent molecular mass of 20 kDa, indicating highly purified proteins were obtained. Fig- ure 2(b) indicates that on the native PAGE, the posi- tion of D37K is higher than that of WT-EoCen which is much closer to the bottom of the gel (the positive pole), exhibiting the much more positive net charge of D37K that is obviously different from the SDS-PAGE (muta- tion of Asp37 does not disrupt the secondary structure shown in CD spectra), indicating that the mutate D37K is surely obtained. 3.2. The Conformation Change of Proteins 2-p-toluidinylnaphthalene-6-sulfonate (TNS) is a useful probe of conformational changes, because its fluores- cence is altered when it binds to hydrophobic patches on the accessible surface of proteins [8,20,21]. Undergoing a large conformational change is required for the trigger proteins (Ca2+-modulated proteins or sensor proteins such as CaM and troponin C) to regulate a vast number of target proteins [22,23]. As shown in Figure 3, TNS had a weak fluorescence in water alone, and the fluorescence intensity increased largely with binding to EoCen and Tb3+-saturated EoCen, accompanied by a blue shift of the maximum fluorescence peak from 500 to 435 nm, indi- cating the probe transferred from the polar to the apolar environment and the Tb3+-saturated centrin exposed more hydrophobic surface. Figure 4 is a set of fluorescence spectra caused by the addition of Tb3+ (3.35×104M) so- lution to 1.5 mL WT-EoCen (8 µM) in the presence of TNS in 10 mM Hepes at pH 7.4, 150 mM, 25 C. The plot of the fluorescence intensity of TNS at 435 nm versus [Tb3+]/[protein] is shown as Figure 5. Figure 2. (a) The purified wild type apoEoCen and D37K monitored by 15% SDS-PAGE. Lane 1, Mr, molecular size marker; Lane 2, WT-EoCen; Lane 3, D37K. (b) The purified wild type apoEoCen and D37K monitored by 15% native PAGE. Lane 1, WT-EoCen; Lane 2, D37K. Figure 3. Fluorescence spectra of TNS alone (a); in the pres- ence of apoEoCen (b); and Tb3+-saturated WT-EoCen (c). The protein concentration is 8 μM in 10 mM Hepes, 150 mM KCl, at pH 7.4 and 25C. 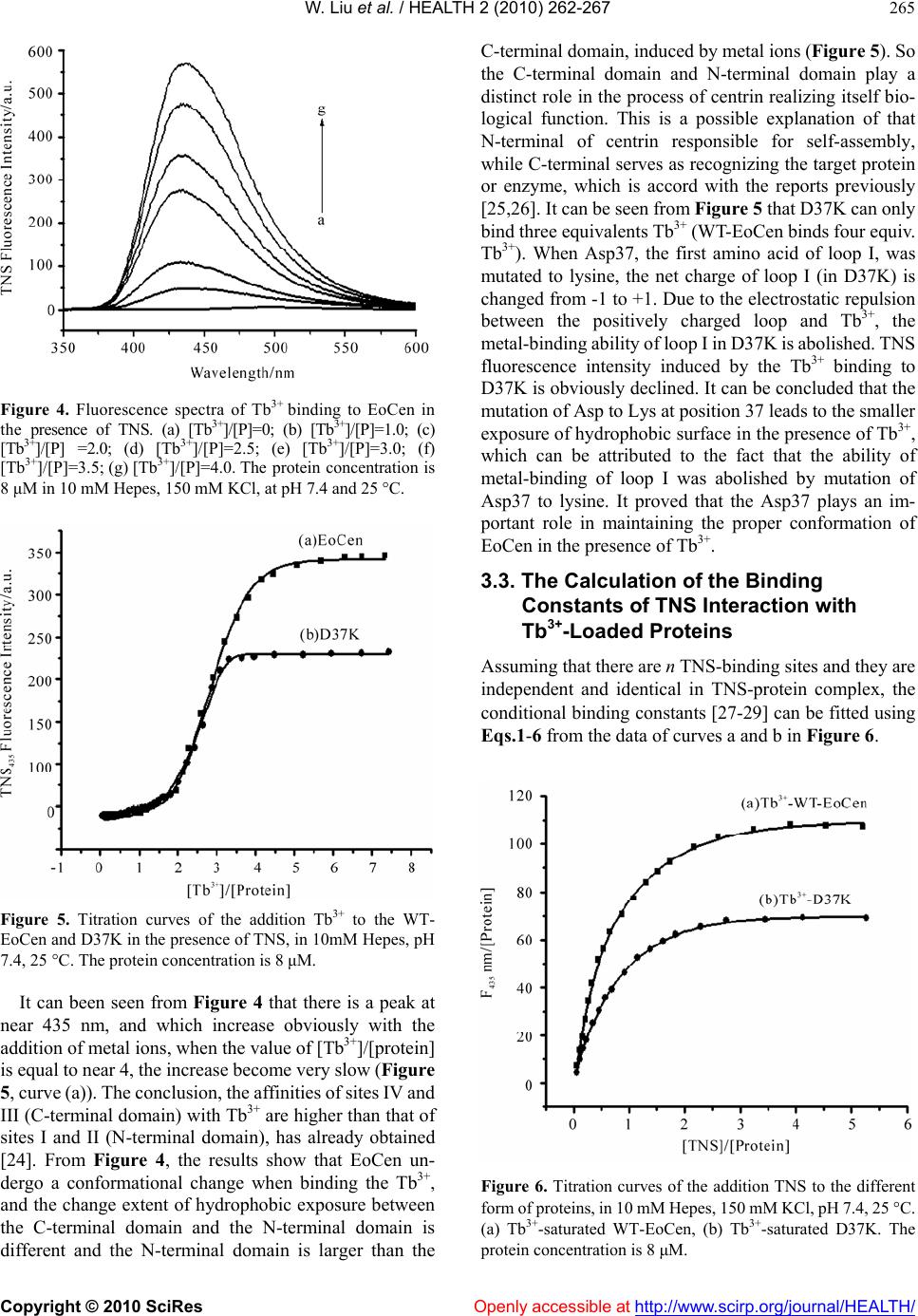 W. Liu et al. / HEALTH 2 (2010) 262-267 Copyright © 2010 SciRes Openly accessible at http://www.scirp.org/journal/HEALTH/ 265 Figure 4. Fluorescence spectra of Tb3+ binding to EoCen in the presence of TNS. (a) [Tb3+]/[P]=0; (b) [Tb3+]/[P]=1.0; (c) [Tb3+]/[P] =2.0; (d) [Tb3+]/[P]=2.5; (e) [Tb3+]/[P]=3.0; (f) [Tb3+]/[P]=3.5; (g) [Tb3+]/[P]=4.0. The protein concentration is 8 μM in 10 mM Hepes, 150 mM KCl, at pH 7.4 and 25 C. Figure 5. Titration curves of the addition Tb3+ to the WT- EoCen and D37K in the presence of TNS, in 10mM Hepes, pH 7.4, 25 C. The protein concentration is 8 μM. It can been seen from Figure 4 that there is a peak at near 435 nm, and which increase obviously with the addition of metal ions, when the value of [Tb3+]/[protein] is equal to near 4, the increase become very slow (Figure 5, curve (a)). The conclusion, the affinities of sites IV and III (C-terminal domain) with Tb3+ are higher than that of sites I and II (N-terminal domain), has already obtained [24]. From Figure 4, the results show that EoCen un- dergo a conformational change when binding the Tb3+, and the change extent of hydrophobic exposure between the C-terminal domain and the N-terminal domain is different and the N-terminal domain is larger than the C-terminal domain, induced by metal ions (Figure 5). So the C-terminal domain and N-terminal domain play a distinct role in the process of centrin realizing itself bio- logical function. This is a possible explanation of that N-terminal of centrin responsible for self-assembly, while C-terminal serves as recognizing the target protein or enzyme, which is accord with the reports previously [25,26]. It can be seen from Figure 5 that D37K can only bind three equivalents Tb3+ (WT-EoCen binds four equiv. Tb3+). When Asp37, the first amino acid of loop I, was mutated to lysine, the net charge of loop I (in D37K) is changed from -1 to +1. Due to the electrostatic repulsion between the positively charged loop and Tb3+, the metal-binding ability of loop I in D37K is abolished. TNS fluorescence intensity induced by the Tb3+ binding to D37K is obviously declined. It can be concluded that the mutation of Asp to Lys at position 37 leads to the smaller exposure of hydrophobic surface in the presence of Tb3+, which can be attributed to the fact that the ability of metal-binding of loop I was abolished by mutation of Asp37 to lysine. It proved that the Asp37 plays an im- portant role in maintaining the proper conformation of EoCen in the presence of Tb3+. 3.3. The Calculation of the Binding Constants of TNS Interaction with Tb3+-Loaded Proteins Assuming that there are n TNS-binding sites and they are independent and identical in TNS-protein complex, the conditional binding constants [27-29] can be fitted using Eqs.1-6 from the data of curves a and b in Figure 6. Figure 6. Titration curves of the addition TNS to the different form of proteins, in 10 mM Hepes, 150 mM KCl, pH 7.4, 25 C. (a) Tb3+-saturated WT-EoCen, (b) Tb3+-saturated D37K. The protein concentration is 8 μM.  W. Liu et al. / HEALTH 2 (2010) 262-267 Copyright © 2010 SciRes Openly accessible at http://www.scirp.org/journal/HEALTH/ 266 The binding equation is presented by: TNS P TNSP n n (1) Given that F∞ is maximum molar fluorescence inten- sity, Fr is fluorescence intensity of every titration dot. The increase of fluorescence intensity resulted from the binding of TNS to protein. The following equations can be obtained: t PFn (2) r F TNS (3) rt b P TNS Fn F (4) where [TNS]b is the bound concentration of TNS. [P]t and [P]b is the total and bound concentration of protein in 10 mM Hepes, pH 7.4, 25 C, respectively. The binding constant can be given as follows: b f TNS TNS P Kn f (5) [TNS]f and [P]f represent the free concentration of TNS and protein, respectively. Finally, equation can be ex- pressed by Eq.6. t t TNS 1 1 TNS{P TNS}Kn b (6) For Tb3+-EoCen, 1/K can be obtained by the linear slope of the plot of [TNS]t/[TNS]b versus {[Tb3+-EoCen]t- [TNS]b}-1, as showed in Figure 7. n is set to be 1, the intercept of Eq.6 is very close to 1, and can also be seen in Figure 6, n=1 is suitable. K (Tb3+-EoCen-TNS) can be calculated to be (7.38±0.25) ×105M-1 from Figure 7. In the similar way, K (Tb3+-D37K-TNS) can be calcu- lated to be (1.16±0.05) ×106M-1. It can be seen that the affinities of TNS binding to EoCen and D37K are different, K (Tb3+-D37K-TNS)>K (Tb3+-EoCen-TNS), showing that D37K, with more positive charges, has larger electrostatic interaction with TNS than EoCen in the presence of Tb3+ and the mutation of Asp37 influence the hydrophobic and electrostatic environment of WT-EoCen. 4. CONCLUSIONS Using the fluorescence spectroscopy, the interaction of TNS and EoCen has been studied in the presence of Tb3+. WT-EoCen and D37K undergo a different extent con- formation change, and D37K exposes lesser hydrophobic surface in the presence of Tb3+, which can be attributed to the fact that the ability of metal-binding of site I was abolished by the mutation of Asp37 to lysine. It can be concluded that the Asp37 plays an important role in maintaining the proper conformation of EoCen in the Figure 7. The plot of [TNS]t/[TNS]b vs {[WT-EoCen-Tb]t-[TNS]b}−1. presence of Tb3+. In the end, the binding constants of TNS with Tb3+-saturated EoCen and D37K were obtained, K(Tb3+-EoCen-TNS)=(7.38±0.25)×105M-1 and K(Tb3+ -D37K-TNS)=( 1.16±0.05) ×106M-1. 5. ACKNOWLEDGEMENTS We acknowledge to the financial support of this work by the National Natural Science Foundation of P.R. China (Nos. 20771068 and 20901048) and the Natural Science Foundation Shanxi Province (No. 2007011024). REFERENCES [1] Thompson, J.R., Ryan, Z.C., Salisbury, J.L. and Kumar, R. (2006) The structure of the human centrin 2-xeroderma pigmentosum group C protein complex. Journal of Bio- logical Chemistry, 281, 18746-18752. [2] Lingle, W.L., Lutz, W.H., Ingle, J.N., et al. (1998) Cen- trosome hypertrophy in human breast tumors: Implica- tions for genomic stability and cell polarity. Proceedings of the National Academy of Scien ces USA, 95, 2950-2955. [3] Jaspersen, S.L. and Winey, M. (2004) The budding yeast spindle pole body: Structure, duplication, and function. An- nual Review of C ell and D evelopmenta l Biology , 20, 1-28. [4] Lingle, W.L., Barrett, S.L., Negron, V.C., et al. (2002) Centrosome amplification drives chromosomal instability in breast tumor development. Proceedi ngs of t h e Nationa l Academy of Sciences USA, 99, 1978-1983. [5] Lingle, W.L. and Salisbury, J.L. (1999) Alter centrosome structure is associated with abnormal mitoses in human breast tumors. American Journal of Pathology, 155, 1941-1951. [6] Lingle, W.L. and Salisbury, J.L. (2000) The role of the centrosome in the development of malignant tumors. Current Topics in Developmental Biology, 49, 313-329. [7] Yang, A., Miron, S., Mouawad, L., et al. (2006) Flexibility and plasticity of human centrin 2 binding to the xeroderma pigmentosum group C protein (XPC) from nuclear exci-  W. Liu et al. / HEALTH 2 (2010) 262-267 Copyright © 2010 SciRes http://www.scirp.org/journal/HEALTH/Openly accessible at 267 sion repair. Biochemistry, 45, 3653-3663. [8] Li, G.T., Wang, Z.J., Zhao, Y.Q., et al. (2007) The spectral studies on the effect of Glu 101 to the metal binding characteristic of Euplotes octocarinatus centrin. Spectro- chim Acta A, 67, 1189-1193. [9] Wiech, H.B., Geier, M., Paschke, T., et al. (1996) Char- acterization of green alga, yeast, and human centrins. Journal of Biological Chemistry, 271, 22453-22461. [10] Cox, J.A., Tirone, F., Durussel, I., et al. (2005) Calcium and magnesium binding to human centrin 3 and interac- tion with target peptides. Biochemistry, 44, 840-850. [11] Wang, Z.J., Ren, L.X., Zhao, Y.Q., et al. (2007) Metal ions-induced conformational change of P23 by using TNS as fluorescence probe. Spectrochim Acta Part A, 66, 1323-1326. [12] Wang, Z.J., Ren, L.X., Zhao, Y.Q., et al. (2008) Investi- gation on the binding of TNS to centrin: An EF-hand pro- tein. Spectrochim Acta Part A, 70, 892-897. [13] Weber, C.V., Lee, D., Chazin, W.J. and Huang, B. (1994) High level expression in Escherichia coli and characteri- zation of the EF-hand calcium-binding protein caltractin. Journal of Biological Chemistry, 269, 15795-15802. [14] Rigden, D.J. and Galperin, M.Y. (2004) The DxDxDG Motif for calcium binding: Multiple structural contexts and implications for evolution. Journal of Biological Chemistry, 343, 971-984. [15] Hu, J., Jia, X., Li, Q., et al. (2004) Binding of La3+ to calmodulin and its effects on the interaction between calmodulin and calmodulin binding peptide, polistes mastoparan. Biochemistry, 43, 2688-2698. [16] Shannon, R.D. (1976) Revised effective ionic radii and systematic studies of interatomic distances in halides and chalcogenides. Acta Crystallographica A, 32, 751-767. [17] Bai, H.J., Liu, W. and Yang, B.S. (2002) Fluorescence studies on the kinetics of terbium (III) removal from monoterbium transferrins by tartaric acid. Acta Chimica Sinica, 60, 1253-1257. [18] Duan, L., Zhao, Y.Q., Wang, Z.J., et al. (2008) Lutetium (III)-dependent self-assembly study of ciliate Euplotes octocarinatus centrin. Journal of Inorganic Biochemistry, 102, 268-277. [19] Pace, C.N., Vajdos, F., Fee, L., et al. (1995) How to measure and predict the molar absorption coefficient of a protein. Protein Science, 4, 2411-2423. [20] McClure, W.O. and Edelman, G.M. (1966) Fluorescent probes for conformational states of proteins. I. Mechanism of fluorescence of 2-p-Toluidinylnaphthalene-6-sulfonate: A hydrophobic probe. Biochemist ry, 5, 1908-1919. [21] Durussela, I., Blouquitb, Y., Middendorpc, S., et al. (2000) Cation- and peptide-binding properties of human centrin 2. FEBS Letters, 472, 208-212. [22] Ikura, M., Clore, G.M., Gronenborn, A.M., et al. (1992) Solution structure of a calmodulin-target peptide complex by multidimensional NMR. Ba x Scie nce, 256, 632-638. [23] Zhang, M., Tanaka, T. and Ikura, M. (1995) Calcium- induced conformational transition revealed by the solution structure of apo calmodulin. Nature S tructural Biology, 2, 758-767. [24] Wang, Z.J., Zhao, Y.Q., Ren, L.X., et al. (2007) Spectral study on the interaction of ciliate Euplotes octocarinatus centrin and metal ions. Journal of Photochemistry and Photobiology A, 186, 178-186. [25] Hu, H.T. and Chazin, W.J. (2003) Unique features in the C-terminal domain provide caltractin with target speci- ficity. Journal of Biological Chemistry, 330, 473-484. [26] Tourbez, M., Firanescu, C., Yang, A., et al. (2004) Cal- cium-dependent self-assembly of human centrin 2. Journal of Biological Chemistry, 279, 47672-47680. [27] Ren, L. X., Zhao, Y.Q., Feng, J.Y., et al. (2006) Terbium- and calcium-binding properties of N-terminal domain of Euplotes centrin. Chinese Journal of Inorganic Chemistry, 22, 87-90. [28] Yang, B.S., Liu, W., Ren, L.X., et al. (2009) The spectral studies on the metal binding characteristic of N-terminal domain of ciliate Euplotes octocarinatus centrin. Journal of Shanxi University (Nature and Science Edition), 32(4), 572-576. [29] Yang, B.S., Yang, P. and Song, L.H. (1984) The binding of Gd (III) to human serum albumin. Chinese Science Bul- letin, 29, 1502-1505.
|