American Journal of Plant Sciences
Vol.3 No.12(2012), Article ID:25801,7 pages DOI:10.4236/ajps.2012.312214
Leaf Anatomy of Prosthechea moojenii (Pabst) W. E. Higgins and P. silvana Cath. & V. P. Castro (Orchidaceae) as an Approach to Taxonomy of Species Occurring in Biomes Cerrado and Atlantic Forest of the Bahia
![]()
1Instituto de Biologia, Departamento de Botânica, Laboratório de Anatomia Vegetal e Identificação de Madeiras, Campus Universitário de Ondina, Universidade Federal da Bahia, Salvador, Brasil; 2Instituto de Pesquisa Jardim Botânico do Rio de Janeiro, Unidade de Botânica Sistemática, Rio de Janeiro, Brasil.
Email: kellyrbleite@gmail.com
Received September 30th, 2012; revised November 11th, 2012; accepted November 24th, 2012
Keywords: Laeliinae; Flora of Bahia; Velamen; Endemism
ABSTRACT
Prosthechea is a neotropical genus, belonging to the family Orchidaceae. The Brazil recorded 32 species of the genus of which nine occur in the state of Bahia, including P. silvana and P. moojenii. The first, P. silvana is epiphytic, endemic Atlantic Forest areas of South and Southeast of the Bahia and morphologically similar to P. moojenii, rupicolous species considered endemic to the Chapada Diamantina. There are doubts whether these two species are really distinct, because they differ only as to the way of life and aspects related to the size of the inflorescence and floral structure. Thus the anatomical study of these two species, emerges as a tool that aims to investigate the delimitation of these taxa. For this study, the sheet was cut freehand with a razor blade to obtain cross sections and treated by chemical means to obtain paradermic sections, all the material was stained with safranine/Astra blue (safrablau) and safranin, respectively, and the sections were then mounted between a semipermanent and cover slipped slides. The results allowed to distinguish P. silvana of the P. moojenii through the following characteristics observed in the leaf (thickening of the cell walls of epidermal cells, amount and disposition of sclerenchyma fibers and fiber arrangement of vascular and vascular bundles in the mesophyll) and root (number of arcs of xylem and phloem, thickening of cells median cortex), infers taxonomic characters for the genus. The data indicated anatomical distinction between the two species studied here of the Prosthechea, however, suggest the anatomical analysis of other organs to reproductive and vegetative.
1. Introduction
Orchidaceae has a cosmopolitan distribution. It is one of the largest families of angiosperms in the number of existing species. There are at least 24,000 species with specialized characteristics in terms of evolution, mainly in attracting insects [1,2]. According to [3] Brazil is one of the countries that have a large diversity of orchids, about 2500 species distributed in 200 genera [1].
Due to this large diversity of species and little knowledge about some groups occurring in the country, certain genera and species have problems of specific identification such as, for instance, the Prosthechea, a neo-tropical genus, comprising about 69 species distributed from South Florida, Mexico to the south of Brazil. In Brazil there are 32 referred species [4] of which nine are registered for the State of Bahia, among them Prosthechea silvana Cath. & V. P. Castro and Prosthechea moojenii (Pabst) W. E. Higgins. According to [5] the species of the genus are epiphytic or rupicolous, many adapted to a humid habitat such as forests of up to 2600 meters of altitude.
The previous taxonomic treatments of the genus Prosthechea, are a complex sequence of invalid and misapplied classifications involving seven genera Epidendrum L., Encyclia Hook., Dichaea Lindl., Prosthechea Knowles & Westc., Epithecia Knowles & Westc., Hormidium Lindl ex Heynhold and Anacheilium Hoffmanns, becoming a classic case of confused nomenclature which resulted in a problematic taxonomic classification of the genus [6].
The species P. moojenii (Figure 1(a)) and P. silvana (Figure 1(b)) are quite similar in the field mainly in the vegetative stage. The P. silvana is an epiphytic species endemic to the Mata Atlantica ( Brazilian Atlantic Forest) of the south and southeast of Bahia, quite similar to the P.moojenii, a rupicolous species considered endemic to the Chapada Diamantina region.
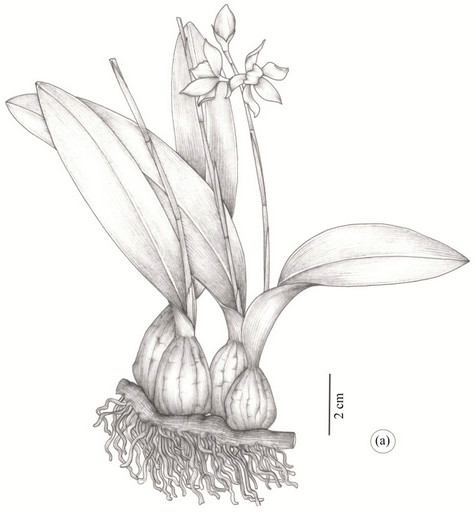
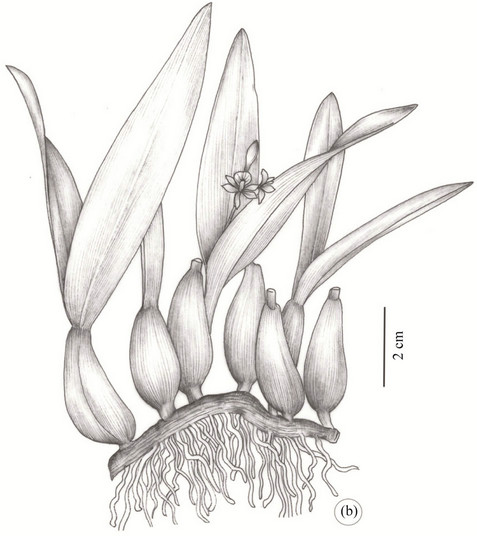
Figure 1. Habit of the Prosthechea (Orchidaceae) species. (a) P. moojenii (Pabst) W. E. Higgins; (b). P. silvana Cath. & V. P. Castro. Scale bars presented in the figures.
The P. moojenii is quite variable in the size and morphology of its vegetative and reproductive organs. It is of a rupicolous species with evident rhizome, bi-leaved ovoid pseudo-bulbs with rigid leaves. It is found in open and sunny places with altitudes that vary between 900 and 1.500 m and encountered so far in the municipalities of Abaíra, Ibicoara, Morro do Chapéu, Palmeiras, Mucugê and Lençóis [3].
The P. silvana grows in humid woods of the south and south coastal area of Bahia (Reconcavo) and presents according to [3] pseudo-bulbs that are narrowly ovoid and bileaved with oblong and slightly curved leaves. It is a medium size plant with approximately 20 cm of height. In this plant sepals and petals have a greenish-yellow color, the labellum is concave, white, with purple striae and growing as an epiphyte.
The main adaptations observed in the vegetative morphology of the orchids evolved to counter adverse environmental conditions, mainly the problems of the conservation of the daily water and seasonality [3] and are observed primarily in the aerial roots of some epiphytes tropical species being in evidence a multi-serial epidermis (multiple skin) consisting of dead cells with thick walls and arranged in a compact manner (velamen) that gives a mechanical protection to the cortex and reduces the loss of water [7,8].
In groups of orchids, anatomical data tend to support the taxonomic treatments in different levels of hierarchy [9]. This answers well the question of delimitation, but, for the genus, the structural analyses are scarce and when carried out focus more on the Laellinae tribe, involving thus few species of the genus. This justifies the importance of this paper which has as a goal to recognize and analyze the anatomic structure of the leaves and the root as parameters to characterize the species of selected Orchidaceae in such a way that the data obtained can verify if there exists an anatomic distinction between the P. moojenii (Pabst). W. E. Higgins and the P. silvana Cath & V. P. Castro, thus expanding the taxonomic delimitation of the referred species and generating information that can contribute to the knowledge of the genus.
2. Material and Methods
The leaves used for the anatomic analysis were fixed in FAA 70% and later preserved in ethyl alcohol of 70% [10]. The specimens that were studied were incorporated to the herbarium collection of the Feira de Santana State University-HUEFS and/or Herbarium Alexandre Leal Costa—ALCB of the Federal University of Bahia, with the following numbers of collectors: P. silvana (Neves M.L.C. et al. 79, Neves M.L.C. et al. 95) collected growing on trees in areas of the Forest Atlantic in the Serra da Jiboia in the municipality of Elisio Medrado and the P. moojenii (Oliveira, R. P. et al. s/n ALCB 97634; SouzaSilva, R. F. 34; Cruz, D. T. 4; Ganev s/n HUEFS 15851; Seixas, F. C. 1), collected among rocks in the grassland areas of the municipality of Morro do Chapéu.
Apical, medium and basal regions of adult roots and leaves of at least three different individuals of each species, always when possible, were sectioned. The paradermic and cross sections were done free hand, with the help of a razor blade, stained with 1% aqueous safranine and astro blue and safranine (modified by Bukatsch, 1972 apud [11]) respectively. Furthermore, to obtain the epidermis, a method of epidermic dissociation was used with hydrogen peroxide of 30% and glacial acetic acid [11] which was then stained with safranine 1%. The laminas were mounted in a glycerin base at 50%. For the roots, the samples were sectioned at 0.5 cm from the base of the rhizome and the radicle apex on. The histochemical test to identify the flavonoids through the solution of potassium hydroxide [11] was done only with fresh P. moojenii samples. This justifies the not testing of P. silvana since the available specimens were only available from the herbarium material.
All the sections were analyzed, described and photographed under the photo microscope Olympus BX41 with a coupled camera of the type Qimaging Go-3 81661688.
3. Results and Discussion
Data referring to the leaf anatomy in the paradermic section of the P. moojenii and P. silvana can be observed in Figure 2 and in cross sections of the leaf and root in Figures 3 and 4.
3.1. Leaf
The described species present in paradermic vision and on both sides, cells of irregular shape varying from quadrangular to polygonal with cell walls of straight contour (Figures 2(a)-(d)) and thicker in the P. silvana (Figures 2(c) and (d)) than in the P. moogenii (Figures 2(a) and (b)). Anomocytic and tetracytic types of stomata were present only in the abaxial side of the epidermis and presented cuticular reinforcement in the region of the ostiole, more strongly thickened in the P. silvana (Figures 2(f) and 3(g)). The P. moojenii showed stomata only of the tetracytic type (Figure 2(e)) while the P. silvana presented both types (Figure 2(f)). The aspects related to the shape of the epidermis and type and stomatic reinforcement had already been observed by [9] with Laeliinae for some species of the genus but not for the species that were studied here.
In cross section the presence of thickened cuticle was observed. It was more evident in the P. silvana (Figure

Figure 2. Photomicrographs in optical microscopy of the epidermis of leaf blades of the Prosthechea species. (a)-(b), E. P. moojenii (Pabst) W. E. Higgins. (a) Adaxial surface; (b) Abaxial surface; (e) Detail of the paracitic stomata; (c), (d), (f) P. silvana Cath. & V. P. Castro. C. Adaxial surface; (d) Abaxial surface; (f) Detail of the anomocitic stomata. Scale bars = 100 µm.
3(f)) than in the P. moojenii (Figure 3(e)). In both species the epidermis is uniseriate formed by cells of oval to rectangular shape (Figures 3(e) and (f)) and the stomata are at the same level of the other epidermal cells. It is possible to observe in the guard cells a greater thickening of the anticlinal wall which forms the ostiole (Figure 3(g)).
In the main nervure of the two species that were studied, one observes two to three layers of parenchymal cells rather vertically elongated above the vascular cluster which corresponds to the hypodermis being more evident since it has more cellular layers in the P. silvana (Figure 3(b)-arrow). The vascular cluster in this region is shifted to the abaxial side and surrounded partially by a cap of pericyclic fibers in the P. silvana (Figure 3(b)) while the P. moojenii shows fibers only in the phloem region. The mesophyll is homogeneous with few inter-
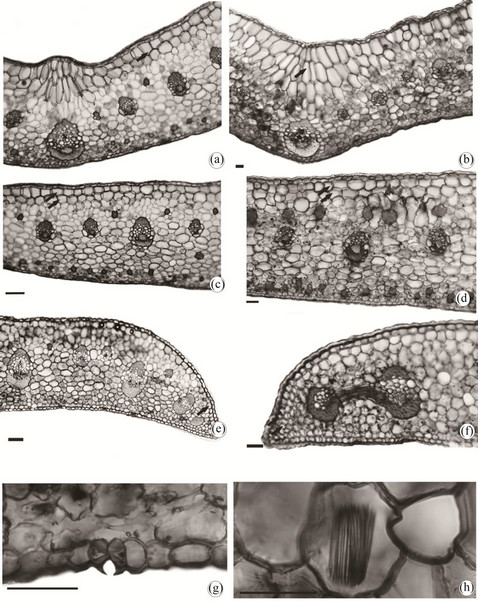
Figure 3. Photomicrographs in optical microscopy of the leaf blades of the Prosthechea species in cross sections. (a), (c), (e), (g) P. moojenii (Pabst) W. E. Higgins and (b), (d), (f), (h) P. silvana Cath. & V. P. Castro; (a)-(b) Midvein; (c)-(d) Mesophyll; (e)-(f) Leaf margin; (g) Detail of wall thickening anticline of the guard cells of the stomata; (h) Detail of raphides in hypodermic cells. Scale bars = 100 µm (arrow = Hypodermis).
cellular spaces and the cells vary from round to elliptical (Figures 3(c) and (d)). The number of cell layers in this region is less in the P. moojenii than in the P. silvana (Figures 3(c) and (d)) respectively. In the region of the mesophyll you can also observe a parenchymatous hypodermis toward only to the abaxial side and adjacent to the epidermis (Figures 3(c) and (d)-arrow) with two intermittent cell layers up to the region of the leaf margin in P. silvana (Figure 3(f)) and only one layer in P. moojenii (Figure 3(e)).
[12] studying anatomical aspects of Laeliinae observed that the Prosthechea bootheiana (Lindl.) W. E. Higgins and P. radiata (Lindl.) W. E. Higgins do not present a hypodermis; thus, differing from the standard shown in the species studied and in both species. One believes that its presence is related to the accumulation of water as already observed by [13] in the Arecaceae species due to an environment in which the plants were collected and the habit they present.
Collateral vascular clusters of different sizes occur throughout the mesophyll in the central region in both species and alter their positions (Figures 3(c) and (d)) reaching the edge (Figures 3(e) and (f)). It is possible to observe that the vascular clusters show a larger amount of fibers forming caps next to the poles of the xylem and phloem in the P. moojenii (Figures 3(a), (c) and (e)) than in the P. silvana (Figures 3(b) and (d)) and in this latter species the fibers next to the vascular clutters are more concentrated in the region of the edge (Figure 3(f)). Tissue of support in the form of a cluster of sclerenchymatous fibers was observed in both species in layers adjacent to the epidermal sides, P. moojenii presenting a smaller amount and a more dispersed organization of this tissue compared to the P. silvana (Figure 3(d)).
From the analysis of the leaf edge, P. moojenii presented a gradual decrease from two to one layer of hypodermic cells, clusters of pericyclic fibers in the poles of the vascular tissue and presence of clusters of scleraenchymatous fibers (Figure 3(e)) and P. silvana (Figure 3(f)) showed a continuity of both layers of hypodermic cells, vascular clusters almost totally surrounded by a large number of pericyclic cells and absence of clusters of sclerenchymatous fibers.
Crystals in the form of raphides were found more frequently in the region of the mesophyll and associated to the cells of the hypodermis in P. silvana (Figure 3(h)).
3.2. Root
It was observed that the two species even though growing in different substrates have plenty of adventitious roots which favor their attachment, be it epiphytic as well as rupicolous. Cross sections of the roots of both species (Figures 4(a) and (k)) showed a pluri-stratified epidermis (Figures 4(a) and (b)) that is the aerofoil that originates from the periclinal divisions of initial cells of the protoderm [14].
In this study the velamen presented five to seven cell layers and is differentiated in the epivelame (first layer of cells) composed of this walled cells and of quadrangular to pentagonal shape and the endovelamen (other layers) with cells of hexa-to heptagonal shape (Figures 4(c) and (d)). All the cells of the velamen had a fine reticulate to anastomotic thickening (Figure 4(e)). These characteristics were also observed by [12] in the P. boothiana.
The presence of a velame is typically observed in Orchidaceae as well as in other groups of monocotyledonae such as Velloziaceae [15] and Bromeliaceae [16,17] and referred by [9] for other species of the Prosthechea genus and by [18] that studied the roots of various species of different genera of Orchidaceae, as an important characteristic for the water retention which permits its reservation and temporary mobility and for the mechanical protection.
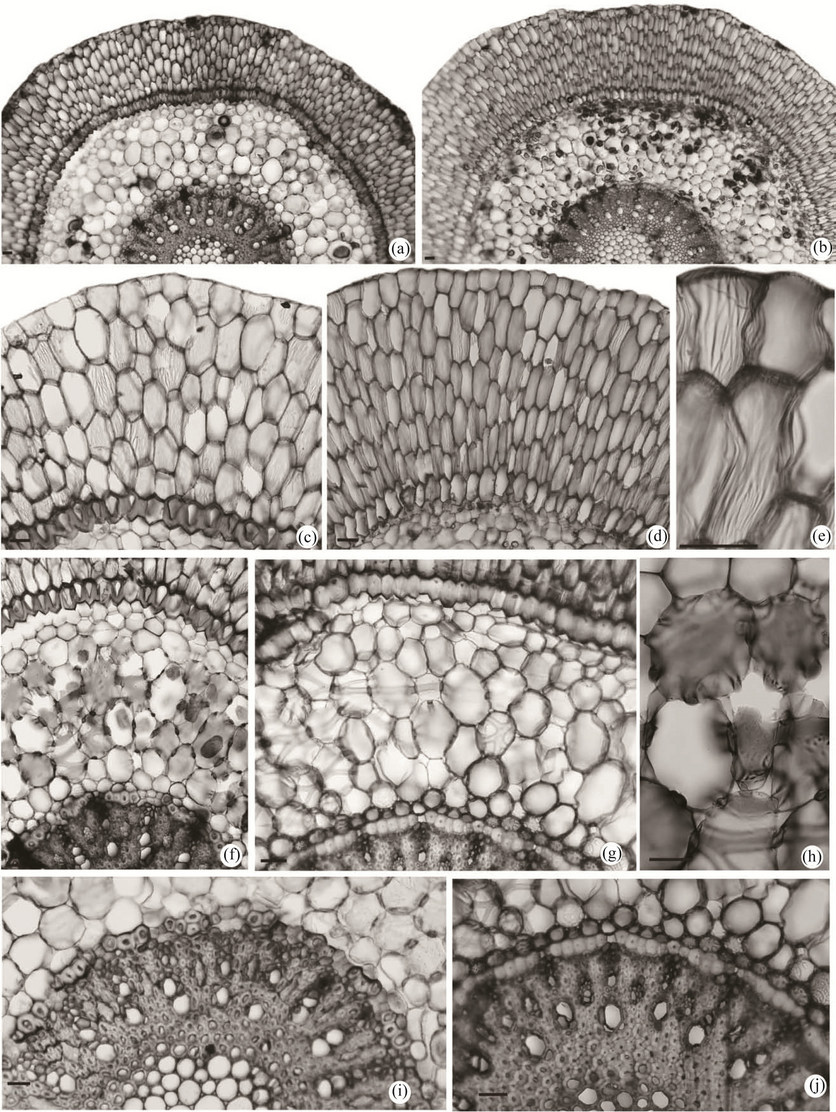
Figure 4. Photomicrographs in optical microscopy of root of the Prosthechea species in cross section. (a), (c), (e), (f)-(g), (i) P. moojenii (Pabst) W. E. Higgins and (b), (d), (h), (j) P. silvana Cath. & V. P. Castro; (a) General aspects of adult root; (b) General aspects of root young; (c)-(d) Detail of velamen and exodermis; (e) Detail of cells of the velamen with reticulate thickening; (f)-(g) Cortex showing cells of the exodermis and endodermis; (h) Detail of cortex showing cells of with wide lignified phi thickenings; (i)-(j) Detail of vascular cylinder. Escala = 100 µm.
The cortical parenchyma also showed differentiation. It was possible to observe three distinct layers in the two species of Prosthechea that were studied; outer, middle and inner cortex (Figures 4(f) and (g)). The outer cortex is the proper exodermis which in the P. moojenii has highly lignified cells with an uneven thickening in U shape, the external periclinal wall being less thick than the inner one and the two anticlines (Figures 4(c) and (f)) according it was written for P. faresiana (Bicalho) W. E. Higgins and P. alagoensis (Pabst) W. E. Higgins by [9] and with homogeneous thickening in O in P. silvana (Figures 4(d) and (g)). The middle cortex has few intercellular spaces with cells of elliptical to rounded shape of unequal size in the two species and show a thickening strongly lignified and of a more un-shaped type in the P. moojenii (Figure 4(h)) and in the shape of a strip in P. silvana (Figure 4(g)). The endodermis shows the cellular walls highly thickened in the periclinal and anticlinal walls in the P. moojenii (Figure 4(i) as well as in the P. silvana (Figure 4(j)) with the presence of passing cells only with primary wall opposite to protoxylem poles. In the two species the endodermis presents a set of thickened cells being more regular in the P. silvana (Figure 4(j)) than in the P. moojenii (Figure 4(i)).
The vascular cylinder had 14 - 17 poles of protoxylem interspaced with phloem in the P. moojenii reaching up to 24 poles of protoxylem in the P. silvana. In the roots of both species one could observe in the middle and base regions, strongly lignified medullar parenchyma.
[16] discussed in their work that the reduction of vascular tissues in epiphytic species is higher due to the fact that these species have roots mainly for their function of fastening.
Raphides in the root were present in the region next to the inner cortex. This type of crystal was also observed in other species of the genus [9] and, according to these authors, besides the crystals, there exist flavonoids that protect the plant from herbivorous predators and are also associated to the metabolism (free radicals) being frequent in plants that live under some kind of stress [19].
In both species the vascular clusters of larger and smaller size are arranged alternately. This suggests a possible taxonomic feature for the genus. However, from the features that were observed, the P. silvana differs from the P. moojenii presenting in the leaf: a thicker cuticle, epidermal cells with thicker walls, anomocytic besides tetracytic stomata, ostiole with reinforcement in the anticline walls, main nervure with a higher number of hypodermic cells that extend with two layers up to the leaf edge, mesophyll with a higher number of sclerenchyma but gradual reduction up to the leaf edge, being completely absent in it and, in the root: medium cortex formed by the thickening strip and vascular cylinder formed by the presence of a higher amount of protoxylem poles.
The characteristics described above point at P. silvana as a species that has a higher number of xeromorphic features compared to the P. moojenii, even the first having been collected in an environment with a higher availability of water and humidity such as the environment of the Mata Atlantica. From the analysis of the characteristics that were observed it is believed that the P. silvana should show more mesamorphic traits as it grows in a humid area of the Mata Atlantica which suggests that these characteristics are remarkable for the species differing from the P. moojenii that inhabit rock crevices in an environment with constant sunshine, not excluding the possibility of water as well as organic matter being available but, in smaller amounts.
Based on the comparison of the anatomic results obtained in the comparative study of leaves and roots of the P. moojenii and the P. silvana, it was verified that the two taxa are indeed distinct and that the anatomy is an important tool for the separation of the species.
Once that the published papers of anatomic analysis in the specific level for the genus are always Prosthechea related to Laellinae, the results obtained here for these two species are highly important. However, it is suggested that anatomic analyses of other vegetative organs and maybe reproductive organs as well, be carried out for a further delineation of the two species that were studied.
4. Acknowledgements
The authors thank FAPESB-Fundação de Amparo à Pesquisa do Estado da Bahia for the financial support for the development of this research (PPP 083/2010 and PNE 0020/2011).
REFERENCES
- V. C. Souza and H. Lorenzi, “Botânica Sistemática: Guia Ilustrado para Identificação das Famílias de Angiospermas da Flora Brasileira, Baseado em APG II,” Plantarum, Nova Odessa, 2005.
- V. C. Oliveira and M. G. Sajo, “Morfoanatomia Caulinar de nove Espécies de Orchidaceae,” Acta Botanica Brasilica, Vol. 15, No. 2, 1999, pp. 177-188. doi:10.1590/S0102-33062001000200004
- A. L. V. Toscano-De-Brito and P. Cribb, “Orquídeas da Chapada Diamantina,” Editora Nova Fronteira, São Paulo, 2005.
- F. De Barros, F. Vinhos, V. T. Rodrigues, F. F. V. A. Barberena, C. N. Fraga and E. M. Pessoa, “Orchidaceae,” Lista de Espécies da Flora do Brasil, Jardim Botânico do Rio de Janeiro, 2012. http://floradobrasil.jbrj.gov.br/2012/FB012106
- W. E. Higgins, “Prosthechea: A Chemical Discontinuity in Laeliinae,” Lankesteriana, Vol. 7, No. 2, 2003, pp. 39- 41.
- W. E. Higgins, “A Reconsideration of the Genus Prosthechea (Orchidaceae),” Phytologia, Vol. 82, No. 5, 1997, pp. 370-383.
- K. Esau, “Anatomia das Plantas com Sementes,” E. Blucher, São Paulo, 1974.
- B. Apezzato-Da-Glória and A. H. Hayashi, “Raiz,” In: B. Apezzato-Da-Glória and S. M. Carmello-Guerreiro, Eds., Anatomia Vegetal, Ed., UFV, Viçosa, 2006, pp. 267-281.
- M. F. O. Pires, J. Semir, G. F. A. M. Pinna and L. P. Felix, “Taxonomic Separation of the Genera Prosthechea and Encyclia (Laeliinae: Orchidaceae) Using Leaf and Root Anatomical Features,” Botanical Journal of Linnean Society, Vol. 143, No. 3, 2003, pp. 293-303. doi:10.1046/j.1095-8339.2003.00195.x
- D. A. Johansen, “Plant Microtechnique,” McGraw Hill Book Co., New York, 1940.
- J. E. Kraus and M. Arduim, “Manual Básico de Métodos em Morfologia Vegetal,” EDUSP, São Paulo, 1997.
- W. L. Stern and B. S. Carlsward, “Comparative Vegetative Anatomy and Systematics of Laeliinae (Orchidaceae),” Botanical Journal of the Linnean Society, Vol. 160, No. 1, 2009, pp. 21-41. doi:10.1111/j.1095-8339.2009.00818.x
- K. R. B. Leite and V. L. Scatena, “Anatomia do Segmento Foliar de Espécies de Syagrus Mart. (Aracaceae) na Chapada Diamantina, Bahia, Brasil,” Sitientibus: Série Ciências Biológicas, Vol. 1, No. 1, 2001, pp. 2-14.
- A. Fahn, “Plant Anatomy,” Pergamon Press, New York, Oxford, 1974.
- N. L. Menezes and A. M. Giulietti, “Campos Rupestres— Paraíso Botânico na Serra do Cipó,” Ciência Hoje, Vol. 5, 1986, pp. 38-44.
- P. B. Pita and N. L. Menezes, “Anatomia da Raiz de Espécies de Dyckia Schult. f. e Encholirium Mart. ex Schult. & Schult. f. (Bromeliaceae, Pitcairnioideae) da Serra do Cipó (Minas Gerais, Brasil), com Especial Referência ao Velame,” Revista Brasileira de Botânica, Vol. 25, No. 1, 2002, pp. 25-34. doi:10.1590/S0100-84042002000100005
- I. V. Silva and V. L. Scatena, “Anatomia de Raízes de Nove Espécies de Bromeliaceae (Poales) da Região Amazônica do Estado de Mato Grosso, Brazil,” Acta Botânica Brasilica, Vol. 25, No. 3, 2011, pp. 618-627. doi:10.1590/S0102-33062011000300015
- I. V. Silva, R. M. S. A. Meira and A. A. Azevedo, “Anatomia de Raízes de Espécies de Orchidaceae do Parque Estadual da Serra do Brigadeiro, Minas Gerais,” Hoehnea, Vo. 37, No. 1, 2010, pp. 147-161.
- H. Yamasaki, Y. Sakihama and N. Ikehara, “FlavonoidPeroxidase Reaction as Detoxification Mechanism of Plant Cell against H2O2,” Plant Physiology, Vol. 115, No. 4, 1997, pp. 1405-1412.