Open Journal of Forestry
Vol.05 No.04(2015), Article ID:55538,8 pages
10.4236/ojf.2015.54036
The Effects of Multilocus Heterozygosity on the Longevity of Seedlings Established on Fallen Logs in Picea jezoensis and Abies sachalinensis
Momoko Okada1, Keiko Kitamura2, Chunlan Lian3, Susumu Goto4
1Kumamoto Branch Office, Japan Finance Corporation, Kumamoto, Japan
2Hokkaido Research Center, Forestry and Forest Products Research Institute, Sapporo, Japan
3Asian Natural Environmental Science Center, The University of Tokyo, Tokyo, Japan
4Education and Research Center, The University of Tokyo Forests, Graduate School of Agricultural and Life Sciences, The University of Tokyo, Tokyo, Japan
Email: gotos@uf.u-tokyo.ac.jp
Copyright © 2015 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 10 March 2015; accepted 10 April 2015; published 13 April 2015
ABSTRACT
Seedling establishment on fallen logs is a major regeneration system for tree species in boreal forests. Seedling survival on fallen logs is affected not only by the microsite environment but also by the genetic factors of individuals. To quantify the genetic effects on seedling longevity, we identified seedlings using a number tag system and collected needles of Picea jezoensis and Abies sachalinensis established on fallen logs in spring 2006. Survival or death of each seedling was investigated during 2006-2012. We genotyped seedlings with microsatellite markers and calculated individual-based multilocus heterozygosity (MLH) for each seedling. A Cox proportional hazards model was applied to evaluate the effects of MLH on seedling longevity of the two species considering the fallen log conditions. The model indicated that MLH positively affected seedling longevity in P. jezoensis, whereas the effects of MLH were not significant in A. sachalinensis. Here, we discuss differences in the effects of MLH on seedling longevity between the two species, considering species characteristics and MLH frequency distribution.
Keywords:
Cox Proportional Hazards Model, Fallen Log, Inbreeding Depression, Microsatellite, Survival Census, Tree Regeneration 
1. Introduction
Tree regeneration on fallen logs is common in boreal forests. Fallen logs provide safe sites for tree seedlings because competition with understory herbs is greatly reduced (Harmon & Franklin, 1989) and few pathogenic fungi can infect tree seedlings on fallen logs (O’Hanlon-Manners & Kotanen, 2004) . Well-decomposed fallen logs with moss cover show reduced evaporation and contain water-soluble nutrients (Takahashi et al., 2000) . However, seedling densities differ markedly among fallen logs, and not all fallen logs provide safe sites (Takahashi et al., 2000; Narukawa et al., 2003; Iijima & Shibuya, 2010) . In addition to the fallen log conditions, genetic factors of each seedling may also impact tree recruitment success. Experimental studies have revealed that selfed and/or inbred progeny suffer from low viability and survivorship (Husband & Schemske, 1996; Kadomatsu et al., 1997; Hirao, 2010) . Parentage analyses of naturally established seedlings have revealed that the proportion of selfed individuals decreases from the seedling stage to the sapling stage (Naito et al., 2005; Isagi et al., 2007) . However, the relative importance of genetic effects and site conditions on seedling longevity has not been evaluated in a natural forest.
Due to their highly polymorphic, co-dominant nature and neutrality, microsatellite markers are useful for detecting heterozygosity-fitness correlations (HFCs) (Hansson et al., 1996; Keller & Waller, 1996; Chapman et al., 2009; Forstmeier et al., 2012) . Previous animal studies have demonstrated that heterozygous individuals grow, survive, and reproduce better than homozygous individuals (Chapman et al., 2009) . Hansson et al. (1996) indicated that individual-based multilocus heterozygosity (MLH) was higher in recruited great reed warblers (Acrocephalus arundinaceus) than in their non-recruited siblings. In contrast to animals, HFCs have rarely been examined in plants. As an exception, Collevatti and Hay (2011) found that MLH of survived individuals at 23 years was significantly higher than that of dead individuals in the neotropical tree, Caryocar brasiliense, based on 10 microsatellite marker genotypes. However, the effects of MLH on seedling survivorship patterns were not described in their study.
Picea jezoensis Carr. (Yezo spruce) and Abies sachalinensis Masters (Todo fir) are two major coniferous species in the mixed forests of Hokkaido, northern Japan (Takahashi et al., 2000; Narukawa et al., 2003) . Studies have revealed that survival of P. jezoensis and A. sachalinensis seedlings is affected significantly by fallen log conditions, which include the hardness of the log surface (Narukawa et al., 2003; Iijima et al., 2007) , moss coverage and/or moss height (Takahashi et al., 2000; Iijima et al., 2007; Iijima & Shibuya, 2010) , and light conditions (Iijima & Shibuya, 2010) . A microsatellite analysis revealed that P. jezoensis predominantly outcrosses (Tomita et al., 2008) . An artificial crossing experiment with A. sachalinensis indicated that selfed progenies suffered from low height growth and survival at the early demographic stage (Kadomatsu et al., 1997) . Thus, these two species have a common mating system and show inbreeding depression similar to other conifers (Husband & Schemske, 1996) . However, the effects of genetic factors on seedling survival of these species have not been investigated.
In the present study, we evaluated the effects of MLH on the longevity of P. jezoensis and A. sachalinensis seedlings established on fallen logs, simultaneously considering the establishment site conditions. Subsequently, we discussed the importance of genetic and environmental factors for seedling longevity of each species.
2. Materials and Methods
2.1. Study Site
The study site was located in the Iwanazawa Forest Reserve, University of Tokyo Hokkaido Forest (UTHF), the University of Tokyo, central Hokkaido, Japan (43˚13'N, 142˚34'E). We established a 200 × 250 m (5 ha) study plot (Figure 1). We measured the diameter at breast height (d.b.h.) of all living trees within the 5 ha plot between 2003 and 2005. We identified 123 P. jezoensis and 170 A. sachalinensis mature trees (d.b.h. > 20 cm). The locations of the trees were identified using a digital compass (LaserAce 300, TimberTech), and their needle leaf tissues were collected for DNA analysis.
We surveyed the forest floor of a 4.29-ha study site located east of the stream in May 2006 and identified 112 fallen logs whose diameters at the bottom end were >20 cm (Figure 1). We defined individuals with <30 cm height on fallen logs as seedlings. Seedlings of both species established on 108 of 112 fallen logs were assigned a number tag, and ages were determined using a bud scale vestige in 2006. At that time, the heights of 211 and

Figure 1. The locations of fallen logs, seedlings, and saplings within the study plot: a) Picea jezoensis and b) Abies sachalinensis. The gray solid line indicates a creek. Large represent the locations of mature trees. Small crosses on the fallen logs represent seedlings. The width of the fallen logs corresponds to the log diameter.
269 seedlings of P. jezoensis and A. sachalinensis were measured, respectively (Table 1). The location of the seedlings was determined using a digital compass (Laser Ace 300, Timber Tech; Figure 1). After identifying the seedlings, we monitored their survival or death in the fall of 2006, 2007, 2009, 2010, 2011, and 2012. Seedling longevity was calculated by adding the age in 2006 to the survived years during the investigation period. If the seedlings were alive in the fall of 2012 (the last census), the longevity of the seedlings was treated as censoring status one for the statistical analysis. We collected needle tissues from half of all P. jezoensis seedlings and stored them in plastic bags with silica gel under dry condition at room temperature. Needle tissues of A. sachalinensis were previously collected by Lian et al. (2008) .
2.2. Fallen log Conditions
Fallen log conditions, such as hardness of the fallen log surface, moss coverage, dwarf bamboo coverage on fallen logs, and moss height of 108 fallen logs, were measured in the fall of 2009. Hardness was measured at five randomly selected sites within each fallen log using a Yamanaka-type soil penetrometer (No. 48915, Takemura Electric Works). Hardness was quantified as the penetration resistance value (kg/cm2). The moss coverage (%) and dwarf bamboo shadow (%), which is a proportion of the surface area of fallen logs covered by dwarf bamboo, were visually assessed in 5% increments in five randomly selected quadrates. Moss height was also measured at these sites for each fallen log. The averages of these values were used as the site condition variables for each fallen log.
2.3. Microsatellite Analysis
DNA was extracted from the dried needles of 211 seedlings and 123 mature trees of P. jezoensis using the DNeasy Plant Mini Kit (Qiagen). Eleven microsatellite loci were used for analyzing P. jezoensis: SpAGC1, SpAGD1, and SpAGG3, developed by Pfeiffer et al. (1997) ; PaGB3 and PgGB5, developed by Besnard et al. (2003) ; EATC1E03 and EATC2G05, developed by Scotti et al. (2002) ; and four primer pairs (pj04, pj08, pj22, and pj24), developed in the present study (Table 2).
Amplification by Polymerase chain reaction (PCR) was conducted using the Multiplex PCR Kit (Qiagen) and the GeneAmp PCR System 9700 (Applied Biosystems) following the manufacturers’ protocols. The size of the PCR products was determined using the ABI 3130 Genetic Analyzer and GeneMapper Software ver. 4.0 (Applied Biosystems).
We used published data sets to genotype 170 mature A. sachalinensis trees and 269 seedlings with seven microsatellite markers (Lian et al., 2008) .

Table 1. Number of seedlings (n) and mean initial height with standard deviation (std) in parenthesis for each age.
Table 2. Microsatellite primer pairs developed for Picea jezoensis in this study.
2.4. Statistical Analysis
MLH, which we calculated as the number of heterozygous loci divided by the total number of loci, was used as a simple measurement of individual heterozygosity (Hansson et al., 1996; Collevatti & Hay, 2011; Forstmeier et al., 2012) . We calculated MLH for 211 seedlings and 123 mature trees of P. jezoensis and for 269 seedlings and 170 mature trees of A. sachalinensis. We analyzed the effects of MLH and site conditions on seedling longevity using the Cox proportional hazards model for each species. Here the dependent variable is the hazard function, which describes how the hazard (e.g., risk or seedling mortality) changes over time, and the effect parameter describes how the hazard relates to different explanatory variables. We used the following model:
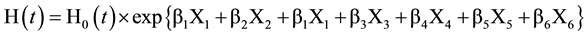
where, H(t) denotes the hazard function at time t, H0(t) corresponds to the baseline hazard, and β is the expected change in the log-relative hazard for a unit change in initial height (X1), MLH (X2), moss coverage (X3), fallen log hardness (X4), moss height (X5), and dwarf bamboo shadow (X6). We used the full model derived from six dependent variables using the coxph function in the R survival package (R Development Core Team, 2012) . Subsequently, we selected variables for the best model based on AIC criterion.
3. Results
3.1. MLH Frequency Distribution
The heterozygosity frequency distribution of seedlings (≤2 years old) was different between P. jezoensis and A. sachalinensis (Figure 2). P. jezoensis exhibited a normal distribution peaking at 0.5 - 0.6, whereas A. sachalinensis displayed a sigmoid distribution peaking at 0.8 - 0.9. Individuals with low heterozygosity (MLH ≤ 0.4) decreased from ≤2-year-old seedlings to ≥3-year-old seedlings in both species. MLH frequency distributions for mature trees were similar to those for seedlings (≥3 years old) for both species. However, the proportion of individuals with low heterozygosity (MLH ≤ 0.4) decreased from seedlings (≥3 years old) to mature trees.
3.2. Seedling Survival of P. jezoensis and A. sachalinensis
Cox proportional hazards regression analyses of seedling longevity were conducted using six variables for P. jezoensis and A. sachalinensis. Initial height, MLH, log hardness, and moss coverage were selected as variables for the best model P. jezoensis, whereas initial height, log hardness, and moss height were selected in A. sacha-

Figure 2. The frequency distribution of multilocus heterozygosity (MLH) of ≤2-year-old seedlings, ≥3-year-old seedlings, and mature trees in P. jezoensis and A. sachalinensis.
linensis; however, MLH was not selected in A. sachalinensis (Table 3). Subsequently, Cox proportional hazards regression analyses using only the selected variables was conducted. The temporal patterns of seedling survival estimated by the selected model are shown in Figure 3. Confidence intervals (CIs) were larger after 10 years due to the limited data. Patterns of seedling survivorship for the two species were similar, but the temporal risk was higher in P. jezoensis for the same year. The estimated survival rate at 5 years was 0.492 (95% CI, 0.423 - 0.572) in P. jezoensis and 0.671 (95% CI, 0.615 - 0.731) in A. sachalinensis. The estimated survival rate at 10 years was 0.158 (95% CI, 0.108 - 0.231) in P. jezoensis and 0.259 (95% CI, 0.204 - 0.327) in A. sachalinensis.
3.3. Effects of Initial Height, MLH, and Site Conditions on Seedling Longevity
Initial height markedly affected seedling longevity of both species (Table 3). The estimated survival rates at 5 years for short (5 cm) and tall (15 cm) P. jezoensis seedlings were 0.586 and 0.944, respectively; those for short and tall A. sachalinensis seedlings were 0.668 and 0.932, respectively. MLH also largely affected P. jezoensis seedling longevity and their estimated survival rates after 5 years for low (0.2) and high (0.8) MLH were 0.344 and 0.568, respectively. In contrast, the effects of log hardness on A. sachalinensis seedling longevity were small. The estimated survival rates at 5 years for soft fallen logs (5 kg/cm2) and hard fallen logs (15 kg/cm2) were 0.627 and 0.721, respectively (Figure 4).
4. Discussion
MLH frequency distributions were compared among ≤2-year-old seedlings, ≥3-year-old seedlings, and mature trees (Figure 2). For both species, MLH distributions of ≥3-year-old seedlings and mature trees shifted higher

Figure 3. The frequency distribution of multilocus heterozygosity (MLH) of ≤2-year-old seedlings, ≥3-year-old seedlings, and mature trees in P. jezoensis and A. sachalinensis. Solid lines indicate the estimated survival functions for the Cox regression of time to seedling death. Dashed lines represent the 95% confidence intervals.
Table 3. Cox proportional harzards regression analyses of Picea jezoensis and Abies sachalinensis seedling survival. Because the dependent variable in the model is a hazard rate, negative β coefficients indicate that a factor has a positive effect (1-βexp) on seedling survival.
ns, not significant; *, p < 0.05; **, p < 0.01; ***, p < 0.001.

Figure 4. Estimated seedling survival functions for the Cox regression of time to seedling death for two situations of each variable: (a) Initial height of Picea jezoensis, short (5 cm) and tall (15 cm); (b) Initial height of Abies sachalinensis, short (5 cm) and tall (15 cm); (c) Multilocus heterozygosity (MLH) of P. jezoensis, low (0.2) and high (0.8); (d) Log hardness for A. sachalinensis, soft log surface (5 kg/m2) and hard log surface (15 kg/m2).
than those of ≤2-year-old seedlings. These shifts were interpreted as purging seedlings with low MLH at the early demographic stage. These trends are consistent with previous studies related to inbreeding depression (Keller & Waller, 1996; Husband & Schemske, 1996) . At the population level, FIS often decreases from the seedling stage to the mature tree stage (Hufford & Hamrick, 2003; Lian et al., 2008) . For example, selfed or inbred progenies tend to be eliminated during the early demographic stages (Karkkainen et al., 1996; Kadomatsu et al., 1997; Isagi et al., 2007; Naito et al., 2005) . Selection of inbred individuals eventually affects genetic variation within the population as FIS often decreases from the seedling stage to the mature tree stage (Hufford & Hamrick, 2003; Lian et al., 2008) .
Statistical modeling enabled us to evaluate the effects of initial height, MLH, and site conditions on seedling longevity for the two species. Initial height was selected as a significantly positive variable for both species (Table 3). This trend is reasonable because tall seedlings tend to survive longer than the short seedlings (Table 1). However, the effects of initial height were slightly larger in P. jezoensis than in A. sachalinensis (Table 1). This difference was caused by a difference in seed and seedling size. Seeds of P. jezoensis are small (approximately 2.4 mg), whereas those of A. sachalinensis are relatively large (approximately 9.8 mg) (ASakawa et al., 1981) . The mean heights of 1-year-old seedlings of P. jezoensis and A. sachalinensis were 1.25 cm and 2.42 cm, respectively (Table 1). Therefore, initial height may affect longevity more in P. jezoensis than in A. sachalinensis.
MLH, not fallen log conditions, significantly affected P. jezoensis seedling longevity (Table 2). The model indicated that relatively heterozygous individuals lived longer than homozygous individuals (Figure 4(c)). This finding was consistent with previous studies in animals and birds (Hansson et al., 1996; Chapman et al., 2009; Hoffman et al., 2014) and with a study in plants (Collevatti & Hay, 2011) . However, offspring survivorship was not quantitatively estimated in those studies. As per our knowledge, the present study is the first to quantify the effects of MLH on seedling longevity in forest trees.
Figure 4(c) suggests that seedlings with high MLH (≥0.8) survived a long time. The survival rates of seedlings at 10 years with MLH = 0.2 and MLH = 0.8 were 6.2% and 22.9%, respectively. If this survivorship pattern continued to the mature tree stage, MLH of most mature trees must be >0.8. Nevertheless, MLH frequency distribution for mature trees was similar to that for seedlings (≥3 years old) (Figure 2). This discrepancy suggests that extending our model to the mature tree stage would be difficult. Our model is constructed with data from established seedlings on fallen logs. Therefore, the initial recruitment process on a fallen log is described by this model. However, the processes from seedlings to juveniles and mature trees must be influenced by other factors, such as height of the fallen log surface from the ground.
No effect of MLH on seedling survival was detected for A. sachalinensis, although this species shows severe inbreeding depression (Kadomatsu et al., 1997) . One possible explanation is that inbreeding depression is negligible in this population. Ubukata et al. (2000) conducted a crossing experiment with A. sachalinensis and found that selfed progenies suffer from low height growth. However, they detected three progenies derived from two populations in a 10-year-old stand that did not exhibit significant inbreeding depression for height growth. Karkkainen et al. (1996) also indicated that the extent of inbreeding depression varies among populations of Pinus sylvestris.
Another possible explanation is that inbreeding depression occurred, but the effects of MLH could not be detected in this study due to marker characteristics. Although MLH frequency distribution in P. jezoensis was normal with a peak of 0.5 - 0.6, MLH frequency distribution in A. sachalinensis was biased with a peak of 0.8 - 0.9. Figure 2 shows that homozygous young A. sachalinensis seedlings seemed to be eliminated. Hoffman et al. (2014) suggested that MLH calculated on the basis of approximately 10 microsatellite markers represents a tiny fraction of the genome and that high-throughput DNA sequencing enables to detect an accurate relationship between MLH and fitness. Thus, further analysis of other populations or other marker systems should be examined to clarify which explanation is true.
5. Conclusion
We evaluated the effects of MLH on P. jezoensis and A. sachalinensis seedling longevity, considering initial seedling height and fallen log conditions. The Cox proportional hazards model enabled us to evaluate the effects of initial height, MLH, and fallen log conditions on P. jezoensis and A. sachalinensis seedling longevity. The model indicated that initial seedling height positively affected seedling longevity in both species, i.e., tall seedlings survived longer than short seedlings. We also found that MLH largely affected P. jezoensis seedling longevity, whereas it did not affect A. sachalinensis seedling longevity. The effects of fallen log conditions detected in A. sachalinensis were relatively small compared with the effects of initial height in the two species and the effects of MLH in P. jezoensis.
Acknowledgements
We thank Y. Takahashi, M. Kimura, N. Miyashita, M. Nakagawa, A. Kurahashi, and the technical staff at the University of Tokyo Hokkaido Forest, the University of Tokyo, for their help in the field and M. Goto for laboratory experiments. We also thank H. Iwata, M. Mimura, and T. Hogetsu for suggestions and N. Miura for editing a draft of this manuscript.
References
- Asakawa, S., Katsuta, M., & Yokoyama, T. (1981). Seeds of Woody Plants in Japan: Gymnospermae (pp. 150). Tokyo: Forest Tree Breeding Association. (In Japanese)
- Besnard, G., Acheré, V., Besnard, G., Faivre, V., Rampant, P., Favre, J. M., & Jendroz, S. (2003). A Set of Cross-Species Amplifying Microsatellite Markers Developed from DNA Sequence Databanks in Picea (Pinaceae). Molecular Ecology, 3, 380-383. http://dx.doi.org/10.1046/j.1471-8286.2003.00456.x
- Chapman, J. R., Nakagawa, S., Coltman, D. W., Slate, J., & Sheldon, B. C. (2009). A Quantitative Review of Heterozygosity-Fitness Correlations in Animal Populations. Molecular Ecology, 18, 2746-2765. http://dx.doi.org/10.1111/j.1365-294X.2009.04247.x
- Collevatti, R. G., & Hay, D. (2011). Kin Structure and Genotype-Dependent Mortality: A Study Using the Neotropical Tree Caryocar brasiliense. Journal of Ecology, 99, 757-763. http://dx.doi.org/10.1111/j.1365-2745.2011.01796.x
- Forstmeier, W., Schielzeth, H., Mueller, J. C., Ellegren, H., & Kempenaers, B. (2012). Heterozygosity-Fitness Correlations in Zebra Finches: Microsatellite Markers Can Be Better than Their Reputation. Molecular Ecology, 21, 3237-3249. http://dx.doi.org/10.1111/j.1365-294X.2012.05593.x
- Hansson, B., Bensch, S., Hasselquist, D., & Akesson, M. (1996). Microsatellite Diversity Predicts Recruitment of Sibling Great Reed Warblers. Proceedings of the Royal Society of London Series B-Biological Sciences, 268, 1287-1291. http://dx.doi.org/10.1098/rspb.2001.1640
- Harmon, M. E., & Franklin, J. F. (1989). Tree Seedlings on Logs in Picae-Tsuga Forests of Oregon and Washington. Ecology, 70, 48-59. http://dx.doi.org/10.2307/1938411
- Hirao, A. S. (2010). Kinship between Parents Reduces Offspring Fitness in a Natural Population of Rhododendron brachycarpum. Annals of Botany, 105, 637-646. http://dx.doi.org/10.1093/aob/mcq018
- Hoffman, J. I., Simpson, F., David, P., Rijks, J. M., Kuiken, T., Thorne, M. A., Lacy, R. C., & Dasmahapatra, K. K. (2014). High-Throughput Sequencing Reveals Inbreeding Depression in a Natural Population. Proceedings of the National Academy of Sciences, 111, 3775-3780. http://dx.doi.org/10.1073/pnas.1318945111
- Hufford, K., & Hamrick, J. (2003). Viability Selection at Three Early Life Stages of the Tropical Tree, Platypodium elegans (Fabaceae, Papilionoideae). Evolution, 57, 518-526. http://dx.doi.org/10.1111/j.0014-3820.2003.tb01543.x
- Husband, B. C., & Schemske, D. W. (1996). Evolution of the Magnitude and Timing of Inbreeding Depression in Plants. Evolution, 50, 54-70. http://dx.doi.org/10.2307/2410780
- Iijima, H., Shibuya, M., & Saito, H. (2007). Effects of Surface and Light Conditions of Fallen Logs on the Emergence and Survival of Coniferous Seedlings and Saplings. Journal of Forest Research, 12, 262-269. http://dx.doi.org/10.1007/s10310-007-0012-1
- Iijima, H., & Shibuya, M. (2010). Evaluation of Suitable Conditions for Natural Regeneration of Picea jezoensis on Fallen Logs. Journal of Forest Research, 15, 46-54. http://dx.doi.org/10.1007/s10310-009-0133-9
- Isagi, Y., Saito, D., Kawaguchi, H., Tateno, R., & Watanabe, S. (2007). Effective Pollen Dispersal Is Enhanced by the Genetic Structure of an Aesculus turbinata Population. Journal of Ecology, 95, 983-990. http://dx.doi.org/10.1111/j.1365-2745.2007.01272.x
- Kadomatsu, M., Kudo, H., Noda, M., Natsume, S., & Kanno, T. (1997). Height Growth of Selfed-Pollinated Abies sachalinensis Seedlings over a Fourteen-Year Period. Journal of Japanese Forest Society, 79, 119-122. (In Japanese with English Summary)
- Karkkainen, K., Koski, V., & Savolainen, O. (1996). Geographical Variation in the Inbreeding Depression of Scots Pine. Evolution, 50, 111-119. http://dx.doi.org/10.2307/2410785
- Keller, L. F., & Waller, D. (1996). Inbreeding Effects in Wild Population. Trends in Ecology and Evolution, 17, 230-241. http://dx.doi.org/10.1016/S0169-5347(02)02489-8
- Lian, C., Goto, S., Kubo, T., Takahashi, Y., Nakagawa, M., & Hogetsu, T. (2008). Nuclear and Chloroplast Microsatellite Analysis of Abies sachalinensis Regeneration on Fallen Logs in a Subboreal Forest in Hokkaido, Japan. Molecular Ecology, 17, 2948-2962. http://dx.doi.org/10.1111/j.1365-294X.2008.03802.x
- Naito, Y., Konuma, A., Iwata, H., Suyama, Y., Seiwa, K., Okuda, T., Lee, S. L., Muhammand, N., & Tsumura, Y. (2005). Selfing and Inbreeding Depression in Seeds and Seedlings of Neobalanocarpus heimii (Dipterocarpaceae). Journal of Plant Research, 118, 423-430. http://dx.doi.org/10.1007/s10265-005-0245-z
- Narukawa, Y., Iida, S., Tanouchi, H., Abe, S., & Yamamoto, S. I. (2003). State of Fallen Logs and the Occurrence of Conifer Seedlings and Saplings in Boreal and Subalpine Old-Growth Forests in Japan. Ecological Research, 18, 267-277. http://dx.doi.org/10.1046/j.1440-1703.2003.00553.x
- O’Hanlon-Manners, D. L., & Kotanen, P. M. (2004). Logs as refuges from fungal pathogens for seeds of eastern hemlock (Tsuga candadensis). Ecology, 85, 284-289. http://dx.doi.org/10.1890/03-3030
- Pfeiffer, A., Olivieri, A. M., & Morgante, M. (1997). Identification and Characterization of Microsatellites in Norway Spruce (Picea abies K.). Genome, 40, 411-419. http://dx.doi.org/10.1139/g97-055
- R Development Core Team (2012). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing http://www.R-project.org/
- Scotti, I., Magni, F., Paglia, G., & Morgante, M. (2002). Trinucleotide Microsatellites in Norway Spruce (Picea abies): Their Features and the Development of Molecular Markers. Theoretical and Applied Genetics, 106, 40-50.
- Takahashi, M., Sakai, Y., Ootomo, R., & Shiozaki, M. (2000). Establishment of Tree Seedlings and Water-Soluble Nutrients in Coarse Woody Debris in an Old-Growth Picea-Abies Forest in Hokkaido, Northern Japan. Canadian Journal of Forest Research, 30, 1148-1155.
- Tomita, M., Suyama, Y., & Saito, H. (2008). Effect of Local Stand Density on Reproductive Processes of the Sub-Boreal Conifer Picea jezoensis Carr. (Pinaceae). Forest Ecology and Management, 256, 1350-1355. http://dx.doi.org/10.1016/j.foreco.2008.06.037
- Ubukata, M., Koono, K. & Itahana, N. (2000). Estimation of Genetic Parameters on Height Growth Using Artificial Mating Families between Plus Trees of Abies sachalinensis. Bulletin of Forest Tree Breeding Center, 17, 135-151. (In Japanese with English Summary)