American Journal of Plant Sciences
Vol.05 No.21(2014), Article ID:50881,27 pages
10.4236/ajps.2014.521337
Soil and Phytosociological Characterisation of Grasslands in the Western Mediterranean
A. Cano-Ortiz1, E. Bioindi2, C. J. Pinto Gomes3, S. Del Río González4, E. Cano5
1Department of Interra Sustainability, Resources Engineering and SL Spain Square, Salamanca, Spain
2Department of Environmental Sciences and Crop Production, Polytechnic University of Marche, Ancona, Italy
3Department of Landscape, Environment and Planning/Institute of Agricultural and Environmental Sciences Mediterranean (ICAAM), University of Évora (Portugal), Rua Romão Ramalho, Portugal
4Department of Biodiversity and Environmental Management (Department of Botany), Livestock Mountain Institute (CSIC-ULE Joint Center), School of Biological and Environmental Sciences, University of León, Vegazana Campus, León, Spain
5Department of Animal Biology, Plant and Ecology, Botany, University of Jaén, Jaén, Spain
Email: ana@interra.es, e.biondi@univpm.it, cpgomes@uevora.pt, sriog@unileon.es, ecano@ujaen.es
Copyright © 2014 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 10 August 2014; revised 13 September 2014; accepted 18 October 2014
ABSTRACT
A study was made of grasslands in olive groves with a high frequency of Hordeum leporinum Link, Chrysanthemum coronarium L., Malva neglecta Wallr., Aegilops geniculata Roth and Dasypyrum villosum (L.) Borbas, initially included in the alliance Hordeion leporini Br.-Bl. in Br.-Bl., Gajewski, Wraber & Walas 1936 corr. O. Bolós 1962, and sub-alliance Malvenion neglectae Gutte 1966. The study was carried out in the western Mediterranean (Spain, Italy, Portugal and northern Morocco). The soil and phytosociological treatment of the samples contributes information for new plant communities. In the study of plant communities, the indices of degree of presence are adapted to those of Van der Maarel. The new alliance Securigero securidacae-Dasypyrion villosi is proposed for the sub-Mediterranean and Euro-Siberian territories, with the new communities Securigero securidacae-Dasypyretum villosi and Convolvulo elegantissimi-Aegilopetum geniculatae. This last community, due to its richness in Aegilops geniculata Roth, could be included in the alliance Tae- niathero-Aegilopion geniculatae Rivas-Martínez & Izco 1977; however, the soil and statistical ana- lyses reveal that it should remain part of the alliance Securigero securidacae-Dasypyrion villosi. The soil and floristic study of Hordeion leporini and Malvenion neglectae allows us to describe the new sub-alliance Resedo albae-Chrysanthemenion coronarii, subordinated to Malvion neglectae (Gutte 1966) status novo.
Keywords:
Soil, Grassland, Community, Association

1. Introduction
The grassland communities studied are included by various authors [1] - [3] , in the alliance Hordeion leporini, which comprises grasslands with a Mediterranean optimum, a sub-nitrophilous or nitrophilous character, and spring growth. This alliance extends throughout the sub-Mediterranean and central European territories. Although it has a Mediterranean optimum, this alliance can be found in Euro-Siberian (centre-European) areas such as the central Pyrenees (Spain) and the central Apennines (Italy), with a presence, in these latter territories and in Italian sub-Mediterranean environments, of species such as Dasypyrum villosum (L.) Borbas. These cal- cicolous species from southeast Europe are frequently found in Italy, and are absent in Spain and Portugal. The same occurs with Securigera securidaca (L.) Deg. et Dorfl., whose area of distribution is in southern Europe un- til southeast France; and Crepis sancta (L.) Babc., found in the samples taken in Italy, with a biogeographic area in the eastern Mediterranean region and in southeast Europe. All the communities of Dasyyirum villosum (L.) Borbas are grassland formations with dense coverage and considerable biomass which thrive in environments with a high quantity of organic matter [4] . This alliance includes some communities which, due to their structure, floristic composition and soil, should have been included in alliances other than Hordeion leporini. Alliance has been studied by other authors [5] - [10] . The soil parameters condition the presence of plant populations and communities, and this fact can be used to determine the soil nutrients according to the degree of presence and abundance of the species.
2. Material and Methods
Study Area
The territory in the study corresponds to the western Mediterranean (Spain, Italy, Portugal and northern Moroc- co). The samplings were taken in the spring of 2006. This study was initiated by [6] [7] in Spain and Portugal, and the Italian areas in Umbria and the Puglia region (Gargano) were inventoried by [5] . The communities in the sampling are distributed throughout the western Mediterranean, and the soil samples were taken in Andalusia (Spain), Alentejo (Portugal) and the regions of Umbria and Gargano (Italy) (Figure 1). Soil samples were taken of substrates with both acid and basic pH, when the plant community presented a high abundance index of the dominant species.
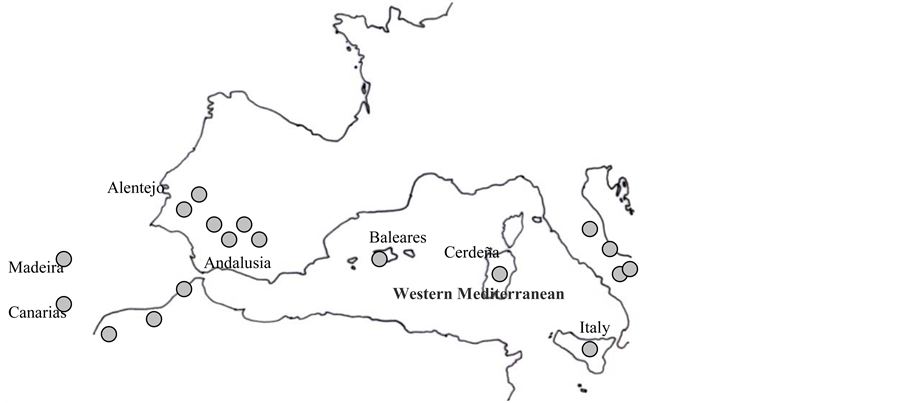
Figure 1. Localisation of soil sampling on the Iberian Peninsula and in Italy.
The floras of [11] and [12] , were used for the floristic and phytosociological study, and [13] for the phytoso- ciological methodology. The soil study was done by taking 265 samples of each inventory of 14 plant communi- ties, and the soil sample was obtained from the plant community where the dominant species has a high abun- dance-dominance index, and at the depth of the root system of each species. 16 soil parameters were analysed in the laboratory: exchangeable calcium (meq/100g), cation exchange capacity (meq/100g), carbonates (%), assimilable phosphorus (p.p.m), exchangeable magnesium (meq/100g), oxidable organic matter (%), total nitrogen (%), pH 1/2.5, exchangeable potassium (meq/100g), pF 1/3 atm. (%), pF 15 atm. (%), electrical conductivity 1/5 (mmhos/cm), clayey texture (%), silty texture (%), sandy texture (%), and exchangeable sodium (meq/100g).
In order to differentiate the communities of Hordeion leporini in the western Mediterranean, a synthetic Excel chart was drawn up based on the 74 association tables which include 847 inventories (Table 1), and comprise 740 rows (species) × 74 columns (association tables). The degrees of presence used were according to [13] , which range from I to V with the following % equivalence: V implies that the degree of presence of the species in the samples taken ranges between 80% and 100%; IV from 60% to 80%, III from 40% - 60%, II from 20% - 40% and I < 20%. In our case we applied the following formula in order to obtain the index value (degree of presence): Ip = n. V/T, where Ip = index of presence; n = number of inventories in the table of the community in which the species is included; V = maximum presence value (the species is included in all the inventories in the community table); T = total number of inventories in the table. Based on this criterion we used the following Ip values; an Ip value of >4.5 is assigned a presence value of V; Ip = 3.5 - 4.5 a value of IV; Ip = 2.5 - 3.5 a value of III; Ip = 1.5 - 2.5: II; and finally Ip < 1.5 is assigned a value of I. Once the synthetic table of the 74 commu- nity tables was drawn up, we adapted these indices to the [42] . I = 3; II = 4; III = 5; IV = 6; V = 7.
A cluster was applied to the Excel chart of 740 rows (species) × 74 columns (association tables) with the Jac- card distance, in order to establish the degree of similarity/dissimilarity between the association tables. It was similarly applied to the synthetic species tables for Italy, the Iberian Peninsula and northern Morocco (Spain, Portugal, northern Morocco), which comprise 444 rows × 26 columns and 528 rows × 47 columns respectively, to determine the distance existing between the different associations present in each territory. The previous analysis was corroborated with DECORANA ordination and RA analysis to establish the difference between the groups.
A table of averages was drawn up using the 265 soil samples and a PCA was applied to determine which soil factors condition the presence of the plant community.
In order to differentiate the Italian communities rich in Dasypyrum villosum (L.) Borbas, a statistical treat- ment of conglomerates was performed. This was done by drawing up several Excel charts; one with the 45 phytosociological field samples plus 63 inventories of proximate associations due to their richness in Dasypyrum villosum (L.) Borbas (281 rows × 108 columns), to which was subsequently applied a cluster analysis using the Euclidean distance.
In order to determine in which alliance to include the Italian community of Convolvulus elegatissimus Miller and Aegilops geniculata Roth, two Excel charts were made with the 45 soil and community samples from Italy plus 41 from the Iberian Peninsula which were very similar to the Italian samples in terms of structure and flo- ristic composition, and comprising 16 soil parameters × 86 columns (samples), with subsequent application of cluster and PCA. The 41 Iberian samples belonged to the associations Trifolio cherleri-Taeniatheretum capitis- medusae Rivas-Martínez & Izco 1977 (TT) and Plantago bellardii-Aegilopetum geniculatae Cano-Ortiz, Pinto Gomes & Cano 2010 (PA), described for Portugal [8] .
The data used were included in a synthetic table of presences with 740 rows (species) × 74 association tables, and in a soil table of 265 samples × 16 soil parameters.
3. Results and Discussion
A statistical treatment was performed on 12 of the 16 soil parameters analysed in each one of the 265 samples, as two of the soil parameters obtained did not appear to be relevant (Table 2). This allows us to confirm that MOO, Nt and CIC are the factors which have the greatest influence in communities AB, SD and CA present in Italy (Ge1) (Figure 2 and Figure 3), with a lower influence of Pa, Mgc and Kc; conversely in the group of grassland communities present on the Iberian peninsula, TT, LR, ArH, BH, ArChr, PA and ArP, the most rele- vant factor is texture (Ge2). The edaphic study of 14 communities served to establish the abundance of the do-

Table 1. Associations studied in the western Mediterranean.

Figure 2. Soil parameters with the greatest influence on plant communities.

Figure 3. Soil parameters with the greatest influence on plant communities. PCA analysis.
Table 2. Average values of the soil parameters studied.
CIC = cation exchange capacity in meq/100g; MOO = oxidable organic matter in %; Nt = total nitrogen in %; Pa = assimilable phosphorus in ppm; Mgc = exchangeable magnesium in meq/100g; Kc = exchangeable potassium in meq/100g; pF 15 atm = pressure at 15 atm (water retention capacity) in %; Tx arc = clayey texture in %; Tx ar = sandy texture in %; Tx lim = silty texture in %; EC = conductivity mmhos/cm; pH. 1AcH.-Anacyclo clavati-Hordeetum leporini Cano-Ortiz et al. 2009. 2ArH.-Anacyclo radiati-Hordeetum leporinii O. Bolòs & Rivas-Martínez in Rivas-Martínez 1978. 3BH.-Bromo scoparii-Hordeetum leporinii Rivas Martínez 1978. 4LR.-Co. Linaria spartea et Raphanum raphanistrum. 5PD.-Papaveri rhoeadis-Diplotaxietum virgatae Rivas Martínez 1978. 6ArP.-Anacyclo radiati-Papaveretum rhoeadis Cano-Ortiz et al. 2009. 7ArChr.-Anacyclo radiati-Chrysanthemetum coronarii (Rivas-Martínez 1978) Cano- Ortiz et al. 2009. 8RChr.-Resedo albae-Chrysanthemetum coronarii O. Bolòs & Molinier 1958. 9UM.-Urtico urentis-Malvetum neglectae (Knapp) Lohmeyer in Tüxen 1950. 10AB.-Aveno barbatae-Brometum diandri Baldoni & Biondi 1993. 11CA.-Convolvulo elegantissimi-Aegilopetum geniculatae. 12SD.- Securigero securidacae-Dasypyretum villosii. 13PA.-Plantagini bellardii-Aegilopetum geniculatae Cano-Ortiz, Pinto & Cano 2010. 14TT.-Trifolio cherleri- Taeniatheretum capitis-medusae Rivas-Martínez & Izco 1977.
minant species of each association, and will be essential for future studies on soil indicators. This work refers mainly to the description of new syntaxa.
In the PCA analysis (Figure 4) we have included 14 soil parameters and 86 samples, 45 of which correspond to the communities AB, CA and SD, plus 41 to the Iberian communities PA and TT. In the PCA correlation analysis, the Italian grasslands AB, CA and SD are grouped on one side and the Iberian grasslands are grouped on the other, thus highlighting the fact that although the CA community in Italy has a structure close to the communities of Taeniatherum caput-medusae (L.) Nevski and Aegilops geniculata Roth, its floristic composition and the substrate on which this community grows allows us to include it with the rest of the Italian communities rich in Dasypyrum villosum (L.) Borbas.
The first cluster analysis (Figure 5) performed to establish the similarity/dissimilarity between the different associations present in the western Mediterranean revealed two groups of associations very distant from each other: group G1 from 1 - 50 and group G2 from 11 to 47, with a distance of 100% between them. Of the 74 community tables, 42 (57.5%) have a distance of 75%, which highlights the substantial differences between the communities in the study.
Taking into consideration the dominant species in each community table, it can be seen in the cluster that the tables rich in Hordeum leporinum Link are distant from those which have a predominance of Chrysanthemum coronarium L. or Dasypyrum villosum (L.) Borbas. The tables rich in Hordeum leporinum Link are distributed with 15 in group G1 and 5 in G2; whereas all the tables with high indices of Chrysanthemum coronarium L. are grouped in G2. When the tables correspond to the same association—for example, 21 and 32—they have a high degree of similarity, with 94.12%. Both the communities with a high frequency of Chrysanthemum coronarium L. and those of Dasypyrum villosum (L.) Borbas have differences of over 75% with regard to those in which Hordeum leporinum Link is the dominant species. The floristic study shows clear differences between the 47 association tables from Spain, Portugal and northern Morocco, with 21 communities of 516 species, and the 27 tables from Italy with 22 communities of 448 species; this highlights a different floristic diversity in grasslands in the Iberian olive groves compared to Italian grasslands.

Figure 4. Soil correlation analysis for the associations in Italy (CA, SD, AB), Spain (TT) and Portugal (PA).
In view of the substantial soil differences in terms of organic matter, nitrogen, phosphorus and potassium, in addition to the floristic and structural differences, we propose for the centre-east of the Mediterranean territories the alliance Securigero securidacae-Dasypyrion villosi nova, an alliance with a Euro-Siberian and sub-Medi- terranean optimum characterised by Centaurea napifolia L., Crepis sancta (L.) Babc., Dasypyrum villosum (L.) Borbas, Bromus erectus Huds., Cerastium ligusticum Viv., Picris hieracioides L., Vulpia ligustica (All.) Link. In this alliance we include the grassland communities from Euro-Siberian and sub-Mediterranean environments which present a high frequency of Dasypyrum villosum (L.) P. Candargy, which have until now been included in the alliance Hordeion leporini Br.-Bl. In Br.-Bl., Gajewski, Wraber & Walas 1936 corr. O. Bolòs 1962. We adopt as type of the new alliance the association Bromo rigidi-Dasypiretum villosi Pignatti 1953.
The communities dominated by Chrysanthemum coronarium L. or by Hordeum leporinum Link have signifi- cant structural, soil and floristic differences. At the soil level the communities of Chrysanthemum coronarium L. present soil parameter values of MO, Nt, P, K, and Mg which are closer to those of the communities of Malven- ion neglectae Gutte 1966 than to those of Hordeion leporinii (Table 2); the frequent presence of species of Malvenion neglectae in the communities of Chrysanthemum coronarium L. and a vegetation structure of Malvenion neglectae [43] , is sufficient reason to propose the new sub-alliance Resedo albae-Chrysanthemenion coronarii all nova which we subordinate to Malvion neglectae (Gutte 1966) status novo.
Separately we applied a Jaccard distance and RA ordination cluster analysis to the westernmost communities (Spain, Portugal and northern Morocco) (association tables 1 - 47) and to the Italian communities (48 - 74) in Table 1. Group EP1 appears in the first case, and includes the associations rich in Chrysanthemum coronarium L. and a group of associations EP2 dominated by Hordeum leporinum Link (Figure 7). In the RA analysis, all the tables of the different associations form a compact group C (tables 1 - 44) with the exception of tables 45, 46

Figure 5. Clustering obtained from 740 rows (plants) × 74 association tables using Jaccard distance.
and 47 corresponding to the associations (45) Schismo calycini-Malvetum trifidae Br.-Bl., Font Quer, G. Br.-Bl., Frey, Jansen & Moor 1936, given by their authors [14] in Catalonia, whereas the associations (46) Euphorbio terracinae-Anacycletum coronati Reyes, Wildpret & León 2001 and (47) Bromo-Hirschfeldietum incanae Ober- dorfer ex Lohmeyer 1975 have been given in Lanzarote by [31] ; this last association was also given in Tenerife by [16] , and is near the compact group of associations. Associations 19 and 47 given in Tenerife and Lanzarote presented a distance of 87.14%, with very little similarity between them; both tables do not therefore correspond to the same association, and have major floristic and biogeographical differences. Table 19, with seven invento- ries given in Tenerife, is rich in Hordeumn leporinum Link and has four inventories without Hirschfeldia incana (L.) Lagr.-Foss.; there is no presence of Hordeum leporinum Link in the 47 given in Lanzarote which present as differentials from the previous Hordeum murinum L. subsp. glaucum (Steud.) Tzvelev, Erodium neuradifolium Delile, Avena fatua L. subsp. meridionales A. W. Hill, Avena sterilis L. subsp. ludoviciana (Durieu) Guillet & Magne and the endemic species Sonchus bourgeaui Sch. Bip. In Webb & Berthel (Figure 6 and Figure 7). W therefore propose a new association for Lanzarote: Soncho bourgeaui-Hirschfeldietum incanae Reyes et al. ex Cano nova, and we propose as type the synthetic inventory in Table 10 published by [31] .
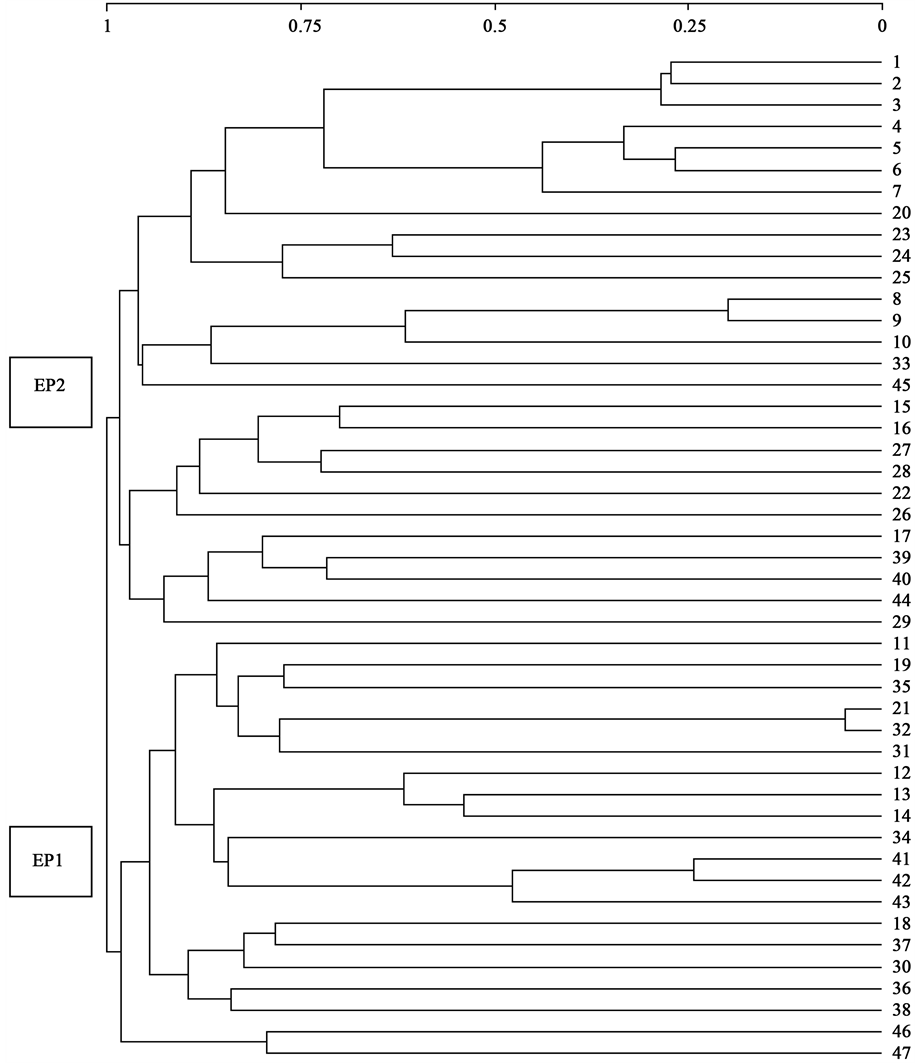
Figure 6. Jaccard distance clustering (Spain-N Morocco-Portugal).

Figure 7. RA ordination analysis applied to the communities present in Spain, Portugal and northern Morocco.
In the case of Italy we applied the same statistical analysis as in the previous case. The cluster separates the tables of the associations belonging to the new alliance Securigero securidacae-Dasypyrion villosi, group M1 (association tables 48 - 74), from group M2 (51 - 63) (Figure 8) belonging to Hordeion leporini and to the new sub-alliance Resedo albae-Chrysanthemenion coronarii, with distances between both groups greater than 75%. On applying the DECORANA ordination analysis, the two groups M1 and M2 are separated. Group M1 in- cludes the association tables 48 AB, 49 CA, 50 SD, 55 DA, 70 BD, 74 BD, 71 VD, 72 LD, 73 AB and 69 RP, but with a subgroup M11 formed by the communities inventoried by us (AB, CA, SD) (Figure 9).
The following cluster (Figure 10) based on the Excel chart with 281 rows (species) × 108 columns (inventories), differentiates various groups. Group G1 (1SD-13oLD) includes 16 of the 18 samples of the association Securigero securidacae-Dasypyretum villosii (1SD-16SD); the two remaining samples (2SD-3SD) are separated from the rest as they have a greater quantity of Hordeum leporinum Link than of Dasypyrum villosum (L.) Borba. Also belonging to group G1 are the inventories of the associations Vulpio ligusticae-Dasypyretum villosi (VD) and Laguro ovati-Dasypyretum villosi (LD), described by [36] , which are together both in the cluster and in the DECORANA ordination analysis. The 11 inventories of Convolvulo elegantissimi-Aegilopetum geniculatae (CA) constitute a very clearly delimited group G2 (19CA-20CA). The following 20 G3 samples (1oBDv-18oDBv) be- long to the association Bromo rigidi-Dasypyretum villosi Pignatti 1953, described for the north-eastern plain of Venice (Italy). The association Aveno barbatae-Brometum diandri belongs to group G4 and comprises two sub- groups: (30AB-43AB), which contains the 16 samples taken by us in grasslands in cultivated olive groves near Spoleto, Trevi, Foligno, Assisi (region of Umbria, Italy). The subgroup (1oAB-12oAB) includes the original samples published by [39] . Finally, a separate group G5 (1oDA-4oDA) contains the inventories corresponding to the association Dasypyro-Aegilopetum triuncialis described by [32] in the Aspromonte.
The DECORANA ordination analysis (Figure 11) gives a first group of inventories (A) which belong to the original associations oAB, oBDv and the inventories taken by us belonging to AB; and a second group (B) con- stituted by the associations VD and LD described by [36] , [38] , which are perfectly separated from the SD and CA communities included in group (C).
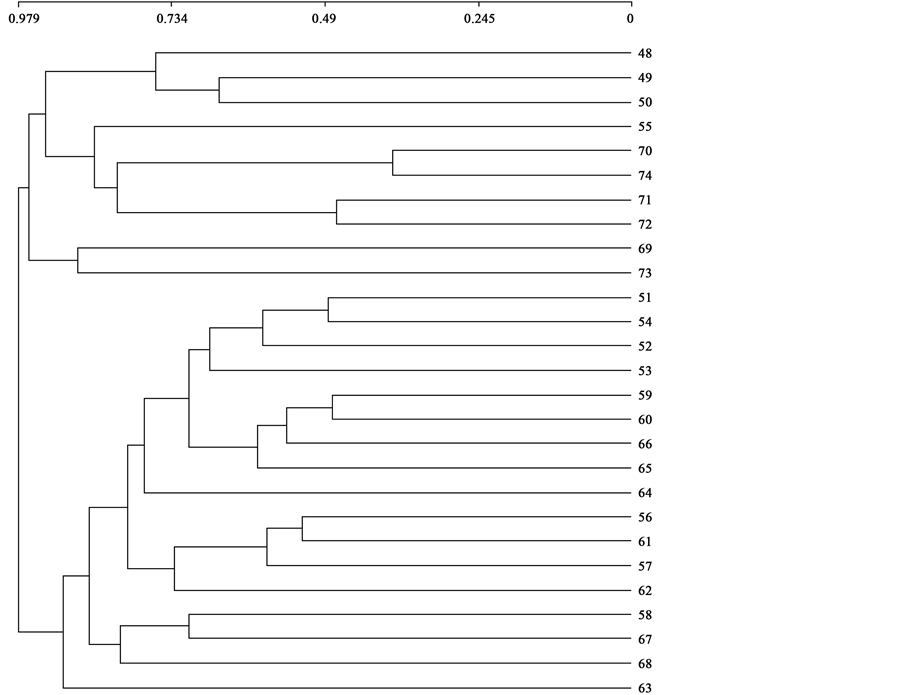
Figure 8. Jaccard distance clustering: Italian communities.

Figure 9. DECORANA ordination analysis: Italian communities.

Figure 10. Euclidean distance cluster analysis for the Italian communities with Dasy- pyrm villosum.
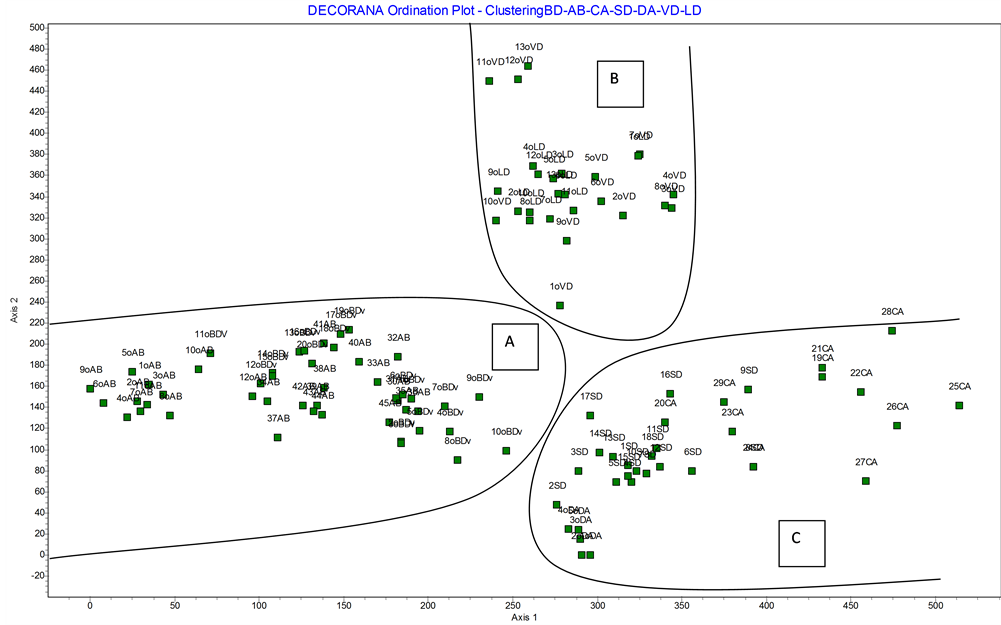
Figure 11. DECORANA ordination analysis for the Italian communities with Dasypyrm villosum.
The formation of two groups of Italian (I1) and Iberian (I2) communities (TT and PA) due to their soil differ- ences, is corroborated by an analysis of conglomerates (Figure 12). The similarly between the CA communi- ties in Italy and PA in Portugal and TT in Spain with regard to their physiognomy and plant structure does not justify their inclusion in the alliance Taeniathero-Aegilopion geniculatae; we thus maintain the CA communi- ties within the new alliance Securigero securidacae-Dasypirion villosi.
As a result of the strong soil, floristic and biogeographical differences, we segregate from the alliance Horde- ion leporini the communities rich in Dasypyrum villosum (L.) Borbas, which we include in the new alliance Se- curigero securidacae-Dasypirion villosi.
The alliance Hordeion leporini Br.-Bl. in Br.-Bl., Gajewski, Wraber et Walas 1936 corr. O. Bolòs 1962, in- cludes grasslands growing in spring with a sub-nitrophilous and nitrophilous character. This alliance has its op- timum in the Mediterranean and radiates to sub-Mediterranean territories. The species Dasypyrum villosum (L.) Borbas grows in the Gargano region in Italy, and is a calcicolous plant from southeast Europe which is absent from Spain and Portugal. A similar situation occurs with Securigera securidaca (L.) Deg. et Dorfl., which is dis- tributed around southern Europe as far as southeast France. Crepis sancta (L.) Babc. appears frequently in the territories sampled in Italy, and is found in south-eastern Europe and the eastern Mediterranean region. This species is common in the region of La Marche, where it has been used to diagnose the vegetation infesting vineyards [40] . All the communities of Dasypyrum villosum (L.) Borbas are grassland formations with dense coverage and substantial biomass which flourish in environments with a high quantity of organic matter and ni- trogen, with a pH always above 8.
As a consequence of the considerable soil, floristic and biogeographical differences, we segregate from the al- liance Hordeion leporine the communities rich in Dasypyrum villosum (L.) Borbas which we include in the new alliance Securigero securidacae-Dasypirion villosi. These are nitrophilous grasslands with a sub-Mediterranean, thermo, meso, supra-Mediterranean and Euro-Siberian (central-European) thermo, meso and sub-temperate character, whose characteristic species are Dasypyrum villosum (L.) Borbas, Securigera securidaca (L.) Deg. et Dorfl., Crepis sancta (L.) Babc. As type association we propose Bromo rigidi-Dasypiretum villosi Pignatti 1953.

Figure 12. Cluster analysis among associations for Italy, Spain and Portugal.
The three types of grasslands studied in Italy belong to three different communities AB, CA, SD, and are per- fectly separated in the cluster and in the PCA (Figure 13 and Figure 14), with the exception of samples 2 and 3 in SD which are separated in the cluster but not in the PCA, due to their greater richness in alliance plants.

Figure 13. Cluster analysis for the three associations studied in Italy.

Figure 14. PCA for the three associations studied in Italy.
Aveno barbatae-Brometum diandri Biondi & Baldoni 1991 has been published by its authors for central Italy [39] , initially located in the regions of Tuscany, Marche, Umbria and Lazio. The sampled community is a grassland with a ruderal character growing on disturbed soils, with therophytic vegetation whose successive evolution leads to perennial herbaceous formations of hemicryptophytes, belonging to the class Agropyretea intermedii- repentis Muller et Görs 1969 and the alliance Convolvulo-Agropyrion repentis Görs 1967—a syntaxon which indicates good soil humidity—whereas in drier areas it contacts with the class Festuco-Brometea Br.-Bl. et Tx. 1943, association Centauro bracteatae-Brometum erecti Biondi et al. 1986. Its soil character indicates that it is located on terrains with pH values of 8.04; C.E.C (meq/100g) 24.685; OOM (%) 6.389; P (p.p.m) 59.04; sandy-silty texture, average CR. This association is dominated by species such as Bromus Diandrus Roth, Bromus madritensis L., Avena barbata Pott ex Link, Hordeum leporinum Link.
The association Convolvulo elegantissimi-Aegilopetum geniculatae Cano-Ortiz, Biondi & Cano nova hoc loco (Table 3 inv. 1-11 typus inv. 8) was inventoried by us in the region of Gargano, a territory located in southeast Italy on the coast of the Adriatic. Precipitation ranges between 686 mm and 1.082 mm , and average annual Tª is between 11.0˚C - 15.3˚C [41] . The dominant materials are basic, with frequent appearance of decarbonated substrates with a neutral-basic character. The association we propose is characterised floristically by Aegilops geniculata Roth and the endemism Convolvulus elegantissimus Miller. This is a grassland which appears relatively frequently in abandoned olive groves, where the organic matter is somewhat inferior to the rest of the nearby communities (Table 2) such as Aveno barbatae-Brometum diandri Biondi et Baldoni 1991 and Securigero se- curidacae-Dasypyretum villosii; however the average annual Tª is higher, and the thermotype is meso- and thermo-Mediterranean with a dry-subhumid ombrotype. The distribution area for this new syntaxon is at least the Gargano. Due to the soil values obtained, its floristic composition and its structure, we include this association Convolvulo elegantissimi-Aegilopetum geniculatae in the new alliance Securigero securidacae-Dasypyrion villosii. The new association is included in the series Cyclamidi hederifolii-Querco ilicis s., Cyclamidi hederifolii- Querco Virginianae s., Fraxino orni-Querco ilicis s. [41] .
Finally the association Seurigero securidacae-Dasypyretum villosii Cano-Ortiz, Biondi & Cano nova hoc loco (Table 4 inv. 1-18 typus inv. 13); this community was inventoried in the region of Gargano, and grows in meso- Mediterranean subhumid environments on basic substrates. It is dominated floristically by Dasypyrum villosum (L.) Borba, Securigera securidaca (L.) Deg. et Dorfl., Echium italicum L., Medicago tenoreana Ser., Medicago disciformis DC., Medicago intertexta (L.) Miller, Bromus ramosus Hudson.
This is a community which grows on soils with a higher content of MOO and nitrogen than Convolvulo ele- gantissimi-Aegilopetum geniculatae, and we include it in the alliance Securigero securidacae-Dasypyrion vil- losii. This new association Securigero securidacae-Dasypiretum villosii (50SD) is statistically separate from the Avenulo barbatae-Brometum diandri Biondi et Baldoni 1991 (48AB), (Figure 9) described for central Italy and which is characterised by Bromus diandrus Roth, Avena barbata Potter, Hordeum leporinum Link, Crepis vesicaria L. subsp. taraxacifolia [39] . It is additionally differentiated from the association Bromo rigidi-Dasypyre- tum villosi Pignatti 1953, also described for the eastern Venetian plain, which is characterised by a dominance of Dasypyrum villosum and Bromus rigidus (L.) Hubbard.
The high frequency of Securigera securidaca (L.) Deg. et Dorfl. in our association, absent in the previous associations, in addition to the bioclimatic and biogeographical differences, are sufficient reason to propose this new association with a sub-Mediterranean character and with a distribution areas which is at least the Gargano region.
4. Conclusions
This study clearly highlights the differentiation of Italian grasslands from Iberian grasslands, which are separated into two alliances: the alliance Hordeion leporini distributed throughout the western Mediterranean, from which we segregate the communities of Chrysanthemum coronarium L., as they present soil, floristic and structural differences which will be included in the new sub-alliance Resedo-Crhysanthemenion coronarii. The analysis of 74 association tables containing 847 inventories and the study of 16 soil parameters in each of the 265 samples provides sufficient information to propose the new alliance Securigero securidacae-Dasypyrion villosi, nitrophilous grasslands with a sub-Mediterranean, thermo, meso, supra-Mediterranean and Euro-Siberian (central-European) thermo, meso and sub-temperate character, whose characteristic species are Dasypyrum villosum (L.) Borba, Securigera securidaca (L.) Deg. et Dorfl., Crepis sancta (L.) Babc. As type association we
Table 3. As. Convolvulo elegantissimi-Aegilopetum geniculatae Cano-Ortiz, Biondi & Cano nova hoc loco.
Sites: All the inventories were taken in the region of Apulia (Gargano), Italy.
Table 4. As. Securigero securidacae-Dasypyretum villosii Cano-Ortiz, Biondi & Cano nova hoc loco.
Sites: All the inventories were taken in the region of Apulia (Gargano), Italy.
propose Bromo rigidi-Dasypyretum villosi Pignatti 1953. In this alliance we include all the associations rich in Dasypyrum villosum (L.) Borbas and the two new associations inventoried by us: Convolvulo elegantissimi-Ae- gilopetum geniculatae and Securigero securidacae-Dasypyretum villosii. We propose the new sub-alliance Resedo albae-Chrysanthemenion coronarii subordinated to the alliance Malvion neglectae, in which we include all the associations rich in Chrysanthemum coronarium L. In order to arrive at these conclusions, it was necessary to take into consideration the value of soil factors as the cause of the abundance of the species, and to establish the values for the different soil parameters which act as soil nutrients for the different grassland communities. We therefore used the synthetic Braun-Blanquet index, adapting it for the first time to the Van der Maarel values; this gives considerable strength to the presence of the species in the community, making it possible to establish the distance between associations. We also propose the new association Soncho bourgeaui-Hirschfeldietum incanae Reyes et al. ex Cano nova, based on the table of [31] .
References
- Brullo, S. (1983) L’Hordeion leporini in Sicilia. Archivio Botanico e Biogeografico Italiano, 58, 55-88.
- Rivas-Martínez, S. (1978) La vegetación de Hordeion leporini en España. Doc. Phytosoc, 9, 377-392.
- Vargas, M.A.A. (1996) Flora y vegetación del valle de Villena (Alicante). Inst. Cultura Juan Gil-Albert. Dip. Porv. Alicante, 261 p.
- Biondi, E., Bagella, S., Cassavechia, S., Pinzi, M. and Vagge, I. (1999) La vegetazione a Dasypyrum villosum (L.) P. Candargy lungo le coste dell’Italia Settentrionale. Documents Phytosociologiques, 19, 439-446.
- Cano-Ortiz, A. (2007) Bioindicadores ecológicos y manejo de cubiertas vegetales como herramienta para la implanta- ción de una agricultura sostenible. Tesis doctoral, Universidad de Jaén, Jaén, 708 p.
- Cano-Ortiz, A., Pinto-Gomes, C.J., Esteban Ruiz, F.J., Torres, A.R., Goñi, J., De la Haza, I. and Cano, E. (2009) Bi- odiversity of Hordeion leporini in Portugal: A phytosociological and edaphic análisis. Acta Botanica Gallica: Botany Letters, 156, 33-48. http://dx.doi.org/10.1080/12538078.2009.10516140
- Cano-Ortiz, A., Pinto-Gomes, C.J., Ruiz, F.J.E. and Cano, E. (2009) Determination of the Nutritional State of Soils by Means of the Phytosociological Method and Different Statistical Techniques (Bayesian Statistics and Decision Trees) in Spain. Acta Botanica Gallica: Botany Letters, 156, 607-624. http://dx.doi.org/10.1080/12538078.2009.10516180
- Cano-Ortiz, A., Pinto-Gomes, C.J. and Cano, E. (2010) Contribution to the Study of the Taenithero-Aegilopion geni- culatae Alliance in Portugal. Acta Botanica Gallica: Botany Letters, 157, 599-610. http://dx.doi.org/10.1080/12538078.2010.10516234
- Cano-Ortiz, A., Del Río González, S. and Gomes, C.J.P. (2013) Impact of Soil Texture on Plant Communities. Plant Sociology, 50, 39-46.
- Costa, C., Capelo, J., Jardin, R., Sequeira, M., Espirito-Santo, D., Lousa, M., Fontinha, S., Aguiar, C. and Rivas-Mar- tínez, S. (2004) Catálogo sintaxonómico e florístico das comunidades vegetais da Madeira e Porto Santo. Quercetea, 6, 61-200.
- Pignatti, S. (1982) Flora d’Italie. Edagricole, Bologna.
- Tutin, et al. (1964-1993) Flora Europea. Vol. I-V, Cambridge University Press, Cambridge.
- Braun-Blanquet, J. (1979) Fitosociología. Bases para el estudio de las comunidades vegetales. Ed. Blume. Madrid.
- Braun-Blanquet, J., Quer, P.F., Braun-Blanquet, G., Frey, E., Jansen, P. and Moor, M. (1936) L’excursion de la sigma en Catalogne (Pâgues 1934). Cavanillesia, 7, 153-167.
- Goday, S.R. (1964) Vegetación y florula de la cuenca extremeña del Guadiana. Excma. Dip. Prov. Badajoz, Madrid, 777 p
- Rivas-Martínez, S., Wildpret, W., Del Arco, M., Rodríguez, O., de Paz, P.P.L., García-Gallo, A., Acebes, J.R., Díaz González, T.E. and Fernández-González, F. (1993) Comunidades vegetales de la Isla de Tenerife (Islas Canarias). Iti- nera Geobotánica, 7, 69-374.
- Roscales, S.S. (2004) Flora y vegetación del macizo occidental de la Sierra de Gredos (Sistema Central, España). Gui- neana, 10, 1-474.
- Fuentes, A.G., Cano, E., Torres, J.A. and Nieto, J. (1994) Notas sobre la vegetación nitrófila de la cuenca del Guadal- quivir. Naturalia Baetica, 6, 125-153.
- Peinado, M., Martínez-Parras, J.M. and Bartolomé, C. (1986) Notas sobre vegetación nitrófila II: Algunas novedades fitosociológicas en Andalucía. Studia Botanica, 5, 53-69.
- Orellana, J.A.V. and de Mera, A.G. (2008) Nuevas aportaciones al conocimiento de la vegetación luso-extremadurense. Estudio de las sierras de las Villuercas (Extremadura, España) y San Mamede (Alto Alentejo, Portugal). Acta Botanica Malacitana, 33, 169-214.
- Maestre, M.A.V. (1997) La sierra de Crevillente: Flora y vegetación. Inst. Cultura Juan Gil-Albert. Dip. Porv. Alicante, 320 p.
- Rivas-Martínez, S., Costa, M. and Loidi, J. (1992) La vegetación de las islas de Ibiza y Formentera (Islas Baleares, España). Itinera Geobotánica, 6, 99-236.
- Fuentes, A.G. and Cano, E. (1993) Aportación al conocimiento de los herbazales nitrófilos hispalenses. Boletín del Instituto de Estudios Giennenses, 148, 275-287.
- Neto, C.S. (2002) A Flora e a Vegetaçao do superdistrito sadense (Portugal). Guineana, 8, 1-269.
- Pereira, M.M.D. (2009) A Flora e Vegetaçao da Serra de Monfurado (Alto Alentejo-Portugal). Guineana, 15, 1-316.
- Bolòs, O., Molinier, R. and Montserrat, P. (1970) Observations phytosociologiques dans l’ile de Minorque. Acta Bo- tanica Barcinon, 5, 1-150.
- Chueca, F.E. (1972) Vegetación y flora de las regiones central y meridional de la provincia de Murcia. Murcia, 451 p
- Magallon, A.R. (1972) Flora y vegetación de la provincia de Alicante. Inst. Est. Alicantinos. Alicante, 403 p.
- de Mera, A.G., Deil, U. and Orellana, J.A.V. (2004) Roadside vegetation in the campo de Gibraltar (sw Spain) and on the Tanger Península (nw Maorocco). Studia Botanica, 23, 63-93.
- Prieto, P., Espinosa, P. and Fábregas, S.F. (1973) Ecología y flora de los tejados de Granada. Trabajos del Departa- mento de Botánica. Universidad de Granada, 2, 97-102.
- Betancort, J.A.R., De La Torre, W.W. and León, M.C. (2001) The Vegetation of Lanzarote (Canary Islands). Phyto- coenologia, 31, 185-247. http://dx.doi.org/10.1127/phyto/31/2001/185
- Brullo, S., Scelsi, F. and Spampinato, G. (2001) La vegetazione dell’ Aspromonte Studio Fitosociologico. Ed. Laruffa. Reggio Calabria.
- Ferro, G. (1980) La vegetazione di Butera (Sicilia meridionale). Attidell’Istituto Botanico e Laboratorio Crittogamico Università di Pavia, 13, 51-118.
- Ferro, G. (2004) Nuovi dati sulla flora e sulla vegetazione dei coltivici e degli incolti di Lipari (Isole Eolie). Quaderni di BotanicaAmbientale e Applicata, 15, 21-39.
- Kruska, K. (1985) Contributo alla conoscenza della vegetazione ruderale delle Marche. Documents Phytosociologiques, 9, 359-368.
- Falleni, G. (1998) Dasypyrum villosum Vegetation in Territory of Rome. Rendiconti Lincei, 9, 149-170. http://dx.doi.org/10.1007/BF02904397
- Biondi, E., Vagge, I., Baldoni, M. and Taffetani, F. (1997) La vegetazione del Parco fluviale del Taro (Emilia-Romag- na). Fitosociologia, 34, 69-110.
- Falleni, G. and Lucchese, F. (1998) The Status of Brometalia rubenti-Tectorum Communities from the Mediterranean Area in Different Syntanomical Schemes. Rendiconti Lincei, 9, 241-255. http://dx.doi.org/10.1007/BF02904407
- Biondi, E. and Baldoni, M. (1991) La vegetazione di margine stradale dell’ordine Brometalia rubenti-tectori nell’Italia Centrale. Annalidi Botanica, 49, 214-217.
- Baldoni, M., Biondi, E. and Loiotile, A (2001) La vegetazione infestante i vigneti nelle Marche. Fitosociologia, 38, 63- 68.
- Falleni, G., Lucchese, F. and Paura, B. (2001) La praterie a Stipa austroitalica du sue settori adriatici meridionali (Mo- lise e Gargano). Fitosociologia, 38, 25-36.
- van der Maarel, E. (1979) Transformation of Cover-Abundance Values in Phytosociology and Its Effects on Commu- nity Similarity. Vegetatio, 39, 97-114. http://dx.doi.org/10.1007/BF00052021
- Gutte, P. (1966) Die Verbreitung einiger Ruderalpflanzengesellschaften in der witeren Umgebung von Leipzig. Wiss.Z. Martin-Luther-Univ. Halle-Wittemberg. Math.-Naturwiss. Reihe 15:937-1010.
Syntaxonomical Checklist
Stellarietea mediae Tüxen, Lohmeyer & Preising ex von Rochow 1951
Chenopodio-Stellarienea Rivas Goday 1956
Sisimbrietalia officinalis J. Tüxen in Lohmeyer & al. 1962 em. Rivas-Martínez, Báscones, T. E. Díaz, Fer- nández-González & Loidi 1991
Hordeion leporini Br.-Bl. in Br.-Bl., Gajewski, Wraber & Walas 1936 corr. O. Bolòs 1962
Bromo scoparii-Hordeetum leporinii Rivas-Martínez 1978
Papaveri rhoeadi-Diplotaxietum virgatae Rivas-Martínez 1978
Anacyclo radiati-Hordeetum leporini O. Bolòs & Rivas-Martínez in Rivas-Martínez 1978
Anacyclo radiati-Papaveretum rhoeadis Cano-Ortiz, Pinto-Gomes, Esteban, Rodríguez-Torres, Goñi, De la Haza & Cano 2009
Anacyclo clavati-Hordeetum leporinii Cano-Ortiz, Pinto-Gomes, Esteban & Cano 2009
Carduo tenuiflori-Hordeetum leporini Br.-Bl. 1931
Bromo-Hirschfeldietum incanae Lohmeyer 1975
Iondrabo auriculatae-Erucetum vesicariae Rivas-Martínez 1978
Rapistro rugosi-Sisymbrietum crassifolii Rivas-Martínez 1978
Hordeo leporini-Glossoppapetum macroti Peinado, Martínez-Parras & Bartolomé 1986
Co. Linaria spartea et Raphanus raphanistrum
Raphano raphanistri-Diplotaxietum catholicae
Asphodelo fistulosi-Hordeetum leporini A. & O. Bolòs in O. Bolòs 1956
Aveno lusitanicae-Hordeetum leporini Espirito Santo, J. C. Costa, Jardin & Sequeira 2004
Bromo madritensis-Galactitetum tomentosae O. Bolòs, Molinier & P. Monserrat 1970
Eruco longirostris-Diplotaxietum erucoidis Rigual 1972 corr. Alcaraz 1984
Sisymbrio irionis-Sinapietum mairei
Schismo calycini-Malvetum trifidae Br.-Bl., Font Quer, G. Br.-Bl., Frey, Jansen & Moor 1936
Euphorbio terracinae-Anacycletum coronati Reyes, Wildpret & León 2001
Soncho bourgeaui-Hirschfeldietum incanae Reyes ex Cano nova hoc loco
Hordeo-Sisymbrietum orientalis Oberd. 1954
Hordeo-Crepidetum bursifoliae Brullo, Scelsi & Spanpinato 2001
Centaureetum napifoliae Brullo 1983
Hypochoerido-Plantaginetum serrariae Brullo 1983
Hordeo-Senecionetum squalidi Brullo 1983
Hordeo-Erodietum acaulis Brullo 1983
Hordeo-Vulpietum ligusticae Brullo 1983
Carduetum australis Brullo 1983
Senecioni cosyrensi-Hordeetum leporini Brullo 1983
Hordeo-Centauretum macracanthae Brullo 1983
Malvion neglectae (Gutte 1966) status novo
Resedo albae-Chrysanthemenion coronarii Cano-Ortiz, Biondi & Cano suball. nova
Resedo albae-Chrysanthemetum coronarii O. Bolòs & Molinier 1958
Anacyclo radiati-Chrysanthemetum coronarii (Rivas-Martínez 1978) Cano-Ortiz, Pinto-Gomes, Esteban, Ro- dríguez-Torres, Goñi, De la Haza & Cano 2009
Malvo parviflorae-Chrysanthemetum coronarii Ferro 1980
Lavatero creticae-Chrysanthemetum coronarii Ferro 2004
Securigero securidacae-Dasypyrion villosi Cano-Ortiz, Biondi & Cano all. nova
Bromo rigidi-Dasypyretum villosi Pignati 1953
Securigero securidacae-Dasypyretum villosi Cano-Ortiz, Biondi & Cano nova hoc loco
Convolvulo elegantissimae-Aegilopetum geniculatae Cano-Ortiz, Biondi & Cano nova hoc loco
Aveno barbatae-Brometum diandri Baldoni & Biondi 1993
Dasypyro-Aegilopetum triuncialis Brullo, Scelsi & Spanpinato 2001
Laguro ovati-Dasypyretum villosi Falleni 1998
Vulpio ligusticae-Dasypyretum villosi Falleni 1998
Rumici-Carduetum pycnocephali Kruska 1985
Artemisietea vulgaris Lohmeyer, Preising & Tüxen ex von Rochow 1951
Carthametalia lanati Brullo in Brullo & Marcenò 1985
Silybo-Urticion Sissingh ex Br.-Bl. & O. Bolòs 1958
Chrysanthemo-Silybetum mariani Bullo 1983