Advances in Bioscience and Biotechnology
Vol.5 No.1(2014), Article ID:42359,9 pages DOI:10.4236/abb.2014.51010
Inoculation with arbuscular mycorrhizal fungi improves seedlings growth of two sahelian date palm cultivars (Phoenix dactylifera L., cv. Nakhla hamra and cv. Tijib) under salinity stresses
1Laboratoire Campus de Biotechnologies Végétales, Département de Biologie Végétale, Faculté des Sciences et Techniques, Université Cheikh Anta Diop, Dakar, Sénégal
2Laboratoire Commun de Microbiologie IRD/ISRA/UCAD, Route des Hydrocarbures, Dakar, Sénégal
3Laboratoire d’Ecologie et d’Ecohydrologie, Département de Biologie Végétale, Faculté des Sciences et Techniques, Université Cheikh Anta Diop, Dakar, Sénégal
4IRD Institut de Recherche pour le Développement, Montpellier, France
Email: *leopoldiatta@gmail.com
Copyright © 2014 Ibrahima Léopold Djitiningo Diatta et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. In accordance of the Creative Commons Attribution License all Copyrights © 2014 are reserved for SCIRP and the owner of the intellectual property Ibrahima Léopold Djitiningo Diatta et al. All Copyright © 2014 are guarded by law and by SCIRP as a guardian.
Received 11 October 2013; revised 23 December 2013; accepted 12 January 2014
KEYWORDS
Sahel; Date Palm; Salinity; Inoculation; Mycorrhizae; Glomus; Proline
ABSTRACT
This study presents an analysis of the impact of mycorrhizal inoculation on growth under salt stress of date palms cultivars Nakhla hamra (NHH) and Tijib known in the Sahel for their earliness in flowering and fruiting. The seedlings were grown in a greenhouse on a sandy substrate watered to field capacity every two days and were subjected to increasing levels of NaCl (0, 1, 2, 4, 6, 8 and 16 g∙L−1) and then inoculated with 5 strains of Glomus (G. aggregatum, G. intraradices, G. verriculosum, G. mosseae, G. fasciculatum). The experimental design was a randomized complete block with three factors (cultivars: Tijib and Nakhla Hamra x seven levels of NaCl concentrations x six levels of inoculum). The results showed that in the absence of NaCl, G. verriculosum significantly enhanced shoot growth: 33.5 cm against 30.3 cm in the control and roots growth: 81.5 cm against 78 cm in NHH, while in Tijib, the growth is stimulated by Glomus fasciculatum: 33.8 cm against 32.7 cm for stems and 90 cm against 86 cm for the roots of inoculated plants and controls. In contrast, in the presence of NaCl 8 g∙L−1, NHH has a better growth in the presence of G. intraradices: 37.3 cm against 30.6 cm for stems and 77 cm against 73 for roots, while Tijib grows better in the presence of G. fasciculatum with respectively 31.9 cm against 31.7 cm and 51.27 cm against 51.6 cm for stems and roots of inoculated plants and controls. Biochemical analysis revealed that changes in levels of proline depend on the cultivar, the mycorrhizal strain used and concentrations of NaCl applied. These results open the prospect of using mycorrhizal fungi to improve the productivity of palm trees in the Sahel.
1. INTRODUCTION
The date palm (Phoenix dactylifera L.) is grown in the Asian and Africa hot arid and semi-arid zones, but also in Australia, in some Latin American countries where it was introduced in the 18th century, and in Mediterranean regions of Europe. This species is an excellent fruit tree of the desert where he plays an economic role through the production of dates, ecological role since it constitutes the backbone of oasis and social role, because it stabilizes rural populations [1]. However, several constraints limit the expansion of its culture. In North Africa, partic ularly in Morocco and Algeria, it is mainly Bayoud disease, Fusarium wilt caused by the fungus Fusarium oxysporum f sp. Albedinis that, for several decades, decimated palms [2]. In sub-Saharan Africa, it is mainly drought, salinization and aging hindering date palm culture and causing the disappearance of many cultivars thereby and the depletion of the gene pool [3]. In the Sahel, the expansion of cultivation is hampered by the lack of availability of cultivars adapted to local soil and climatic conditions. In addition, the overlaps between the period of fruit ripening and the rainy season or soil salinity are major obstacles to the development of date palm. In Senegal, land salinization particularly affects coastal areas where it leads to the formation of extensive salt flats where it remains only a few species of plants more or less tolerant [4]. The environmental degradation due to salinization caused a mobilization of different actors (state, the scientific community, economic operators) involved in the field of agriculture. Their actions aim not only to curb environmental degradation, but also to fight against the exodus of rural populations to urban centers by safeguarding the oasis ecosystem [5].
The development of date palm in the Sahelian area therefore requires the selection and dissemination of early and salinity tolerant cultivars [6,7].
Despite the full range of strategies to fight against salinity (leaching, drainage, flood and use of irrigation water quality), no use of soil microbiological potential, particularly arbuscular mycorrhizal (AM) fungi has not been recommended and approved to date for increasing the productivity of palm. The work of [8] and those of [9] showed that fungi can grow naturally in various saline environments where they can improve plant growth. [10] determined that the AM fungi generally observed in saline soils belong to the order of Glomerales particularly Glomus.
In the present work we studied the influence of inoculation with 5 strains of mycorrhizal fungi of the genus Glomus (G. aggregatum, G. intraradices, G. verriculosum, G. mosseae and G. fasciculatum) on plants cultivars Nahla hamra (NHH and Tijib subjected to salt stress conditions). The impacts of inoculation on the growth of root and above ground part and the evolution of physiological and biochemical parameters associated with plant development were particularly studied to determine best symbiotic partners (cultivars of date palms/mycorrhizal strains) by simulating salinity levels observed in the Sahelian environment.
2. MATERIAL AND METHODS
2.1. Plant Material
The study was conducted on seeds harvested on females trees of two varieties of date palm Nahla hamra (NHH) and Tijib, characterized by very early flowering and fruiting, selected directly in the palm of Kseir Torchane town located at 20 km in the North of the Atar in Mauritania (latitude 20˚31ʹ North 013˚03ʹ West).
2.2. Fungal Material
To assess the impact of inoculation on growth of date palm seedlings, we used five strains of mycorrhizal fungi of the genus Glomus of the collection of the “Laboratoire Commun de Microbiologie (LCM, IRD/ISRA/UCAD)’’ of the research center ISRA-IRD of Dakar Bel-Air in Sénégal. The origin and references of these strains are given in Table 1.
2.3. Conditions of in Vitro Germination of Seeds
The seeds were sterilized with H2SO4 96% for 10 min and then rinsed with sterile distilled water. They were then soaked in sterile water for 24 h before being germinated in tubes (25 × 150 mm) containing 20 ml of water agar (8 g∙L−1 of agar) and placed in lighting module (80 μE∙s−1∙m−2) with a 12 h/12 h photoperiod at a constant temperature of 27˚C ± 0.2˚C.
2.4. Greenhouse Culture Conditions and Experimental Design
After three months of in vitro culture, the plants thus obtained were transferred to the greenhouse in plastic pots 12 cm high and 16 cm in diameter. These pots were filled to 4/5 of the volume of soil from Sangalkam. The soil of Sangalkam is sandy (94.9% sand content, clay 3.6%, silt 0.8%), poor in minerals, especially nitrogen (0.02%) and phosphorus (4.8 mg∙kg) [12]. The soil was watered to field capacity 48 hours before the transfer of plants.
To assess the threshold of salt stress in the greenhouse, a first series of experiments was set up by using a ran-

Table 1. Origin and references of strains of arbuscular mycorrhizal (AM) fungi of genus Glomus.
domized complete block design with two factors: cultivars (2) x NaCl levels (7). NaCl was previously tested at 0, 1, 2, 4, 6, 8 and 16 mg∙L−1 and a total of 20 plants per cultivar and per salinity condition were used. The mycorrhizal inoculation of seedlings was performed 4 months after their mobilization in greenhouse. Twenty gram (20 g) of fungal inoculum of each of the five Glomus strains tested was placed at 5 cm in depth around the collar of the plant. Four months after inoculation, three levels of salinity (0, 8 and 16 mg∙L−1 NaCl) previously defined following the first experiment were applied. These salt concentrations corresponded respectively for the plants of studied cultivars to the levels without stress, moderately stressed and highly stressed. A randomized block with three factors: Cultivar (2) x NaCl levels (3) x inoculation conditions (5 mycorrhizal strains + control non-inoculated) has been set up for this second experiment. For each condition of salinity, a total of 20 plants/ cultivar/condition of inoculation was used, that equals to 360 plants per cultivar. The plants were watered to field capacity every two days and maintained under these conditions for 2 months.
2.5. Evaluation Methods of Mycorhization
The observation of root colonization was made after staining by the method of [13] and the percentage of colonization was determined according to the “Gridline Intersect” method of [14]. Mycorrhizal parameters were evaluated according to the method of [15] (Figure 1). The intensity of mycorrhizal (I) corresponded to the observed proportion of root fragments colonized by AM fungi. Each fragment was then analyzed using a ranked intensity mycorrhizal rated from zero (0) to five (5).
2.6. Dosage of Proline
The tolerance of the cultivars NHH and Tijib to NaCl was studied by evaluating the proline rates accumulated by plants. The protocol described by [16] was used to
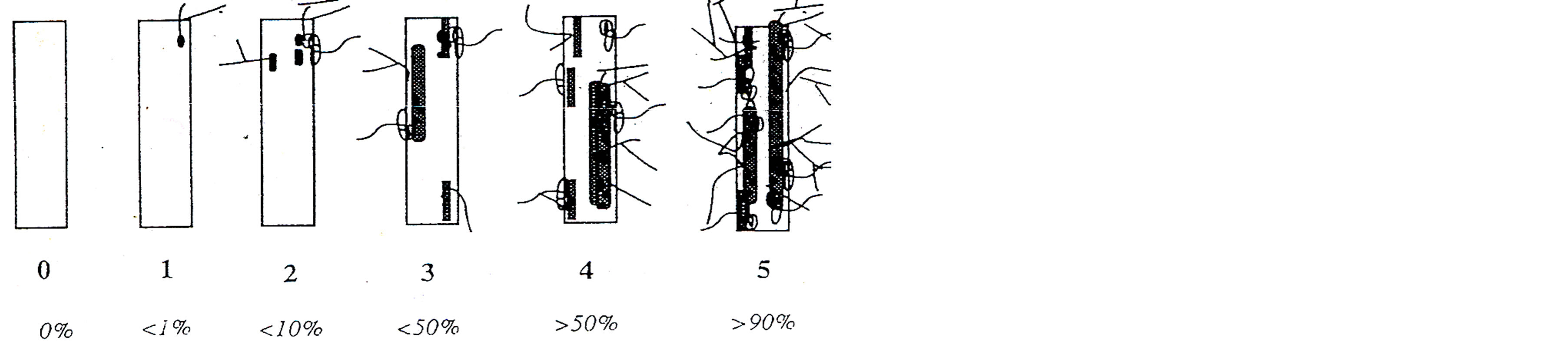
Figure 1. Rating of mycorrhizal infection (Class 0 to Class 5) [15]. I (%) = (95n5 + 70n4 + 5n2 + 30n3 + n1)/total number of fragments observed; n5 = number of recorded five fragments; n4 = number of fragments scored 4, n3 = number of recorded fragments 3, n2 = number of recorded fragments 2, n1 = number of fragments denoted 1.
extract and determine proline. The extraction was made from a composite mixture of 100 mg of leaf segments of three plants per condition. The concentration of proline was determined in a spectrophotometer by measuring the optical density (OD) at 520 nm and three replicates per culture condition were performed.
2.7. Statistical Analysis
For each culture condition, biochemical parameters (proline content), mycorrhiza (intensity of mycorrhiza) and shoot growth (elongation of epicotyl) and ground plants (number of secondary roots, elongation of the taproot) were measured. The data analysis was performed using ANOVA/MANOVA (General Linear Model) STATISTICA software, version 6. Students—Newman and Keuls’ test at 5% was used to compare means after analysis of variance.
3. RESULTS
3.1. Effect of Addition of NaCl on Plant Growth in Cultivars NHH and Tijib
The results show that the elongation of the aboveground part is variable depending on the studied cultivars and the tested NaCl concentrations (Figures 1 and 2). In the cultivar NHH, there is a very significant medium effect on stems growth (F = 32.40, P = 0.000). In this cultivar, the application of NaCl at concentrations greater than or equal to 8 g∙L−1 in the culture medium, resulting in a significant decrease in elongation of the epicotyl in length going from 18.9 cm on average in controls to 16.17 cm in presence of 8 g∙L−1 NaCl (Figure 2). In the cultivar Tijib, our results also showed significant differences (F = 3.345, P = 0.009) between the control (21.86 cm) and plants treated with NaCl 8 g∙L−1 in which the average length of epicotyls of 20.21 cm (Figure 3). However, regardless of the cultivar concerned, the NaCl concentration of 16 g∙L−1 has a depressive effect on the elongation of the epicotyl. The results suggest that greenhouse plants Tijib tolerate better the effect of NaCl than NHH. However, it appears from these two cultivars that the threshold of perception of salt stress was observed from the NaCl concentration of 8 g∙L−1 (Photo 1).
3.2. Effect of Salt Stress on the Intensity of Mycorrhiza in Cultivars NHH and Tijib
The ANOVA show that there is a variety effect (F = 48.09, P = 0.000), as well as medium effect (F = 13.45, P = 0.000) and the interaction variety x inoculation conditions (F = 10.65, P = 0.000) is very significant on the intensity of mycorrhizal observed after 4 months of culture. In the cultivar NHH, G. intraradices allows having the highest values at 8 g∙L−1 NaCl (28.81%) of intensity of mycorr-
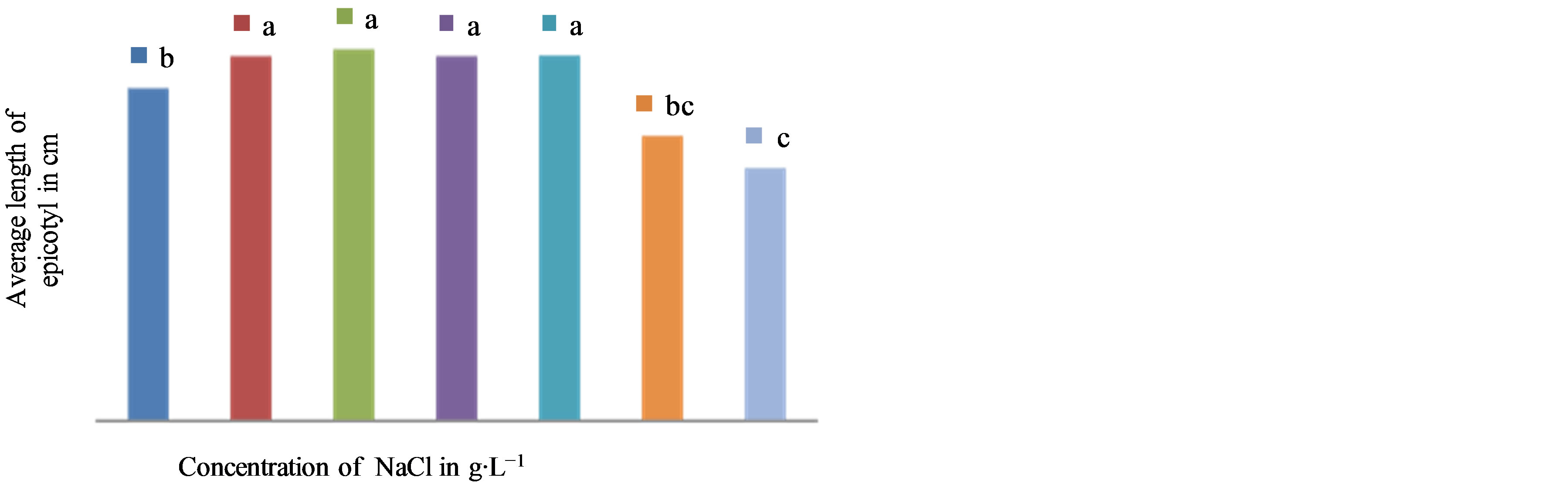
Figure 2. Effect of NaCl concentration on the growth of epicotyls of seedlings in NHH after 6 weeks of culture in greenhouse. Means that do not share a letter are significantly different at 5% using Student-Newman and Keuls’ test.
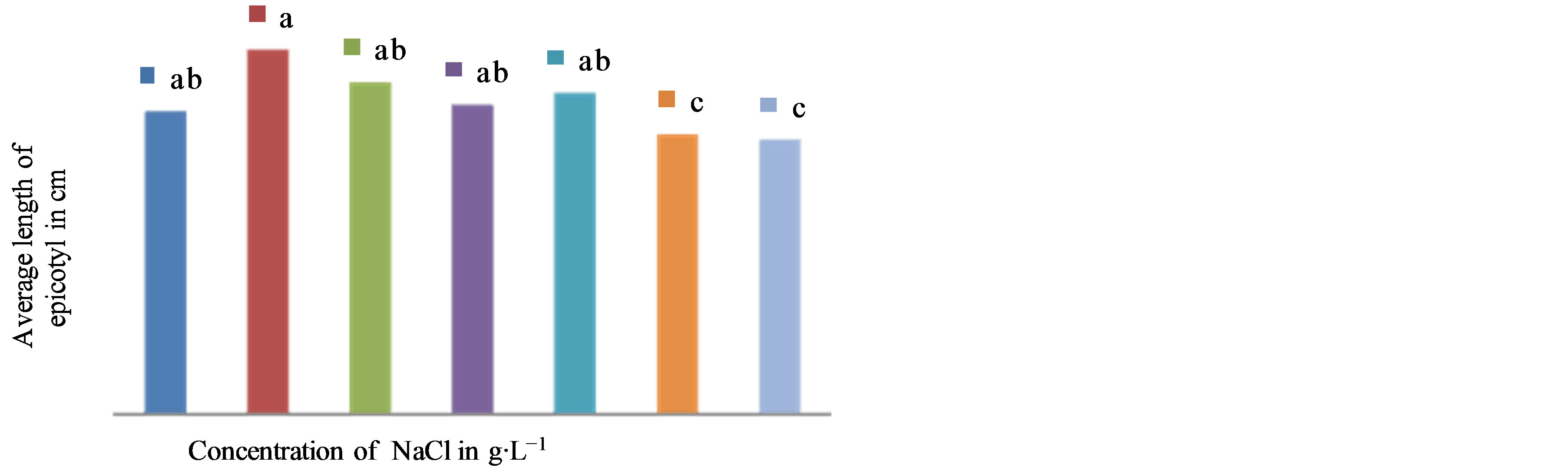
Figure 3. Effect of NaCl concentration on the growth of epicotyls of seedlings in Tijib after 6 weeks of culture in greenhouse. Means that do not share a letter are significantly different at 5% using Student-Newman and Keuls’ test.

Photo 1. Growth and development of date palm seedlings under different salinity. 0T: Control without salt, 8T: NaCl 8 g∙L−1, 16T: NaCl 16 g∙L−1. We can see the good development of the control; salt sensitivity is evident from 8 g∙L−1 and plants become necrotic at 16 g∙L−1 NaCl.
hization compared to all the tested inoculation conditions (Figure 4, Photo 2). In contrast, in the cultivar Tijib, it is rather with G. fasciculatum that the maximum values of the intensity of mycorrhiza are observed (8.10%) in
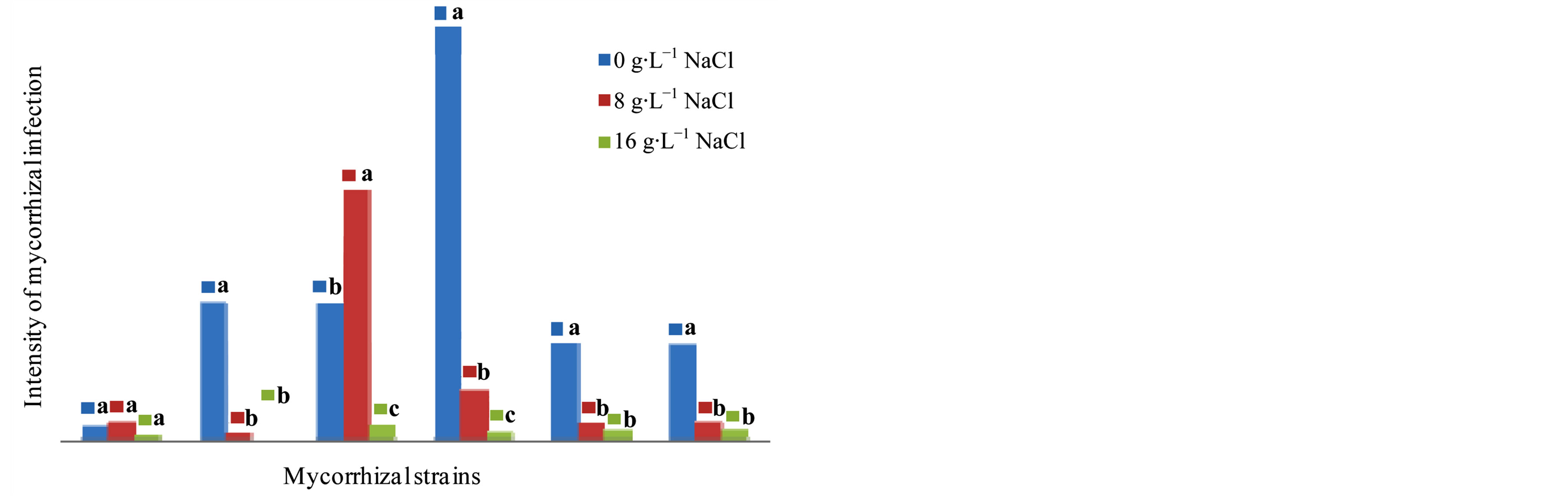
Figure 4. Impact of five strains of Glomus on the intensity of mycorrhizal plants in the cultivar NHH in the presence of 0, 8 and 16 g∙L−1 after 4 months in greenhouse. Means that do not share a letter are significantly different at 5% using Student-Newman and Keuls’ test.
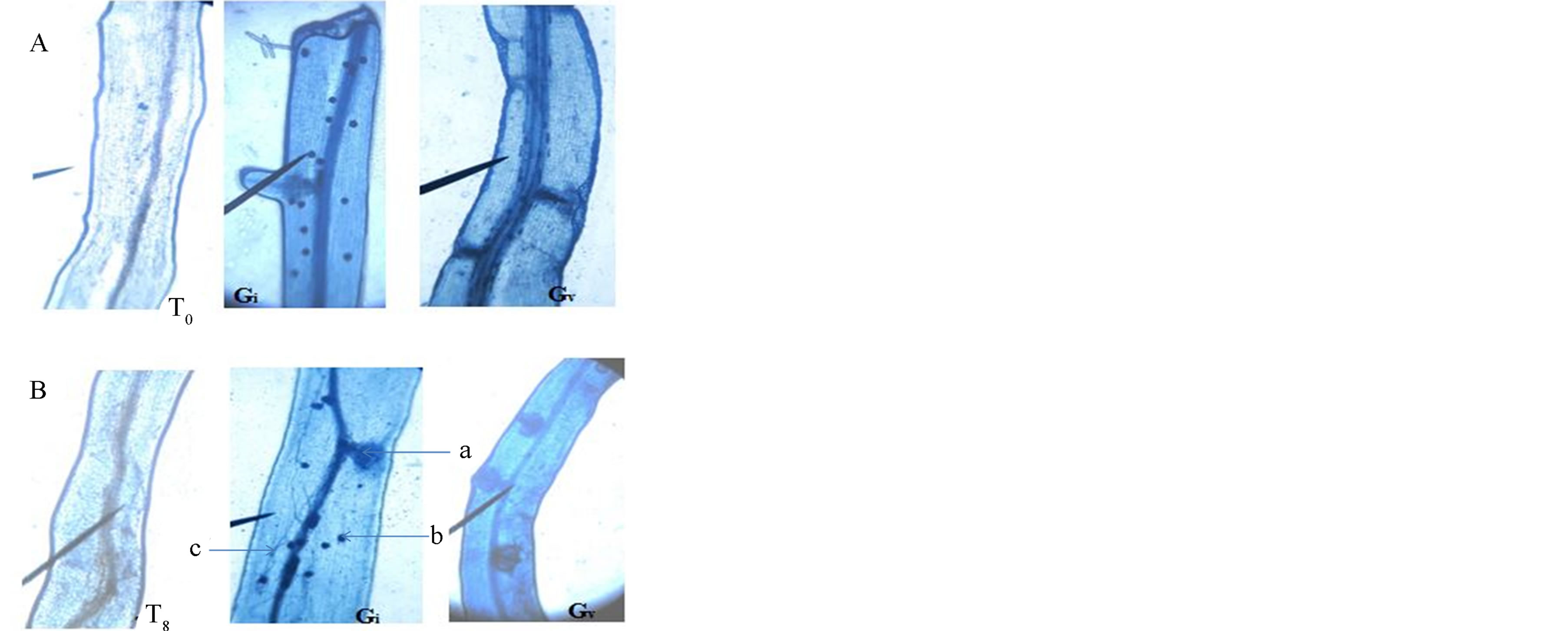
Photo 2. Colonization of plant roots by NHH strains of fungi G. intraradices and G. verriculosum in the absence of stress (A) and stress situation (B). T0 (Witness) Gi (G. intraradices) Gv (G. verriculosum). In absence of stress, we can observe in both strains the presence of spores (b). However, when stress is applied, vesicles (a) and hyphae (c) mycelium are developed.
stressed conditions (Figure 5, Photo 3).
3.3. Effect of Inoculation on the Growth of Stems of Plants in Cultivars NHH and Tijib Grown in the Absence of Salt Stress
The lengthening of the aboveground part of the inoculated plants in the absence of salt varies among cultivars studied and mycorrhizal strains tested (Tables 2 and 3). The analysis of variance showed no significantly interaction variety x mycorrhizal strains in NHH (F = 0.00, P = 0.085) and Tijib (F = 0.00, P = 0.279). However, our results show that the strain Glomus verriculosum stimulates the de-
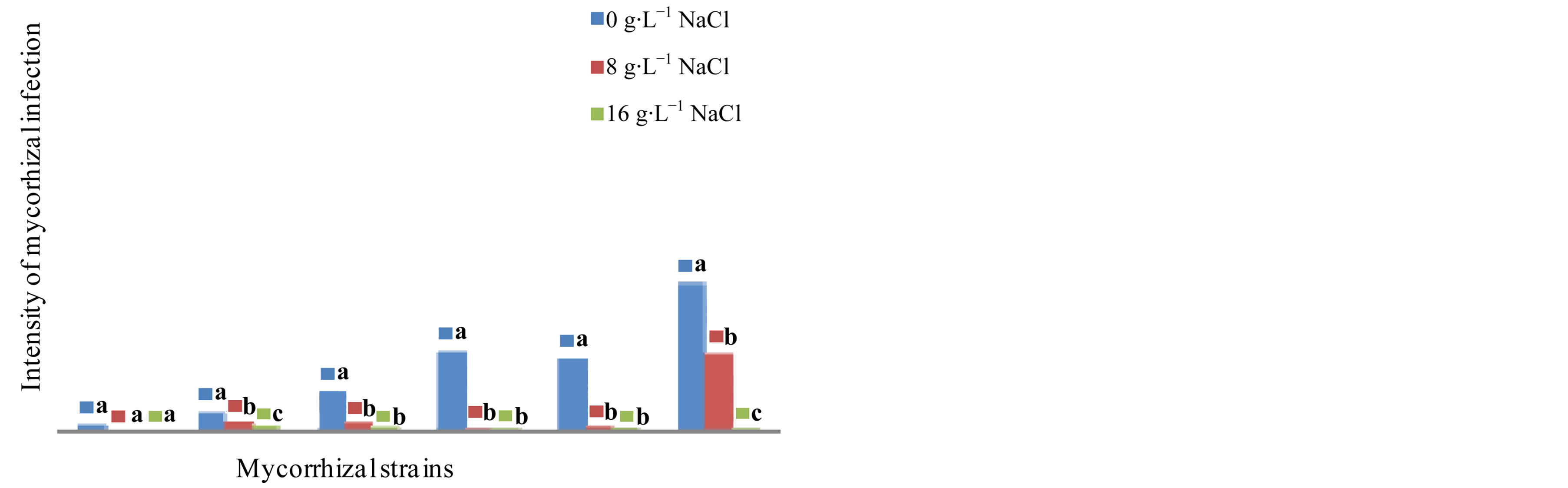
Figure 5. Impact of five strains of Glomus on the intensity of mycorrhizal plants in the cultivar Tijib in the presence of 0, 8 and 16 g∙L−1 after 4 months in greenhouse. Means that do not share a letter are significantly different at 5% using Student-Newman and Keuls’ test.
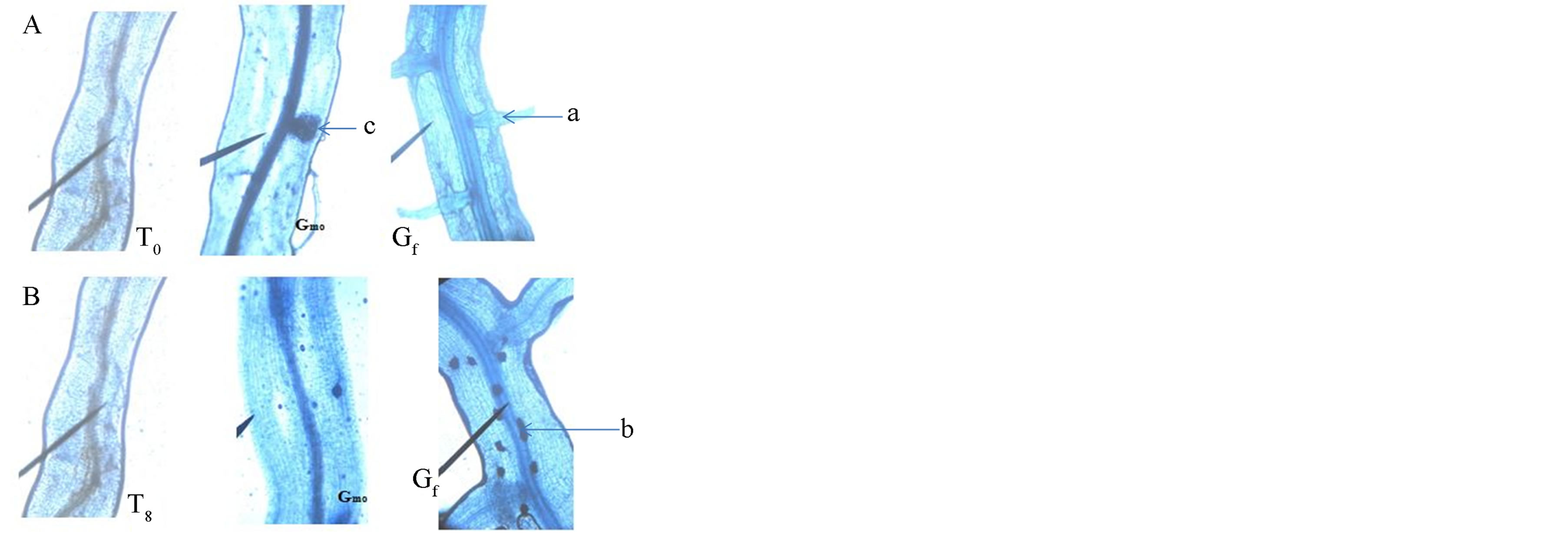
Photo 3. Colonization of plant roots by Tijib strains of fungi G. and G. mosseae fasciculatum in the absence of stress (A) and in the presence of salt (B). T0 (Witness) Gmo (G. mosseae) Gf (G. fasciculatum). In absence of stress, we observe the development of vesicles (c) and hyphae (a). In contrast, in presence of stress, only spores (b) are observed in mycorrhizal plants with the strain G. fasciculatum.
velopment of the epicotyl in NHH, with an average of 33.5 cm against 30.3 in controls (Table 2). While in Tijib is rather Glomus fasciculatum that promotes the growth of the aerial parts with an average of 33.8 cm against 31.4 cm for the control plants (Table 3).
3.4. Effect of Inoculation on the Growth of Stems of Plants in Cultivars NHH and Tijib Subjected to Salt Stress
The results show that G. intraradices significantly stimulated (F = 32.157, P = 0.000) stem growth of plants in the cultivar NHH when NaCl is used in the range 8 to 16 g∙L−1. Indeed, compared to control plants and all of the strains tested, G. intraradices allows obtaining the higher
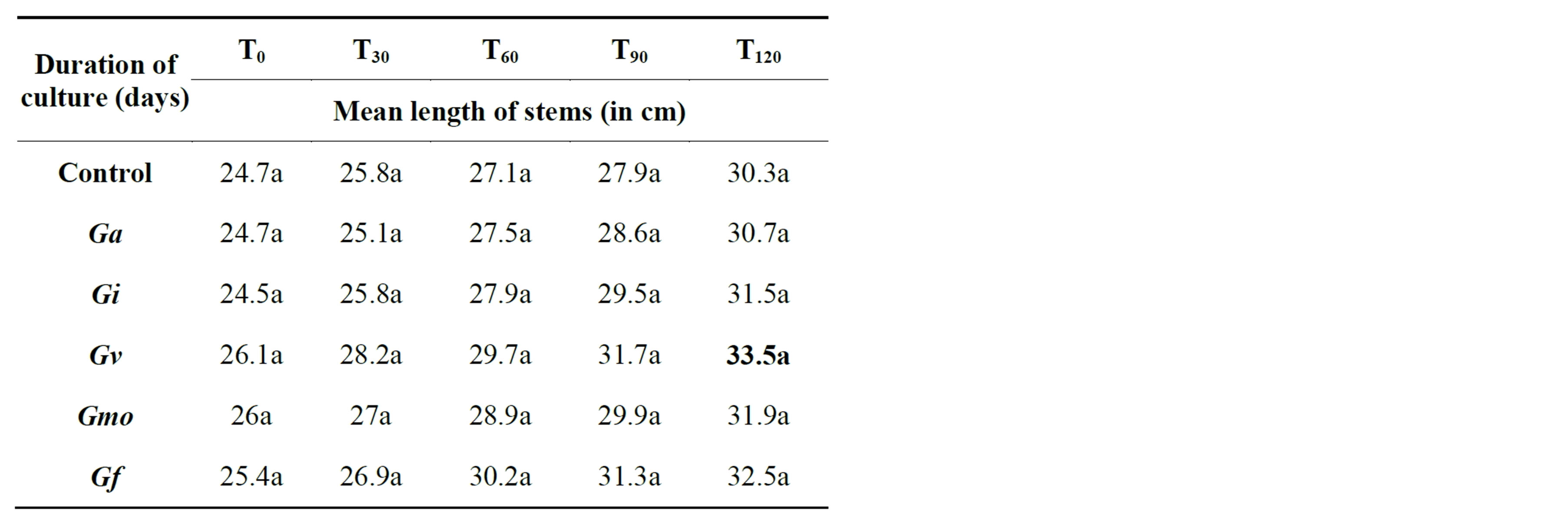
Table 2. Impact of inoculation on date palm plants stems growth in the le cultivar Nakhla hamra after 4 months in greenhouse. Means that do not share a letter are significantly different at 5% using Student-Newman and Keuls’ test.
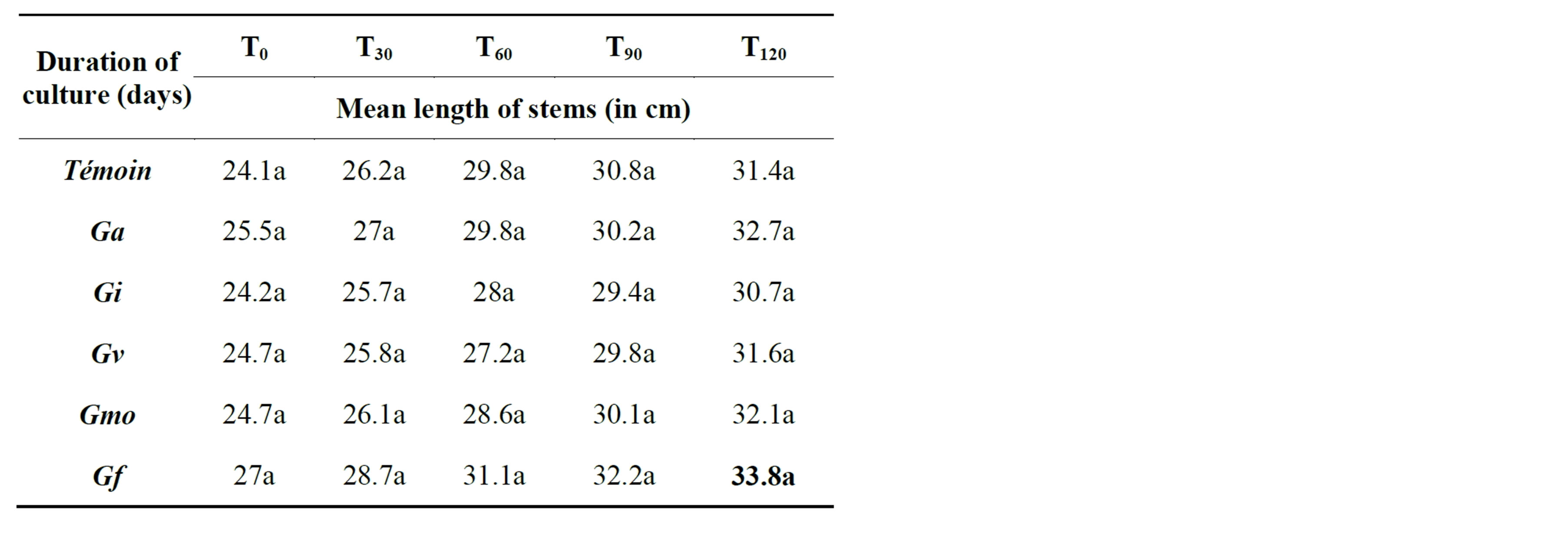
Table 3. Impact of inoculation on date palm plants stems growth in the cultivar Tijib after 4 months in greenhouse. Means that do not share a letter are significantly different at 5% using Student-Newman and Keuls’ test.
average growth of stems (32.16 cm) in presence of NaCl 8 g∙L−1. In contrast, in the cultivar Tijib, it is rather the strain G. fasciculatum that significantly stimulates (F = 17.299, P = 0.000) stem growth in presence of salt stress. Our results show that G. fasciculatum provides the longest stems (average 33.26 cm) in the presence of a NaCl concentration of 8 g∙L−1 compared to control plants and all tested strains (Table 4).
3.5. Effect of Salt Stress on Root Growth of Mycorrhizal Plants in NHH
The results presented in Figures 6 and 7 show the influence of mycorrhizal strains on the elongation of the pivot and the production of secondary roots in NHH after 4 months of culture in presence of different tested concentrations of NaCl. The analysis of variance, clearly indicate that there is as well variety effect (F = 15.370, P = 0.000), as medium effect (F = 21.979, P = 0.000) and a very significant interaction variety x mycorrhizal strains (F = 9.268, P = 0.000) effect on the elongation of the pivot and the production of secondary roots. Inoculation
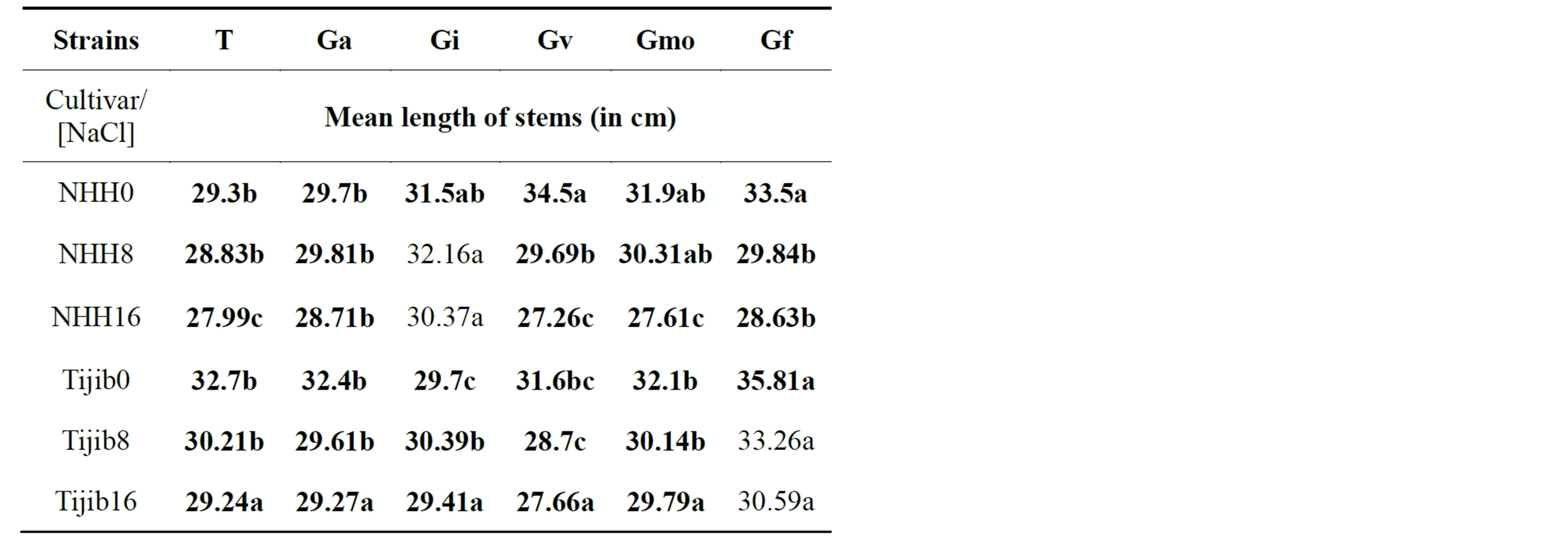
Table 4. Impact of inoculation on date palm plants stems growth in the cultivars NHH and Tijib after 4 months in greenhouse. Means that do not share a letter are significantly different at 5% using Student-Newman and Keuls’ test.
of plants with the strain G. intraradices stimulates root elongation in NHH varying from an average of 73 cm in the control to 77 cm in cultivated plants in presence of 8 g∙L−1 of NaCl (Figure 6). The same is true for the production of secondary roots that vary from 42 roots per plant averaged 50 in stressed plants (Figure 7). In contrast, G. verriculosum, stimulates the production of secondary roots (52 roots on average) and the elongation of the taproot (81.5 cm on average) but inhibits root growth in presence of salt. At 16 g∙L−1 in the culture medium, NaCl has a depressive effect on the growth and production of roots whatever the strain used.
3.6. Effect of Salt Stress on Root Growth of Mycorrhizal Plants in Tijib
The results show the influence of mycorrhizal strains on the elongation of the taproot (Figure 8) and the production of secondary roots (Figure 9) in Tijib after 4 months of culture in the presence of different concentrations of tested NaCl. The analysis of variance indicates that there is a variety effect (F = 14.410, P = 0.000), and also a medium effect (F = 12.817, P = 0.000) and a high significant interaction variety x strains mycorrhizal (F = 8.629, P = 0.000) effect on the elongation of the taproot and the production of secondary roots. In presence of NaCl 8 g∙L−1, the inoculation of Tijib plants significantly reduced root elongation that is less than the average length of non-inoculated plants (53.27 cm). However, at the same salt concentration, the production of secondary roots is stimulated by inoculation with G. fasciculatum the average number of changes including 42.5 roots/plant in controls to 45 in inoculated plants.
3.7. Effect of NaCl on the Accumulation of Proline in NHH and Tijib Plants
The results of the analysis of variance revealed not only a medium effect (F = 10.62, P = 0.000) but also interaction cultivar x inoculation conditions (F = 29.873, P = 0.000) effect very significant on proline production in the culti-
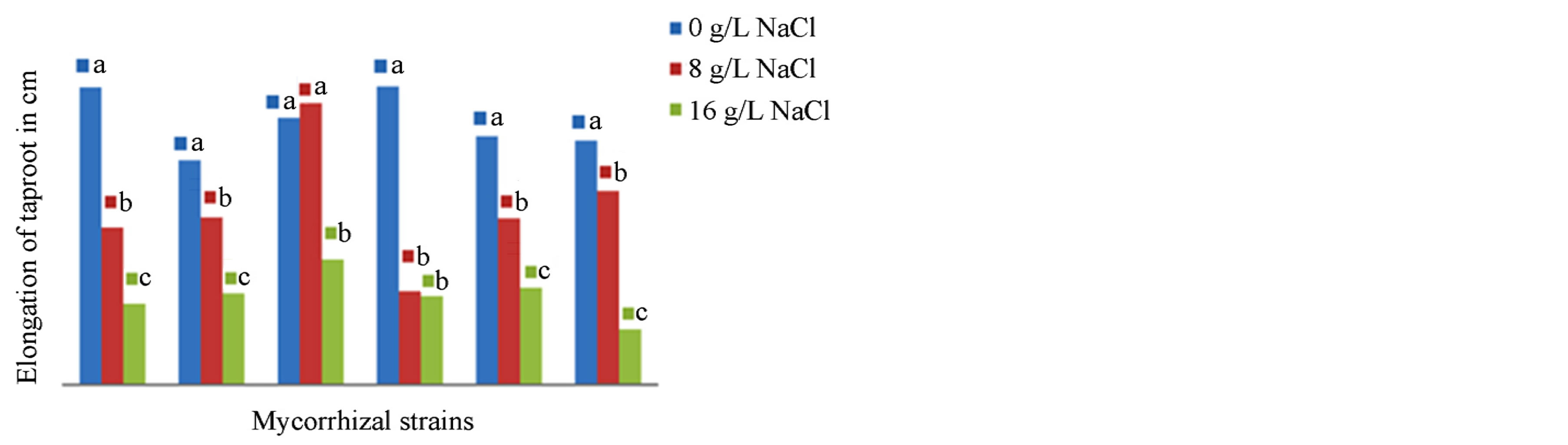
Figure 6. Impact of five strains of Glomus on the taproot elongation in the cultivar NHH in presence of 0, 8 and 16 g∙L−1 after 4 months in greenhouse. Means that do not share a letter are significantly different at 5% using Student-Newman and Keuls’ test.
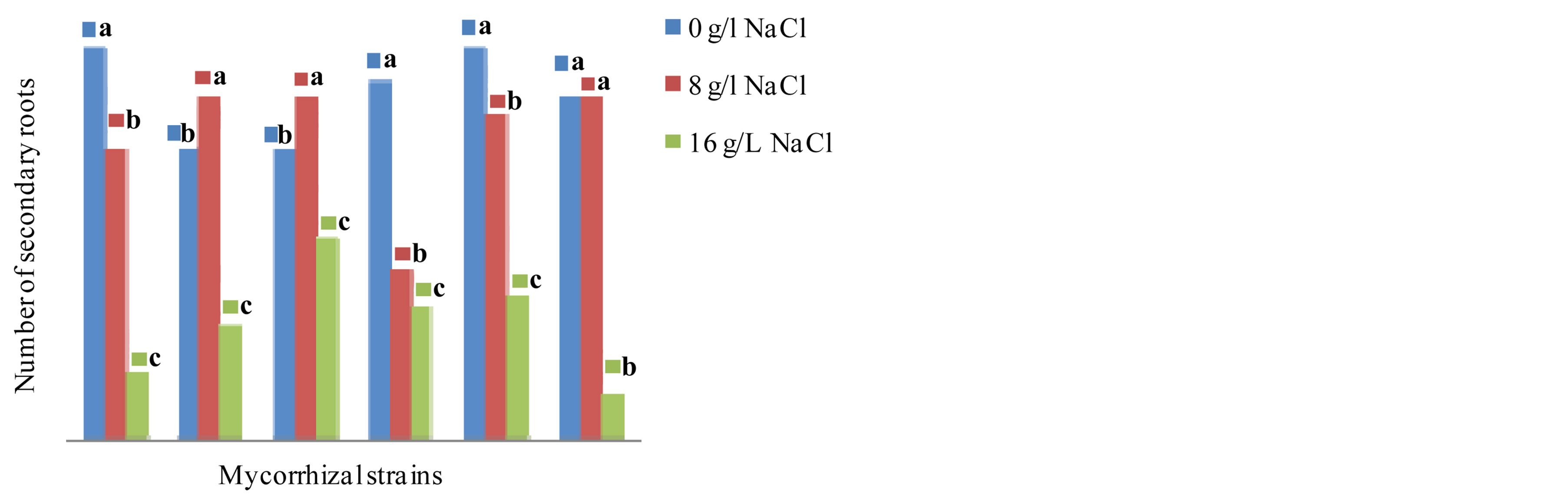
Figure 7. Impact of five strains of Glomus on the production of secondary roots in the cultivar NHH in presence of 0, 8 and 16 g∙L−1 of NaCl after 4 months in greenhouse. Means that do not share a letter are significantly different at 5% using Student-Newman and Keuls’ test.
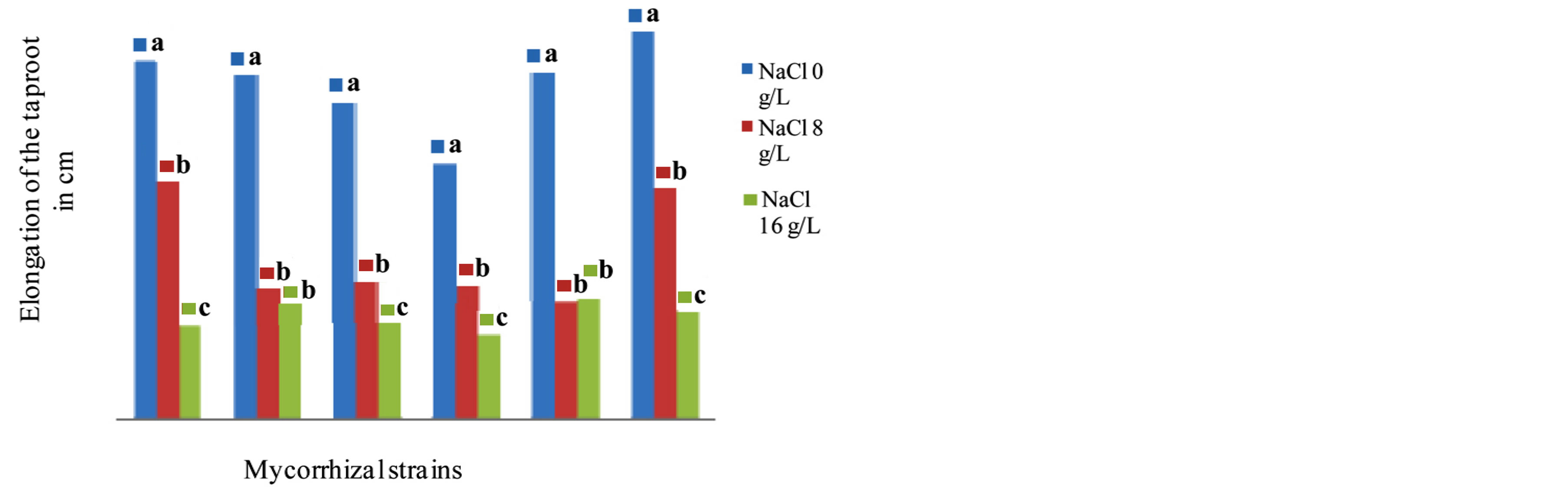
Figure 8. Impact of five strains of Glomus on the taproot elongation in the cultivar Tijib in presence of 0, 8 and 16 g∙L−1 after 4 months in greenhouse. Means that do not share a letter are significantly different at 5% using Student-Newman and Keuls test.
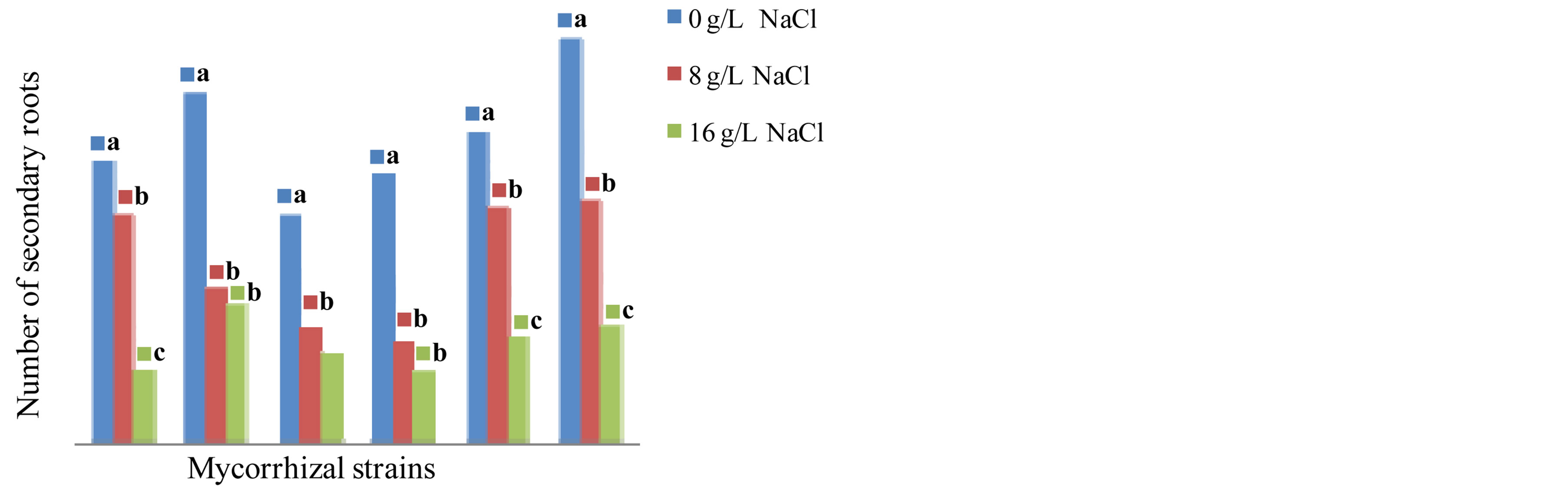
Figure 9. Impact of five strains of Glomus on the secondary roots production in the cultivar Tijib in presence of 0, 8 and 16 g∙L−1 after 4 months in greenhouse. Means that do not share a letter are significantly different at 5% using Student-Newman and Keuls’ test.
vars NHH and Tijib. The results (Figure 10) show that upon application of salt stress, seedlings of the cultivar NHH quickly and intensely accumulate proline. Indeed, the contents of proline appear 1.75 times to 4 times higher, respectively, in the presence of 8 and 16 g∙L−1 of salt compared to control plants when inoculated with G. intraradices. In contrast, in the cultivar Tijib, accumulation of proline is more intense and faster when plants are inoculated with G. fasciculatum (Figure 11). In this cultivar, the proline content increased by 700 to 1500 nmoles∙g−1 fresh matter at 8 g∙L−1 and 1800 nmoles∙g−1 fresh matter in presence of 16 g∙L−1 NaCl.
4. DISCUSSION
The defined culture conditions allowed us to test the degree of NaCl tolerance in seedlings of two cultivars of date palms Sahelian selected for earliness in flowering and fruiting. In our study, the intensity of mycorrhiza and growth and proline biosynthesis in plants of date palm appeared strongly dependent on cultivar, inoculation conditions and levels of NaCl in the culture environment. Indeed, the results showed that upon reaching the threshold of tolerance to salt, NaCl significantly reduces the growth of date palm seedlings in greenhouse. Our results confirm those obtained in in vitro culture conditions by [7] who found the same depressive effect of NaCl in both cultivars but with a better tolerance of plants of cultivar Tijib to salt stress compared to the cultivar NHH when salt concentrations exceed 8 g∙L−1. The results also appear similar to those reported by [17] and [18]. These authors, in fact, found respectively as NaCl greatly reduces the growth of seedling epicotyl in Vigna radiata as well as the growth and seed production in Tunisian barley accessions. The work of [8] and those of [9] showed that arbuscular mycorrhizal (AM) fungi can grow naturally in various saline environments where they can im-
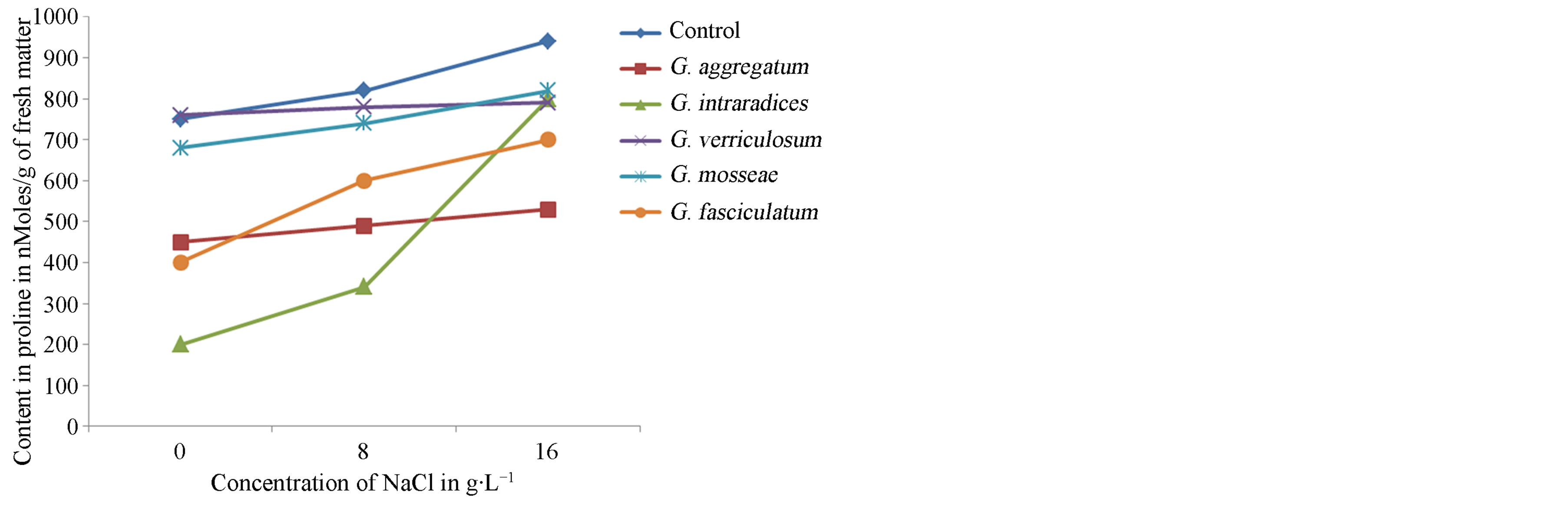
Figure 10. Effect of NaCl concentration on proline production in plants of cultivar NHH after 4 months in greenhouse.
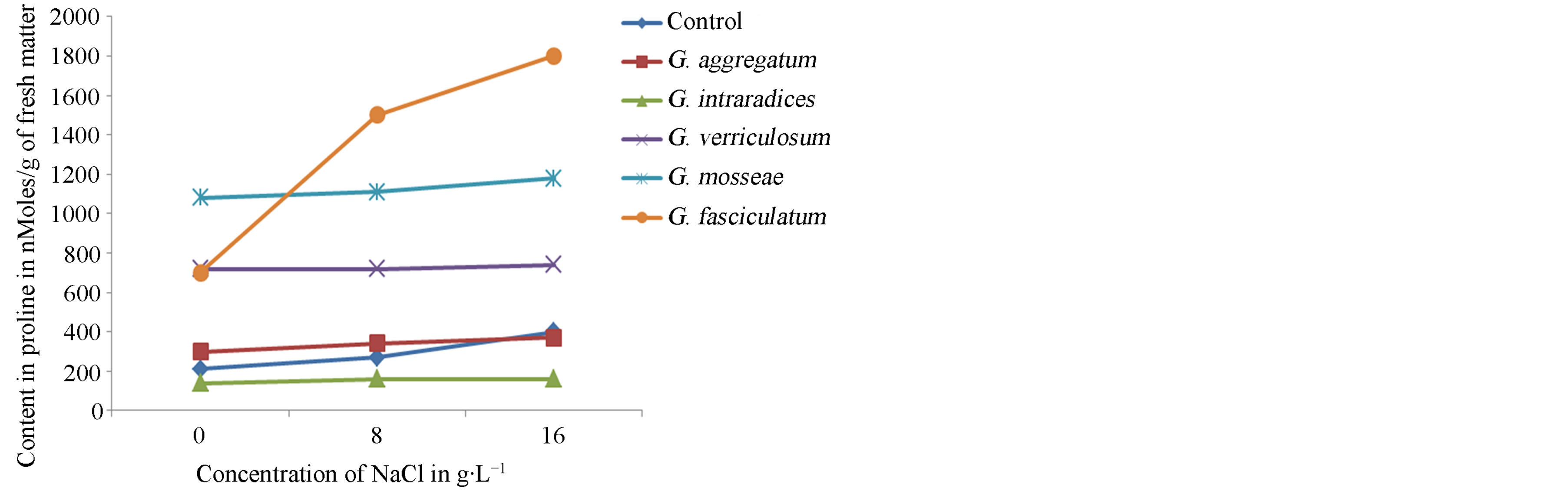
Figure 11. Effect of NaCl concentration on proline production in plants of cultivar Tijib after 4 months in greenhouse.
prove plant growth. [10] determined that the AM fungi generally observed in saline soils belong to the genus Glomus and order Glomerales.
Under our experimental conditions, the analysis parameters and mycorrhizal growth of date palm seedlings allowed us to highlight the existence of symbiotic couples in which the intensity of mycorrhiza and growth of stems and roots of the plants have been optimized in presence or not of salt stress. Indeed, in the absence of salt stress, the average optimal values of the intensity of mycorrhizal (47.54%) and plant growth in terms of shoot length (34.5 cm) and roots (81.5 cm ) and number of roots (57.5 roots/plant) are observed in cultivar NHH when plants were inoculated with G. verriculosum. In contrast, in Tijib, the intensity of colonization of the root system (16%) and stem (35.8 cm) and roots (87 cm) elongation, and the average production of roots (75 roots/plant) are enhanced in the presence of G. fasciculatum. In the presence of salt stress, it is rather with the strain G. intraradices that seedlings of NHH have not only the higher mycorrhizal intensity (28%) but also longer stems (32.16 cm) and roots (77 cm) and stronger root production (50 roots/plant).
However, in Tijib, the values of mycorrhizal intensity (8.10%) and stem growth (33.26 cm) and roots (51.62 cm) and the average production of roots (45 roots/plant) are optimized when the plants were inoculated with G. fasciculatum. Many studies have also shown that AM fungi increase the salt tolerance of plants. These results are similar to our findings. This tolerance is manifested through the implementation of various mechanisms, such as improving the absorption of water and phosphorus [19], the production of growth hormone [20], improved soil mycorrhizal conditions (Lindremann, 1994), changes in physiological and biochemical properties of the host plant [21], 2008) and root protection against toxic ions [22]. In addition, fungi cause greater nitrogen assimilation in the host plant under salt stress [23].
However, the inoculation of cultivars NHH and Tijib with strains of G. aggregatum, G. mosseae showed no benefits of AM fungi on the growth of plants. [24] also found a negative effect of inoculation with Gigaspora albida and Scutellospora heterogama on the intensity of mycorrhiza and growth of Passiflora edulis in unsterilized soil. The absence of response [25] and inhibition of plant growth [26] have also been observed under saline conditions. Under these conditions, it appears that spore germination is inhibited or delayed by increasing concentrations of NaCl [27]. The accumulation of proline has been demonstrated in many species and in different situations of water deficit stress in Sesuvium portulacastrum [28] and salinity in Atriplex L. halimus [29]. In our culture conditions, the date palm mycorrhizal seedlings and subjected to salt stress accumulate high concentrations of proline. Our results showed a strong interaction between cultivar and mycorrhizal strains on proline biosynthesis. They establish that upon application of salt stress, seedlings of the cultivar NHH quickly accumulate proline which contents appear 1.75 times to 4 times higher than controls when plants were inoculated with Glomus intraradices and grown respectively 8 and 16 g∙L−1 of salt. In contrast, in the cultivar Tijib, accumulation of proline appeared more intense and faster when plants were inoculated with Glomus fasciculatum. G. intraradices and G. fasciculatum also appeared in our experimental conditions, such as AM fungi that better colonize the root system of the cultivars NHH and Tijib inducing growth of plants. These strains would then push the limits of salt tolerance in NHH and Tijib plants in the range of tested NaCl. With the exception of these strains, our results showed an almost stabilizing proline contents for other conditions inoculation. This suggests that the limits of salt tolerance are achieved for these conditions. The results of salt stress in relation to the accumulation of proline in two date palm cultivars studied are in good agreement with those presented in the millet by [30]. These authors, in fact, established a positive correlation between the degree of plant resistance to stress and kinetics of accumulation of proline. This kinetic is fast and temporary susceptible species when it appears later inresistant species. Accumulation is a form of adaptation to osmotic stress. However, in our experimental conditions, it was not possible to establish a correlation between the improvement of the effect of inoculation and the accumulation of proline. These results confirm the observations of [12] in Acacia mangium. In order to refine this study, it would be interesting to complement these results by assays of glutamine synthetase activity, enzyme involved in the biosynthesis of glutamic acid which acts as precursor in the biosynthesis of proline [31].
5. CONCLUSION
Date palm cultivation is today one of the main activities on which lies much of the economy of the Sahel. It appears severely compromised by two main constraints: soil salinization and drought. These constraints have led to the virtual disappearance of some cultivars contributing to the depletion of the gene pool in the Sahel. The above results contribute to a better understanding of physiological adaptation mechanisms implemented in the date in under salt stress. The impact of arbuscular mycorrhizal fungi has significantly improved the growth of date palm seedlings under conditions of salt stress. The strategy adopted in this study could be extended to genotypes of interest to provide, ultimately, a better counseling of farmers on the choice of cultivars to be used for the installation of palm trees in areas where salt or water is a limiting factor.
ACKNOWLEDGEMENTS
We thank the FIRST that funded the project, M. Saleck and the University of Nouakchott (Mauritania), facilitated the access to germplasm in the Atar region. M. Jean Bakhoum for his technical assistance, Dioumacor Fall and Ibou Diop for their support during the mixture. The authers thank the Support and Training Department (DSF) of the French Institut de Recherche pour le Developpement (IRD) for granting this work.
REFERENCES
- Munier, P. (1973) Le palmier dattier. Techniques Agricoles et Productions Tropicales. Paris 5eme, Maisonneuve et Larose, 217.
- Heselmans, M. (1997) Setting research priorities through an international date palm network. Biotechnology and Development Monitor, 30,18-20.
- Ould Sidina, C. (1999) Présentation des oasis mauritaniennes. In: Agroéconomie des Oasis, Groupe de Recherche et d’Information pour le Développement de l’Agriculture d’Oasis (GRIDAO-CIRAD), eds, 49-51.
- Barbiero, L., Mohamedou, A. O., Laperrousaz, C., Furian, S. and Cunnac, S. (2004) Polyphasic origin of salinity in the Senegal delta and middle valley. Catena, 58, 101-124. http://dx.doi.org/10.1016/j.catena.2004.03.003
- F.A.O. (2005) Global network on integrated soil management for sustainable use of saltaffected soils.
- Ferry, M., Bouguedoura, N. and El Hadrami, I. (1998) Patrimoine génétique et techniques de propagation in vitro pour le développement du palmier dattier. Sécheresse, 9, 139-146.
- Sané, D., Ould Kneyta, M., Diouf, D., Diouf, D., Badiane, F. A., Sagna, M. and Borgel, A. (2005) Growth and development of date palm (Phoenix dactylifera L.) seedlings under drought and salinity stresses. African Journal of Biotechnology, 4, 968-972.
- Yamato, M., Ikeda, S. and Iwase, K. (2008) Community of arbuscular mycorrhizal fungi in coastal vegetation on Okinawa Island and effect of the isolated fungi on growth of sorghum under salt-treated conditions. Mycorrhiza, 18, 241-249. http://dx.doi.org/10.1007/s00572-008-0177-2
- Daei, G., Ardekani, M.R., Rejali, F., Teimuri, S. and Miransari, M. (2009) Alleviation of salinity stress on wheat yield, yield components, and nutrient uptake using arbuscular mycorrhizal fungi under field conditions. Journal of Plant Physiology, 166, 617-625. http://dx.doi.org/10.1016/j.jplph.2008.09.013
- Wang, F.Y., Liu R.J., Lin, X.G. and Zhou, J.M. (2004) Arbuscular mycorrhizal status of wild plants in salinealkaline soils of the Yellow River Delta. Mycorrhiza, 14, 133-137. http://dx.doi.org/10.1007/s00572-003-0248-3
- Diop, T. A., Guèye M., Dreyfus, B. L., Plenchette, C. and Strullu, D. G. (1994) Indigenous arbuscular mycorrhizal fungi associated with Accacia albida Del. In different areas of Senegal. Applied and Environmental Microbiology, 60, 3433-3436.
- Diouf, D., Forestier, S., Neyra, M., and Lesueur, D. (2003) Optimisation of inoculation of Leucaena leucocephala and Acacia mangium with rhizobium under greenhouse conditions. Annals of Forest Science, 60, 379-384. http://dx.doi.org/10.1051/forest:2003029
- Phillips, I.M. and Hayman, D.S. (1970). Improved procedures for clearing and staining parasitic and vesiculararbuscular mycorrhizal fungi for rapid assessement of infection. Transactions of the British Mycological Society, 55, 158-161. http://dx.doi.org/10.1016/S0007-1536(70)80110-3
- Giovannetti, M. and Mosse, B. (1980) An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New physiologist, 84, 489-500. http://dx.doi.org/10.1111/j.1469-8137.1980.tb04556.x
- Trouvelot, A., Kough, J.L. and Gianinazzi-Pearson, V. (1986) Mesure du taux de mycorhization ayant une signification fonctionnelle. Dans: Aspects physiologiques et génétiques des mycorhizes, Dijon, 1985. INRA (éd.), 217-221.
- Monneveaux, P. and Nemmar, M. (1986) Contribution à l’étude de la résistance à la sécheresse chez le blé tendre (Triticum aestivum L.) et le blé dur (Triticum durum Desf.). Etude de l’accumulation de proline au cours du développement. Agronomie, 6, 583-590. http://dx.doi.org/10.1051/agro:19860611
- Promila, K. and Kumar, S. (2000) Vigna radiata seed germination under salinity. Biol. Plant, 43, 423-426. http://dx.doi.org/10.1023/A:1026719100256
- Cheikh M’Hahmedi, H., Abdellaoui, R., Kadri, K., Ben Naceur, M. and Bel Hadj, S. (2008) Evaluation de la tolérance au stress salin de quelques accessions d’orge (Hordeum vulgare l.) cultivées en Tunisie: Approche physiologique. Sciences & Technologie C, 28, 30-37.
- Ruiz-Lozano, J.M. (2003) Arbuscular mycorrhizal symbiosys and alleviation of osmotic stress. New perspectives for molecular studies. Mycorrhiza, 13, 309-317. http://dx.doi.org/10.1007/s00572-003-0237-6
- Fortin, J. A., Plenchette, C. and Piché, Y. (2008) Les mycorhizes, la nouvelle révolution verte. Eds. Multimondes et Quae, 131.
- Smith, S. E. and Read, D. J. (2008) Mycorrizal symbiosis, 3rd Edition, Academic Press, Waltham.
- Hammer, E. C., Nasr, H., Pallon, J., Olsson, P. A. and Wallander, H. (2010) Elemental Composition of Arbuscular Mycorrhizal Fungi from with Excessive Salinity. In: Tryckeriet I E-hyset, Lund. Nutriment Balance and Salinity Stress in Arbuscular Mycorrhizal Fungi, 105-122.
- Evelin, H., Kapoor, R. and Giri, B. (2009) Arbuscular mycorrhizal fungi in alleviation of salt stress: A review. Annals of Botany, 104, 1263-1280. http://dx.doi.org/10.1093/aob/mcp251
- Cavalcante, U.M.T., Maia, L.C., Costa, C.M.C and Santos, V.F. (2001) Mycorrhizal dependency of passion fruit (Passiflora edulis f. flavicarpa). Fruits, 56, 317-324. http://dx.doi.org/10.1051/fruits:2001132
- Graham, J.H. and Syvertsen, J.P. (1989) Vesicularar-buscular mycorrhiza increase chloride concentration in citrus seedlings. New Phytologist, 113, 29-36. http://dx.doi.org/10.1111/j.1469-8137.1989.tb02392.x
- Coperman, R.H., Martin, C.A. and Sutz, J.C. (1996) Tomato growth in response to salinity and mycorrhizal fungi from saline or nonsaline soils. Hortscience, 31, 341-344.
- Juniper, S. and Abbott, L.K. (2006) Soil salinity delays germination and limits growth of hyphae from propagules of arbuscular mycorrhizal fungi. Mycorrhiza, 16, 371-379. http://dx.doi.org/10.1007/s00572-006-0046-9
- Slama, I., Messedi, D., Ghnaya, T. and Abdely, C. (2004) Effet du déficit hydrique sur la croissance et l’accumulation de la proline chez Sesuvium portulacastrum. Revue des Régions Arides, 1, 234-241.
- Bidai, Y. (2001) Le métabolisme de la proline chez l’Atriplex halimus L. stressée à la salinité. Mémoire de magister en Physiologie Végétale, Université Es-Senia, Oran.
- Hubac, C. and Viera and Da Silva, J. (1980) Indicateurs métaboliques de contraintes mésologiques. Physiologie Végétale, 18, 45-53.
- Boggess, S.F. and Stewart, C.R. (1976) Contribution of arginine to proline accumulation in water stressed barley leaves. Plant Physiology, 58, 796-797. http://dx.doi.org/10.1104/pp.58.6.796
NOTES
*Corresponding author.