X. X. Wu et al. / J. Biomedical Science and Engineering 3 (2010) 221-225
Copyright © 2010 SciRes.
225
JBiSE
The most frequent (frequency>0.05) BoLA-DRB3.2
alleles of 835 Holstein dairy cattle re-ported by Sharif et
al. [11] were BoLA-DRB3.2*3(0.0652), *8(0.0079),
*11(0.0874), *16(0.0048), *22(0.2353), *23(0.1017),
*24(0.0922). In the present study, the most frequent al-
leles were BoLA-DRB3.2*3(0.0652), **11(0.0874),
*16(0.0048), *22(0.2353), *23(0.1017), *24(0.0922).
Sharif et al reported that BoLA-DRB3.2 *8 was the most
common allele type in Holstein, However, in this study,
the BoLA-DRB3.2 *16 was at low frequency (0.0048),
while the BoLA-DRB3 *22 was the most common al-
lele (0.2353). Therefore, BoLA polymorphism infor-
mation from the research herd seemed representative of
the regional Holstein population. These differences may
be largely due to the long-term adaptation to different
geographical and climatic conditions. Therefore, BoLA
polymorphism information from the research herd
seemed representative of the regional Holstein popula-
tion. These differences may be largely due to the long-
term conditions.
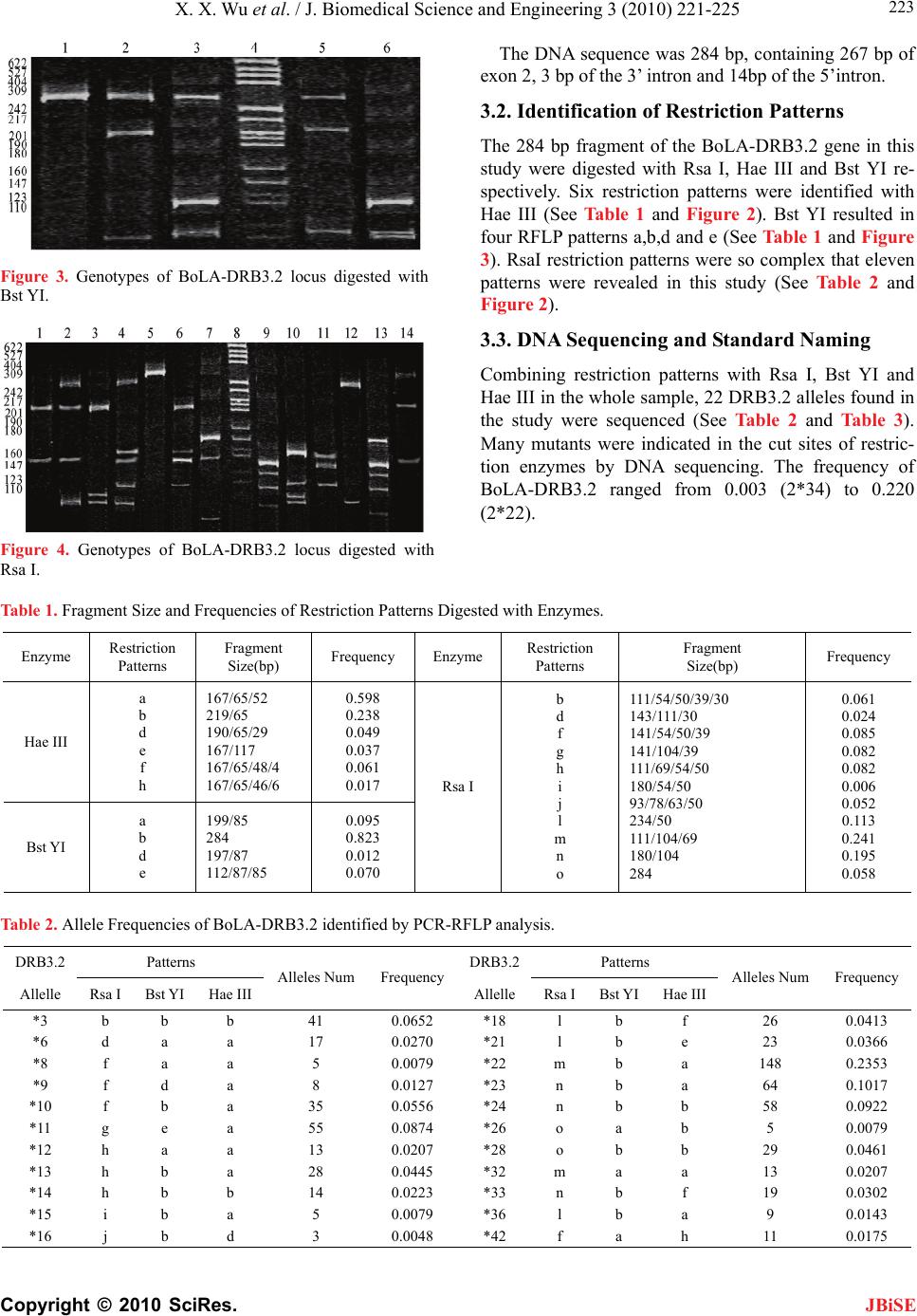
Many mutants, changing the restriction site and the
restriction maps, were identified in the cut sites by DNA
sequencing. RFLP, as a method to identify the polymor-
phism of BoLA-DRB3, can only define the position of
mutants. Other methods must be used to find the detail
mutants. Result of our study demonstrated that the
BoLA-DRB3.2 locus is highly polymorphic in Chinese
Holstein. PCR-RFLP may therefore be a rapid and useful
method for DRB3 typing and studying the evolutionary
changes in dairy cattle.
Sharif reported calving season had no significant ef-
fect on SCC, but this research indicated that calving
season significantly affect SCC; there was a trend that
cows calving in spring and summer have high SCC. Re-
lated to BoLA-DRB3.2 and SCC, the result in this re-
search is different from Sharif et al. [11], among the
most frequently detected alleles, BoLA-DRB3.2*3 has
significant correlation to low SCC, BoLA-DRB3.2*8, 18
and 26 maybe have same function, but their frequencies
are very low.
Contradictory results from different studies invest-
tigating associations between BoLA-DRB3.2 alleles and
mastitis indicate that future studies should focus on as-
sociations of mastitis with BoLA heliotypes rather than
with single BoLA genes.
5. ACKNOWLEDGEMENTS
This The authors appreciate the assistance of the Shanghai Brightlight
Dairy Company. This work was supported by 863 high-tech projects
“The Breeding technology of Multi-locus Combination of Chinese
Holstein” (2008AA10Z144), Natural Science Foundation of Jiangsu
Province, College of major projects (09KJA230002), land 863 Na
tional key project (2008AA101010).
REFERENCES
[1] Andersson, L. and Davies, C.J. (1994) The major histo-
compatibility complex. Cell-Mediated Immunity in
Ruminants, CRC, 37-57.
[2] Davies, C.J., Joosten, I., Andersson, L., Arriens, M.A.,
Bernoco, D., Bissumbhar, B., Byrns, G., van Eijk, M.J.,
Kristensen, B. and Lewin, H.A. (1994) Polymorphism of
Bovine MHC Class II genes. Joint Report of the Fifth
International Bovine Lymphocyte Antigen (BoLA)
Workshop. European Journal of Immunogenetics, 21(4),
259-289.
[3] Van Eijk, M.J., Stewart-Haynes, J.A. and Lewin, H.A.
(1992) Extensive polymorphism of the BoLA-DRB3
gene distinguished by PCR-RFLP. Animal Genetics, 23,
483-496.
[4] Do Nascimento, C.S., Machado, M.A., Martinez, M.L.,
da Silva, M.G.B., Martins Guimares, M.F., Campos, A.L.,
Sousa Azevedo, A.L., Teodoro, R.L., Silva erneque, R.
and Facioni Guimare, S.E. (2006) Association of the
Bovine major histocompatibility complex (BoLA)
BoLA-DRB3 gene with fat and protein production and
somatic cell score in Brazilian gyr dairy cattle (Bos
indicus). Genetics & Molecular Biology, 29(4), 641-647.
[5] Nassiry, M.R., Shahroodi, F.E., Mosafer, J.A.,
Mohammadi, A., Manshad, E., Ghazanfari, S.,
Mohammad Abadi, M.R. and Sulimova, G.E. (2005)
Analysis and frequency of Bovine lymphocyte antigen
(BoLA-DRB3) alleles in Iranian Holstein cattle. Russian
Journal of Genetics, 41(6), 664-668.
[6] Rupp, R., Hernandez, A., Mallard, B.A. (2007) Associ-
ation of Bovine leukocyte antigen (BoLA) DRB3.2 with
immune response, mastitis, and production and type
traitsin Canadian Holsteins. Journal of Dairy Science, 90,
1029-1038.
[7] Duangjinda, M., Buayai, D., Pattarajinda, V., Phasuk, Y.,
Katawatin, S., Vongpralub T. and Chaiyotvittayakul, A.
(2009) Detection of Bovine leukocyte antigen DRB3
alleles as Candidate markers for clinical mastitis re-
sistance in Holstein x zebu. Journal of Animal Science,
87, 469-476.
[8] Jeseph, S. and Rusell, D.W. (2001) Molecular cloning: A
laboratory manual. Cold Spring Habor Laboratory Press.
[9] Miretti, M.M., Ferro, J.A., Lara, M.A. and Conte, E.P.B.
(2001) Restriction fragment length polymorphism (RFLP)
in Exon 2 of the BoLA-DRB3 gene in South American
cattle. Bio Gene, 39(9-10), 311-324.
[10] Maillard, J., Renard, C., Chardon, P., Chantal, I. and
Bensaid, A. (1999) Characterization of 18 new
BoLA-DRB3 alleles. Animal Genetics, 30, 200-203.
[11] Sharif, S., Mallard, B.A., Wilkie, B.N., Sargeant, J.M.,
Scott, H.M., Dekkers, J.C.M. and Leslie, K.E. (1998)
Association of the Bovine major histocompatibility
complex DRB3 (BoLA-DRB3) alleles with occurrence
of disease and milk somatic cell score in Canadian dairy
cattle. Animal Genetics, 29, 185-193.