Journal of Biomedical Science and Engineering
Vol.7 No.9(2014), Article
ID:47570,10
pages
DOI:10.4236/jbise.2014.79062
The Effects of Adiponectin on Bone Metabolism
Yuan Yu Lin1, Ching Yi Chen1, Chih Chien Chen1, Han Jen Lin1, Harry John Mersmann1, Shinn Chih Wu1, Shih Torng Ding1,2*
1Department of Animal Science and Technology, National Taiwan University, Taiwan
2Institute of Biotechnology, National Taiwan University, Taiwan
Email: *sding@ntu.edu.tw
Copyright © 2014 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 27 April 2014; revised 10 June 2014; accepted 25 June 2014
ABSTRACT
Osteoporosis and its related bone fractures are growing medical problems, especially in industrial countries, and thus the knowledge of regulation of bone metabolism is critical to develop therapeutic approaches. Bone adipocytes share common mesenchymal precursors with osteoblasts and chondrocytes and their numbers in bone marrow are altered in various pathophysiological conditions. Several findings suggest that accelerated adipogenesis in bone marrow, known as fatty marrow, is associated with the progression of osteoporosis. Apart from its demonstrated antiatherosclerogenic and insulin-sensitizing actions, the adipokine adiponectin and its receptors have been shown to be expressed in bone tissues and participate in bone metabolism. Here we review recent findings regarding the regulation of bone metabolism by adiponectin and its receptors and the underlying mechanisms. We also provide future perspectives for research.
Keywords:Adiponectin, Mesenchymal Stem Cells, Bone and Osteoporosis

1. Introduction
The relationship between bone and fat formation within the bone marrow microenvironment is complex and remains an area of active investigation. Clinical and experimental findings suggest that acceleration of adipogenesis in bone marrow, known as fatty marrow, is associated with the progression of osteoporosis and aging [1] [2] . It is now clear that adipose tissue is a complex, essential and highly active metabolic and endocrine organ [3] [4] . Adipocytes express and secrete various endocrine hormones such as leptin, adiponectin (ApN), TNF-α and IL-6 [4] . Among these hormones, leptin is the first adipokine found to have function in both energy and bone metabolism [5] [6] . These observations suggest that energy metabolism and bone mass are regulated by the same hormones. Over the past decades, ApN and its receptors (AdipoR1 and AdipoR2) have been found to regulate energy homeostasis and have protective functions for metabolic and cardiovascular diseases [7] . In addition, accumulating evidences indicate that ApN measurement may serve as a useful screening tool for predicting osteoporosis [8] . ApN and its receptors are expressed in cells of osteoblastic and osteoclastic lineages, suggesting that they play roles in regulating bone metabolism [9] [10] . Here, we summarize the effects of ApN on bone metabolism and propose future research directions.
2. ApN and Its Receptors
ApN was originally identified by four independent groups and was named Acrp30 [11] and AdipoQ [12] in mice, and apM1 [13] or GBP28 [14] in humans. It belongs to the complement 1q family [15] and forms multimer complexes through the collagen-like domain in the circulatory system. Multi-mer complexes include trimers, hexamers and high-molecular-weight (HMW) forms [16] . ApN-deficient mice exhibit features of insulin resistance, dyslipidemia and hypertension [17] . ApN exerts its insulin-sensitizing effect by regulating glucose utilization and fatty acid metabolism [7] . Administration of ApN to mice decreases the plasma concentration of glucose, free fatty acids and triglycerides, increases muscular fatty acid oxidation, induces weight loss [18] and reverses obesity-associated insulin resistance [19] .
Yamauchi et al. first cloned the cDNA encoding adiponectin receptors 1 (AdipoR1) and 2 (AdipoR2) from humans and mice [20] . Our group, then cloned the porcine counterparts [21] . AdipoR1 and AdipoR2 contain seven transmembrane domains with structure, topology and function distinct from those of the G-protein-coupled receptors [20] . AdipoR1 and AdipoR2 serve as the major AdipoRs in vivo, with the former activating the AMP kinase (AMPK) pathway and the latter activating the peroxisome proliferator-activated receptor alpha (PPARα) pathway in liver to enhance insulin sensitivity and decrease inflammation [7] . Disruption of both AdipoR1 and R2 exterminate ApN binding and downstream actions including abolishment of ApN-induced AMPK activation and decreased activity of the PPAR-α signaling pathway; The result of disruption is to increase tissue triglyceride content, inflammation, oxidative stress, insulin resistance and glucose intolerance [22] . Recent research indicates ApN has anti-proliferative effects on cancer cells and a cardio-protective role increasing longevity [23] . Figure 1 represents the effects of adiponectin on peripheral tissues and hypoadiponectinemia related diseases.
3. Bone Remodeling
Bone remodeling is a continuous process throughout adult life consisting of the resorption of senescent bone
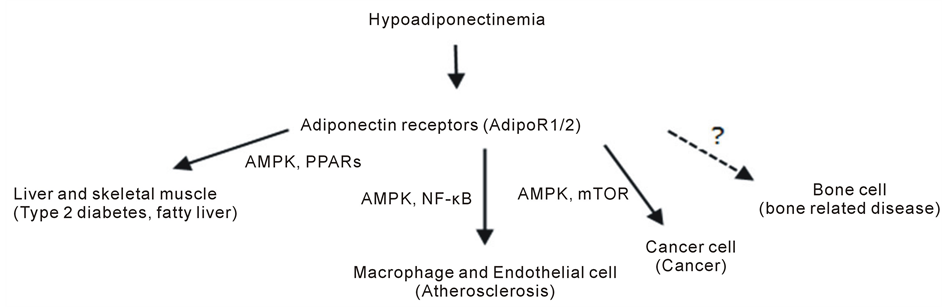
Figure 1. Current targets and mediators of adiponectin and its receptors. Adiponectin and its receptors (AdipoR1/R2) play critical roles in the regulation of peripheral tissue functions and development of obesity-related disease, such as type 2 diabetes, fatty liver, atherosclerosis, bone related disease or cancer. AdipoR1/2 serve as receptors for adiponectin actions, which are mediated by AMPK, PPARs, NF-κB and mTOR. The related diseases are indicated in parentheses. (AMPK, 5’ adenosine monophosphate-activated protein kinase; PPARs, peroxisome proliferative activated receptors; NF-κB, nuclear transcription factor κB; mTOR, mammalian target of rapamycin; modified from [23] ).
by osteoclasts (Oc) and the formation of new bone from osteoblasts (Ob). Differentiated Oc are derived from hematopoietic stem cells [24] , whereas Ob are derived from mesenchymal stem cells [25] . In the process of bone remodeling, Oc adhere to bone to remove the exterior rigid structure by acidification and proteolytic digestion. Once the resorption site is created, Ob intrude and begin new bone formation along with the secretion of osteoid, a mineralizing factor. The process is completed by coverage of the bone surface by lining cells, a type of terminally differentiated Ob [26] . Bone disease is caused by an imbalance in bone remodeling. At the physiological level, mechanisms of bone remodeling are regulated locally by cytokines and systemically by hormones [27] . PTH and 1,5-dihydroxy vitamin D have anabolic actions, which are opposed by calcitonin [27] -[29] . Several systemic regulators, such as, insulin-like growth factor (also acting through local regulators) [30] , glucocorticoids [31] , thyroid hormone [32] and estrogen [33] contribute to bone formation and bone resorption. In addition, local factors derived from bone cells, like TGF-β, have inhibitory functions on osteoclastogenesis [34] regulate cell growth and differentiation and induce anabolic activity in Ob [35] . IL-1 and TNF-α serve to quiesce Oc and participate in bone resorption pathophysiology [36] . The receptor activator of NF-κB ligand (RANKL), RANK and osteoprotegrin (OPG) are essential for Oc differentiation. RANKL is expressed by Ob as a soluble factor that binds with RANK on Oc with recruitment of tumor necrosis factor receptor-associated factor 6 (TRAF6) and activation of the transcription factor, nuclear factor of activated T-cells cytoplasmic 1 (NFATc1) to induce of Ob differentiation related genes via the NF-κB pathway [37] [38] . OPG is a decoy receptor for RANKL which plays an inhibitory role in osteoclastogenesis [39] . OPG-deficient mice exhibit a decrease in total bone density and develope osteoporosis [40] . The regulation of bone remodeling not only involves osteoblastic and osteoclastic cell lineages but also other marrow cells. Adipocytes are derived from the same progenitor cells as Ob, and the equilibrium between the two cell types is important in bone remodeling [41] [42] . Several regulatory pathways of adipocyte differentiation from bone marrow mesenchymal stem cells interact with osteoblast differentiation pathway [43] . The major adipogenic transcriptional factors, C/EBPα and PPARγ, are regulated by extracellular signaling involved in osteoblastgenesis [43] . Mesenchymal stem cell differentiation into Ob requires Wnt signaling activation, which stimulates osteoblastogenesis and suppresses adipogenesis by blocking PPARγ and C/EBPα [44] [45] . Cell lineage allocation in the bone marrow microenvironment is shown in Figure 2.

Figure 2. Cell lineage allocation and regulation in the bone marrow microenvironment (modified from [30]). Osteoblasts and adipocytes are derived from the same progenitors, the mesenchymal stem cells. To achieve full differentiated status, multiple critical transcription factors must be activated. RANKL is expressed by osteoblasts as a soluble factor, which binds to RANK on osteoclasts. Further recruitment of TRAF6 and activation of NFATc1 induce osteoclast differentiation. Cytokines released from the adipocytes participate in the bone remodeling process. (CEBPα, CAAT/enhancer binding protein α; PPARγ, peroxisome proliferative activated receptor γ; RANK, receptor activator of nuclear transcription factor κB; RANKL, receptor activator of nuclear transcription factor κB ligand; OPG, osteoprotegerin; IL, interleukin; MCSF, macrophage colony stimulating factor; MSX, MSH homeobox homolog; RUNX, runt-related transcription factor; TRAF6, tumor necrosis factor receptor-associated factor 6; NFATc1, nuclear factor of activated T-cells cytoplasmic 1).
4. The Actions of Adiponectin on Bone Metabolism
4.1. Effects of ApN on Bone Metabolism and on Osteoblast and Osteoclast Differentiation
ApN stimulates osteoblastogenesis and chondrocytogenesis, but suppresses osteoclastogenesis and therefore promotes bone formation [9] [46] . There are three distinct ApN actions in bone formation: 1) a direct positive endocrine action through circulating ApN; 2) an autocrine/paracrine action; 3) indirect endocrine effects by interacting with other signaling pathways such as insulin and bone morphogenetic protein 2 (BMP2).
Several studies confirm the positive endocrine role of ApN in bone formation. Challa et al. suggest that ApN increases chondrocyte proliferation, proteoglycan synthesis and matrix mineralization by upregulating the expression of type II collagen and Runx2 and increasing the activities of alkaline phosphatase [46] . Similarly, Oshima et al. (2005) demonstrated that ApN-treated C57BL/6J mice have increased trabecular bone mass and enhanced mineralization of Ob, accompanied by a decreased number of Oc and bone resorption activity [47] . Potential mechanisms for circulating ApN to modulate Oc differentiation are that ApN augmentes gene expression of several osteogenic markers and increases Ob differentiation in mesenchymal progenitor cells [48] . Moreover, ApN activates p38 mitogen-activated protein kinase via AdipoR1, which results in c-Jun activation and upregulation of the target gene, cyclooxygenase-2 (COX-2). ApN also stimulates BMP2 expression in a COX2- dependent manner and therefore increases Ob differentiation [48] . An alternative action of ApN, discovered by Huang et al. is that ApN stimulates osteoblast differentiation by increasing BMP-2 expression with involvement of AMPK, p38 and NF-κB [49] .
ApN regulates bone formation in an autocrine/paracrine manner. ApN and its receptors are expressed in osteoblastic and osteoclastic cells, indicating participation in bone metabolism not only through an endocrine pathway, but also locally in the bone [9] . The authors analyzed cultures of bone marrow cells from ApN-knockout (Ad−/−) and WT mice and find a significant decrease of osteogenesis in Ad−/− marrow cell culture, compared to WT. Collectively, these results suggest a positive autocrine/paracrine action of ApN on bone formation.
ApN also has indirect effects on bone, possibly through modulating growth factor action or insulin sensitivity. In the presence of insulin and BMP2 (but not IGF-1), ApN stimulates Ob differentiation in bone marrow cells [9] . Co-culture of Ob with the secretory products of adipocytes produces an inhibitory effect on Ob differentiation, which can be reversed by knockdown of adipoR1 [50] .
4.2. Involvement of Insulin Signaling, Long Term Adaption and Compensation in ApN-Modulated Bone Metabolism
Most studies indicate that the effects of ApN and its receptors have a positive role on bone metabolism, but some opposite aspects are also described [51] . One confounding factor is insulin signaling. Shinoda et al. (2006) treated a bone marrow cell culture with insulin or recombinant mouse ApN and found that insulin impairs the effect of ApN treatment on osteogenesis and restores the number of colonies. In addition, long term adaption and compensation in transgenic or knockout mice may modify ApN-modulated bone metabolism. Analyses of bone characteristics by radiological and histological examination of the femur, tibia and vertebrae in male WT and Ad−/− littermates indicates no difference between WT and Ad−/− mice [9] . Williams et al. (2009) also observed the same results in WT and Ad−/− 22 wk old mice [51] . In this circumstance, long term adaption and compensation produced no change in bone development in vivo in ApN knockout mice. Tu et al. (2011) isolated the bone cells from genetically double-labeled mBSP9.0Luc/β-ACT-EGFP transgenic mice and transplanted them into ApN−/− or wild type mice to investigate the effect of temporary exposure to ApN on bone growth and metabolism; growth of bone explants in ApN−/− mice is significantly retarded. Moreover, micro-CT analysis and tartrate-resistant acid phosphatase staining revealed decreased bone volume, cortical bone and increased Ob number in bone explants in ApN−/− mice [52] . ApN inhibits RANKL-induced osteoclastogenesis from RAW264.7 cells, down-regulates RANKL-enhanced osteoclastogenic regulators, and increases Oc apoptosis [52] . The effects of ApN on bone metabolism including the in vivo mouse model are shown in Table1
5. Involvement of AMPK in ApN-Modulated Bone Metabolism
Bone remodeling is an energy intensive process and bones need to balance energy constantly in response to nutrient availability during growth and bone turnover. During the last decade adenosine 5’-monophosphate-activated protein kinase (AMPK) emerged as a key in the regulation of energy homeostasis and as the mediator of

Table 1 . Effects of ApN on bone metabolism.
central and peripheral effects of numerous hormones [53] . In addition, AMPK signaling activates ApN receptor signaling pathways in liver, muscle and adipose tissues [54] -[57] . In vitro studies demonstrate that AMPK modulates bone cell differentiation and function. AMPK is expressed in mouse tibia and ROS 17/2.8 osteoblastic cells [58] . Murine Ob and human mesenchymal stem cells treated with AMPK activators (AICAR or metformin) stimulate Thr-172 phosphorylation of AMPK in a dose/time-dependent manner. In contrast, treatment with the AMPK inhibitor (compound C) or knockdown of AMPK by shRNA-lentivirus infection inhibites AMPK phosphorylation and induces osteogenesis [58] [59] . Shah et al. (2010) analyzed the bone phenotype of 4 month-old male WT and AMPKα1−/−knockout mice by micro-CT. The AMPKα1−/−KO mice have less trabecular bone [58] . Furthermore, AMPK acts as a negative regulator in differentiation of Oc [60] . Treatment of pre-Oc with compound C (AMPK inhibitor) potentiates bone resorption and formation of TRAP-positive multinucleated cells in a dose-dependent manner [60] . Treatment with the globular form of ApN inhibits TNF-α/ RANKL-induced Oc formation from a RAW264 clone via AMPK signaling [61] . The elucidation of the importance of AMPK signaling in bone is still in early stage, but already reveals that AMPK activation affects bone formation and bone mass; therefore, AMPK signaling might act as a significant pathway in skeletal physiology.
6. Conclusions and Future Perspectives
ApN and its receptors are expressed in bone and bone stromal cells which suggest that they play critical roles in bone metabolism. The present review summarizes effects of ApN on bone metabolism, both in physiological states and in Ob or Oc culture in vitro. There are still several controversial results on the correlation of ApN and bone mineral density or other bone parameters. The critical role of ApN in the bone remodeling process and bone metabolism is an indisputable fact. Herein, we propose some possible future research directions.
1) In the circulatory system, ApN exists multi-mer complexes, such as trimers, hexamers and HMW forms [16] . Several researches suggest that different forms of ApN may give rise to different activities and have different roles in insulin sensitivity [62] [63] . In both mice and human diabetic patients, an increased ratio of HMW ApN to total ApN in the plasma correlates with improvement in insulin sensitivity during treatment with an insulin-sensitizing drug, TZD; there is no corelation with total amount of ApN [63] . Only one research group indicates that HMW ApN affects bone metabolism in both male and female hemodialysis patients [64] . We speculate that different multi-mer forms of ApN cause different influences on bone formation, but more molecular level observations are needed to confirm this speculation.
2) Energy equilibrium in mammals is controlled by the actions of circulatory hormones that coordinate fuel production and utilization in metabolically active tissues. Bone remodeling can be affected by metabolic related hormones, implying that bones are involved in the control of energy homeostasis [65] [66] . Osteocalcin, the most abundant non-collagenous protein of the bone extracellular matrix regulates β-cell proliferation, insulin secretion and glucose homeostasis [66] [67] . Furthermore, there are several consistent results showing the insulin-osteocalcin endocrine loop in humans [68] [69] were decreased in adipocyte and pancreatic islets respectively co-culture with osteocalcin knockout osteoblasts [65] . Whether osteocalcin has any interaction with ApN in bone metabolism or as a feed-back loop via ApN needs further investigation.
3) Mao et al. [70] used the cytoplasmic domain of AdipoR1 as bait to screen a yeast two-hybrid cDNA library derived from human brain. They found that APPL1 (adaptor protein containing pleckstrin homology domain, phosphotryosine binding domain (PTB) and leucine zipper motif) interacted with ApN receptors in mammalian cells and the interaction was stimulated by ApN. It has been suggested that APPL1 mediates the effect of ApN on inhibition of osteoclastogenesis and bone resorption [52] . Although this observation is an indirect evidence, we speculate that APPL1 may have functions to mediate ApN effects on bone development. APPL1 not only has an AdipoR1 binding domain in the PTB domain, but also has a FSH receptor binding domain [71] . The existence of FSH receptor domain in APPL1 increases the possibility of APPL1 mediating the regulation of ApN on bone metabolism [71] [72] . In order to investigate this possible regulator, we have to further investigate the role of APPL1 by culture system or loss-of-function and gain-of-function model.
4) Epigenic regulation—MicroRNAs (miRNAs) are small endogenous RNA fragments (19 - 25 nt) that regulate gene expression by targeting mRNA in post-transcriptional stage [73] . Controling the differentiation of the mesenchymal stem cells into osteogenic or adipogenic lineage is important for maintaining healthy bone and is necessary for prevention of bone related disease. Recently, there are some evidences showing that the differentiation between Ob and adipocytes is tightly controlled by miRNAs [74] [75] . We speculate that miRNA may have the effects on regulating expression of ApN and therefore affect bone formation or bone resorption.
5) AdipoR2 is expressed in bone-forming cells [10] . So far, there is no evidence showing the effect of ApN through AdipoR2 in bone formation or bone resorption. The role of AdipoR2 in bone metabolism must be investigated.
6) Bone marrow microenvironment and its endocrine or paracrine regulatory systems are actively studied. Osteoporosis models such as ovariectomy and senescence have been used extensively in rodents or other mammals [76] [77] . Ovariectomy mimics postmenopausal women whose bone loss is caused from acute ovary hormonal deficiency. Ovariectomy destroys the overall hormonal balance, but is a difficult model to investigate changes in bone microenvironments. More appropriate models are needed to investigate regulatory mechanisms.
Acknowledgements
The study was supported by grants from the National Science Counsel in Taiwan.
References
- Meunier, P., Aaron, J., Edouard, C. and Vignon, G. (1971) Osteoporosis and the Replacement of Cell Populations of the Marrow by Adipose Tissue. A Quantitative Study of 84 Iliac Bone Biopsies. Clinical Orthopaedics and Related Research, 80, 147-154. http://dx.doi.org/10.1097/00003086-197110000-00021
- Gimble, J.M., Zvonic, S., Floyd, Z.E., Kassem, M. and Nuttall, M.E. (2006) Playing with Bone and Fat. Journal of Cellular Biochemistry, 98, 251-266. http://dx.doi.org/10.1002/jcb.20777
- Ahima, R.S. and Flier, J.S. (2000) Adipose Tissue as an Endocrine Organ. Trends in Endocrinology and Metabolism, 11, 327-332. http://dx.doi.org/10.1016/S1043-2760(00)00301-5
- Kershaw, E.E. and Flier, J.S. (2004) Adipose Tissue as an Endocrine Organ. The Journal of Clinical Endocrinology and Metabolism, 89, 2548-2556. http://dx.doi.org/10.1210/jc.2004-0395
- Ducy, P., Amling, M., Takeda, S., Priemel, M., Schilling, A.F., Beil, F.T., et al. (2000) Leptin Inhibits Bone Formation through a Hypothalamic Relay: A Central Control of Bone Mass. Cell, 100, 197-207. http://dx.doi.org/10.1016/S0092-8674(00)81558-5
- Karsenty, G. (2006) Convergence between Bone and Energy Homeostases: Leptin Regulation of Bone Mass. Cell Metabolism, 4, 341-348. http://dx.doi.org/10.1016/j.cmet.2006.10.008
- Yamauchi, T. and Kadowaki, T. (2008) Physiological and Pathophysiological Roles of Adiponectin and Adiponectin Receptors in the Integrated Regulation of Metabolic and Cardiovascular Diseases. International Journal of Obesity, 32, S13-18. http://dx.doi.org/10.1038/ijo.2008.233
- Kelesidis, I., Kelesidis, T. and Mantzoros, C.S. (2006) Adiponectin and Cancer: A Systematic Review. British Journal of Cancer, 94, 1221-1225. http://dx.doi.org/10.1038/sj.bjc.6603051
- Shinoda, Y., Yamaguchi, M., Ogata, N., Akune, T., Kubota, N., Yamauchi, T., et al. (2006) Regulation of Bone Formation by Adiponectin through Autocrine/Paracrine and Endocrine Pathways. Journal of Cellular Biochemistry, 99, 196-208. http://dx.doi.org/10.1002/jcb.20890
- Berner, H.S., Lyngstadaas, S.P., Spahr, A., Monjo, M., Thommesen, L., Drevon, C.A., et al. (2004) Adiponectin and Its Receptors Are Expressed in Bone-Forming Cells. Bone, 35, 842-849. http://dx.doi.org/10.1016/j.bone.2004.06.008
- Scherer, P.E., Williams, S., Fogliano, M., Baldini, G. and Lodish, H.F. (1995) A Novel Serum Protein Similar to C1q, Produced Exclusively in Adipocytes. The Journal of Biological Chemistry, 270, 26746-26749. http://dx.doi.org/10.1074/jbc.270.45.26746
- Hu, E., Liang, P. and Spiegelman, B.M. (1996) AdipoQ is a Novel Adipose-Specific Gene Dysregulated in Obesity. The Journal of Biological Chemistry, 271, 10697-10703. http://dx.doi.org/10.1074/jbc.271.18.10697
- Maeda, K., Okubo, K., Shimomura, I., Funahashi, T., Matsuzawa, Y. and Matsubara, K. (1996) cDNA Cloning and Expression of a Novel Adipose Specific Collagen-Like Factor, apM1 (AdiPose Most abundant Gene Transcript 1). Biochemical and Biophysical Research Communications, 221, 286-289. http://dx.doi.org/10.1006/bbrc.1996.0587
- Nakano, Y., Tobe, T., Choi-Miura, N., Mazda, T. and Tomita, M. (1996) Isolation and Characterization of GBP28, a Novel Gelatin-Binding Protein Purified from Human Plasma. Journal of Biochemistry, 120, 803-812. http://dx.doi.org/10.1093/oxfordjournals.jbchem.a021483
- Wong, G.W., Wang, J., Hug, C., Tsao, T.S. and Lodish, H.F. (2004) A Family of Acrp30/Adiponectin Structural and Functional Paralogs. Proceedings of the National Academy of Sciences of the United States of America, 101, 10302-10307. http://dx.doi.org/10.1073/pnas.0403760101
- Okamoto, Y., Kihara, S., Funahashi, T., Matsuzawa, Y. and Libby, P. (2006) Adiponectin: A Key Adipocytokine in Metabolic Syndrome. Clinical Science, 110, 267-278. http://dx.doi.org/10.1042/CS20050182
- Kubota, N., Terauchi, Y., Yamauchi, T., Kubota, T., Moroi, M., Matsui, J., et al. (2002) Disruption of Adiponectin Causes Insulin Resistance and Neointimal Formation. The Journal of Biological Chemistry, 277, 25863-25866.http://dx.doi.org/10.1074/jbc.C200251200
- Fruebis, J., Tsao, T.S., Javorschi, S., Ebbets-Reed, D., Erickson, M.R., Yen, F.T., et al. (2001) Proteolytic Cleavage Product of 30-kDa Adipocyte Complement-Related Protein Increases Fatty Acid Oxidation in Muscle and Causes Weight Loss in Mice. Proceedings of the National Academy of Sciences of the United States of America, 98, 2005-2010. http://dx.doi.org/10.1073/pnas.98.4.2005
- Yamauchi, T., Kamon, J., Waki, H., Terauchi, Y., Kubota, N., Hara, K., et al. (2001) The Fat-Derived Hormone Adiponectin Reverses Insulin Resistance Associated with both Lipoatrophy and Obesity. Nature Medicine, 7, 941-946.http://dx.doi.org/10.1038/90984
- Yamauchi, T., Kamon, J., Ito, Y., Tsuchida, A., Yokomizo, T., Kita, S., et al. (2003) Cloning of Adiponectin Receptors That Mediate Antidiabetic Metabolic Effects. Nature, 423, 762-769. http://dx.doi.org/10.1038/nature01705
- Ding, S.T., Liu, B.H. and Ko, Y.H. (2004) Cloning and Expression of Porcine Adiponectin and Adiponectin Receptor 1 and 2 Genes in Pigs. Journal of Animal Science, 82, 3162-3174.
- Yamauchi, T., Nio, Y., Maki, T., Kobayashi, M., Takazawa, T., Iwabu, M., et al. (2007) Targeted Disruption of AdipoR1 and AdipoR2 Causes Abrogation of Adiponectin Binding and Metabolic Actions. Nature Medicine, 13, 332-339.http://dx.doi.org/10.1038/nm1557
- Yamauchi, T. and Kadowaki, T. (2013) Adiponectin Receptor as a Key Player in Healthy Longevity and Obesity-Related Diseases. Cell Metabolism, 17, 185-196. http://dx.doi.org/10.1016/j.cmet.2013.01.001
- Teitelbaum, S.L. (2000) Bone Resorption by Osteoclasts. Science, 289, 1504-1508.http://dx.doi.org/10.1126/science.289.5484.1504
- Owen, M. (1988) Marrow Stromal Stem Cells. Journal of Cell Science, 63-76.http://dx.doi.org/10.1242/jcs.1988.Supplement_10.5
- Manolagas, S.C. and Jilka, R.L. (1995) Bone Marrow, Cytokines, and Bone Remodeling—Emerging Insights into the Pathophysiology of Osteoporosis. The New England Journal of Medicine, 332, 305-311.http://dx.doi.org/10.1056/NEJM199502023320506
- Raisz, L.G. (1999) Physiology and Pathophysiology of Bone Remodeling. Clinical Chemistry, 45, 1353-1358.
- Dempster, D.W., Cosman, F., Parisien, M., Shen, V. and Lindsay, R. (1993) Anabolic Actions of Parathyroid Hormone on bone. Endocrine Reviews, 14, 690-709.
- Li, Y.C., Amling, M., Pirro, A.E., Priemel, M., Meuse, J., Baron, R., et al. (1998) Normalization of Mineral Ion Homeostasis by Dietary Means Prevents Hyperparathyroidism, Rickets, and Osteomalacia, But Not Alopecia in Vitamin D Receptor-Ablated Mice. Endocrinology, 139, 4391-4396.
- Rosen, C.J. and Donahue, L.R. (1998) Insulin-Like Growth Factors and Bone: The Osteoporosis Connection Revisited. Experimental Biology and Medicine, 219, 1-7. http://dx.doi.org/10.3181/00379727-219-44310
- Advani, S., LaFrancis, D., Bogdanovic, E., Taxel, P., Raisz, L.G. and Kream, B.E. (1997) Dexamethasone Suppresses in Vivo Levels of Bone Collagen Synthesis in Neonatal Mice. Bone, 20, 41-46.http://dx.doi.org/10.1016/S8756-3282(96)00314-6
- Kawaguchi, H., Pilbeam, C.C. and Raisz, L.G. (1994) Anabolic Effects of 3,3’,5-Triiodothyronine and Triiodothyroacetic Acid in Cultured Neonatal Mouse Parietal Bones. Endocrinology, 135, 971-976.
- Pacifici, R. (1998) Cytokines, Estrogen, and Postmenopausal Osteoporosis—The Second Decade. Endocrinology, 139, 2659-2661.
- Takai, H., Kanematsu, M., Yano, K., Tsuda, E., Higashio, K., Ikeda, K., et al. (1998) Transforming Growth Factor-Beta Stimulates the Production of Osteoprotegerin/Osteoclastogenesis Inhibitory Factor by Bone Marrow Stromal Cells. The Journal of Biological Chemistry, 273, 27091-27096. http://dx.doi.org/10.1074/jbc.273.42.27091
- Ramirez-Yanez, G.O., Hamlet, S., Jonarta, A., Seymour, G.J. and Symons, A.L. (2006) Prostaglandin E2 Enhances Transforming Growth Factor-Beta 1 and TGF-Beta Receptors Synthesis: An in Vivo and in Vitro Study. Prostaglandins, Leukotrienes and Essential Fatty Acids, 74, 183-192. http://dx.doi.org/10.1016/j.plefa.2006.01.003
- Steeve, K.T., Marc, P., Sandrine, T., Dominique, H. and Yannick, F. (2004) IL-6, RANKL, TNF-Alpha/IL-1: Interrelations in Bone Resorption Pathophysiology. Cytokine and Growth Factor Reviews, 15, 49-60.http://dx.doi.org/10.1016/j.cytogfr.2003.10.005
- Takayanagi, H., Kim, S., Koga, T., Nishina, H., Isshiki, M., Yoshida, H., et al. (2002) Induction and Activation of the Transcription Factor NFATc1 (NFAT2) Integrate RANKL Signaling in Terminal Differentiation of Osteoclasts. Developmental Cell, 3, 889-901. http://dx.doi.org/10.1016/S1534-5807(02)00369-6
- Nakashima, T., Hayashi, M. and Takayanagi, H. (2012) New Insights into Osteoclastogenic Signaling Mechanisms. Trends in Endocrinology and Metabolism, 23, 582-590. http://dx.doi.org/10.1016/j.tem.2012.05.005
- Lacey, D.L., Timms, E., Tan, H.L., Kelley, M.J., Dunstan, C.R., Burgess, T., et al. (1998) Osteoprotegerin Ligand Is a Cytokine That Regulates Osteoclast Differentiation and Activation. Cell, 93, 165-176.http://dx.doi.org/10.1016/S0092-8674(00)81569-X
- Bucay, N., Sarosi, I., Dunstan, C.R., Morony, S., Tarpley, J., Capparelli, C., et al. (1998) Osteoprotegerin-Deficient Mice Develop Early Onset Osteoporosis and Arterial Calcification. Genes and Development, 12, 1260-1268.http://dx.doi.org/10.1101/gad.12.9.1260
- Chamberlain, G., Fox, J., Ashton, B. and Middleton, J. (2007) Concise Review: Mesenchymal Stem Cells: Their Phenotype, Differentiation Capacity, Immunological Features, and Potential for Homing. Stem Cells, 25, 2739-2749.http://dx.doi.org/10.1634/stemcells.2007-0197
- Duque, G. (2008) Bone and Fat Connection in Aging Bone. Current Opinion in Rheumatology, 20, 429-434.http://dx.doi.org/10.1097/BOR.0b013e3283025e9c
- Muruganandan, S., Roman, A.A. and Sinal, C.J. (2009) Adipocyte Differentiation of Bone Marrow-Derived Mesenchymal Stem Cells: Cross Talk with the Osteoblastogenic Program. Cellular and Molecular Life Sciences, 66, 236-253.http://dx.doi.org/10.1007/s00018-008-8429-z
- Ross, S.E., Hemati, N., Longo, K.A., Bennett, C.N., Lucas, P.C., Erickson, R.L., et al. (2000) Inhibition of Adipogenesis by Wnt Signaling. Science, 289, 950-953. http://dx.doi.org/10.1126/science.289.5481.950
- Krishnan, V., Bryant, H.U. and Macdougald, O.A. (2006) Regulation of Bone Mass by Wnt Signaling. Journal of Clinical Investication, 116, 1202-1209. http://dx.doi.org/10.1172/JCI28551
- Challa, T.D., Rais, Y. and Ornan, E.M. (2010) Effect of Adiponectin on ATDC5 Proliferation, Differentiation and Signaling Pathways. Molecular and Cellular Endocrinology, 323, 282-291. http://dx.doi.org/10.1016/j.mce.2010.03.025
- Oshima, K., Nampei, A., Matsuda, M., Iwaki, M., Fukuhara, A., Hashimoto, J., et al. (2005) Adiponectin Increases Bone Mass by Suppressing Osteoclast and Activating Osteoblast. Biochemical and Biophysical Research Communications, 331, 520-526. http://dx.doi.org/10.1016/j.bbrc.2005.03.210
- Lee, H.W., Kim, S.Y., Kim, A.Y., Lee, E.J., Choi, J.Y. and Kim, J.B. (2009) Adiponectin Stimulates Osteoblast Differentiation through Induction of COX2 in Mesenchymal Progenitor Cells. Stem Cells, 27, 2254-2262.http://dx.doi.org/10.1002/stem.144
- Huang, C.Y., Lee, C.Y., Chen, M.Y., Tsai, H.C., Hsu, H.C. and Tang, C.H. (2010) Adiponectin Increases BMP-2 Expression in Osteoblasts via AdipoR Receptor Signaling Pathway. Journal of Cellular Physiology, 224, 475-483.http://dx.doi.org/10.1002/jcp.22145
- Liu, L.F., Shen, W.J., Zhang, Z.H., Wang, L.J. and Kraemer, F.B. (2010) Adipocytes Decrease Runx2 Expression in Osteoblastic Cells: Roles of PPARgamma and Adiponectin. Journal of Cellular Physiology, 225, 837-845.http://dx.doi.org/10.1002/jcp.22291
- Williams, G.A., Wang, Y., Callon, K.E., Watson, M., Lin, J.M., Lam, J.B., et al. (2009) In Vitro and in Vivo Effects of Adiponectin on Bone. Endocrinology, 150, 3603-3610. http://dx.doi.org/10.1210/en.2008-1639
- Tu, Q., Zhang, J., Dong, L.Q., Saunders, E., Luo, E., Tang, J., et al. (2011) Adiponectin Inhibits Osteoclastogenesis and Bone Resorption via APPL1-Mediated Suppression of Akt1. The Journal of Biological Chemistry, 286, 12542-12553. http://dx.doi.org/10.1074/jbc.M110.152405
- Jeyabalan, J., Shah, M., Viollet, B. and Chenu, C. (2012) AMP-Activated Protein Kinase Pathway and Bone Metabolism. Journal of Endocrinology, 212, 277-290. http://dx.doi.org/10.1530/JOE-11-0306
- Kadowaki, T., Yamauchi, T., Kubota, N., Hara, K., Ueki, K. and Tobe, K. (2006) Adiponectin and Adiponectin Receptors in Insulin Resistance, Diabetes, and the Metabolic Syndrome. The Journal of Clinical Investigation, 116, 1784-1792. http://dx.doi.org/10.1172/JCI29126
- Yamauchi, T., Kamon, J., Minokoshi, Y., Ito, Y., Waki, H., Uchida, S., et al. (2002) Adiponectin Stimulates Glucose Utilization and Fatty-Acid Oxidation by Activating AMP-Activated Protein Kinase. Nature Medicine, 8, 1288-1295.http://dx.doi.org/10.1038/nm788
- Tomas, E., Tsao, T.S., Saha, A.K., Murrey, H.E., Zhang, C.C., Itani, S.I., et al. (2002) Enhanced Muscle Fat Oxidation and Glucose Transport by ACRP30 Globular Domain: Acetyl-CoA Carboxylase Inhibition and AMP-Activated Protein Kinase Activation. Proceedings of the National Academy of Sciences of the United States of America, 99, 16309-16313. http://dx.doi.org/10.1073/pnas.222657499
- Berg, A.H., Combs, T.P., Du, X., Brownlee, M. and Scherer, P.E. (2001) The Adipocyte-Secreted Protein Acrp30 Enhances Hepatic Insulin Action. Nature Medicine, 7, 947-953. http://dx.doi.org/10.1038/90992
- Shah, M., Kola, B., Bataveljic, A., Arnett, T.R., Viollet, B., Saxon, L., et al. (2010) AMP-Activated Protein Kinase (AMPK) Activation Regulates in Vitro Bone Formation and Bone Mass. Bone, 47, 309-319.http://dx.doi.org/10.1016/j.bone.2010.04.596
- Kim, E.K., Lim, S., Park, J.M., Seo, J.K., Kim, J.H., Kim, K.T., et al. (2012) Human Mesenchymal Stem Cell Differentiation to the Osteogenic or Adipogenic Lineage Is Regulated by AMP-Activated Protein Kinase. Journal of Cellular Physiology, 227, 1680-1687. http://dx.doi.org/10.1002/jcp.22892
- Lee, Y.S., Kim, Y.S., Lee, S.Y., Kim, G.H., Kim, B.J., Lee, S.H., et al. (2010) AMP Kinase Acts as a Negative Regulator of RANKL in the Differentiation of Osteoclasts. Bone, 47, 926-937. http://dx.doi.org/10.1016/j.bone.2010.08.001
- Yamaguchi, N., Kukita, T., Li, Y.J., Kamio, N., Fukumoto, S., Nonaka, K., et al. (2008) Adiponectin Inhibits Induction of TNF-Alpha/RANKL-Stimulated NFATc1 via the AMPK Signaling. FEBS Letters, 582, 451-456.http://dx.doi.org/10.1016/j.febslet.2007.12.037
- Waki, H., Yamauchi, T., Kamon, J., Ito, Y., Uchida, S., Kita, S., et al. (2003) Impaired Multimerization of Human Adiponectin Mutants Associated with Diabetes. Molecular Structure and Multimer Formation of Adiponectin. The Journal of Biological Chemistry, 278, 40352-40363. http://dx.doi.org/10.1074/jbc.M300365200
- Pajvani, U.B., Hawkins, M., Combs, T.P., Rajala, M.W., Doebber, T., Berger, J.P., et al. (2004) Complex Distribution, Not Absolute Amount of Adiponectin, Correlates with Thiazolidinedione-Mediated Improvement in Insulin Sensitivity. The Journal of Biological Chemistry, 279, 12152-12162. http://dx.doi.org/10.1074/jbc.M311113200
- Amemiya, N., Otsubo, S., Iwasa, Y., Onuki, T. and Nitta, K. (2012) Association between High-Molecular-Weight Adiponectin and Bone Mineral Density in Hemodialysis Patients. Clinical and Experimental Nephrology, 17, 411-415.http://dx.doi.org/10.1007/s10157-012-0723-2
- Lee, N.K., Sowa, H., Hinoi, E., Ferron, M., Ahn, J.D., Confavreux, C., et al. (2007) Endocrine Regulation of Energy Metabolism by the Skeleton. Cell, 130, 456-469. http://dx.doi.org/10.1016/j.cell.2007.05.047
- Fulzele, K., Riddle, R.C., DiGirolamo, D.J., Cao, X., Wan, C. and Chen, D. (2010) Insulin Receptor Signaling in Osteoblasts Regulates Postnatal Bone Acquisition and Body Composition. Cell, 142, 309-319.http://dx.doi.org/10.1016/j.cell.2010.06.002
- Ferron, M., Wei, J., Yoshizawa, T., Del Fattore, A., DePinho, R.A., Teti, A., et al. (2010) Insulin Signaling in Osteoblasts Integrates Bone Remodeling and Energy Metabolism. Cell, 142, 296-308. http://dx.doi.org/10.1016/j.cell.2010.06.003
- Hwang, Y.C., Jeong, I.K., Ahn, K.J. and Chung, H.Y. (2012) Circulating Osteocalcin Level Is Associated with Improved Glucose Tolerance, Insulin Secretion and Sensitivity Independent of the Plasma Adiponectin Level. Osteoporosis International, 23, 1337-1342. http://dx.doi.org/10.1007/s00198-011-1679-x
- Hwang, Y.C., Jeong, I.K., Ahn, K.J. and Chung, H.Y. (2009) The Uncarboxylated Form of Osteocalcin Is Associated with Improved Glucose Tolerance and Enhanced Beta-Cell Function in Middle-Aged Male Subjects. Diabetes/Metabolism Research and Reviews, 25, 768-772.
- Mao, X., Kikani, C.K., Riojas, R.A., Langlais, P., Wang, L., Ramos, F.J., et al. (2006) APPL1 Binds to Adiponectin Receptors and Mediates Adiponectin Signalling and Function. Nature Cell Biology, 8, 516-523.http://dx.doi.org/10.1038/ncb1404
- Deepa, S.S. and Dong, L.Q. (2009) APPL1: Role in Adiponectin Signaling and Beyond. American Journal of Physiology, Endocrinology and Metabolism, 296, E22-E36. http://dx.doi.org/10.1152/ajpendo.90731.2008
- Prior, J.C. (2007) FSH and Bone—Important Physiology or Not? Trends in Molecular Medicine, 13, 1-3.http://dx.doi.org/10.1016/j.molmed.2006.11.004
- Lee, R.C., Feinbaum, R.L. and Ambros, V. (1993) The C. Elegans Heterochronic Gene Lin-4 Encodes Small RNAs with Antisense Complementarity to Lin-14. Cell, 75, 843-854. http://dx.doi.org/10.1016/0092-8674(93)90529-Y
- Guo, L., Zhao, R.C. and Wu, Y. (2011) The Role of MicroRNAs in Self-Renewal and Differentiation of Mesenchymal Stem Cells. Experimental Hematology, 39, 608-616. http://dx.doi.org/10.1016/j.exphem.2011.01.011
- Lian, J.B., Stein, G.S., van Wijnen, A.J., Stein, J.L., Hassan, M.Q., Gaur, T., et al. (2012) MicroRNA Control of Bone Formation and Homeostasis. Nature Reviews Endocrinology, 8, 212-227. http://dx.doi.org/10.1038/nrendo.2011.234
- Turner, R.T., Maran, A., Lotinun, S., Hefferan, T., Evans, G.L., Zhang, M., et al. (2001) Animal Models for Osteoporosis. Reviews in Endocrine and Metabolic Disorders, 2, 117-127. http://dx.doi.org/10.1023/A:1010067326811
- Pietschmann, P., Skalicky, M., Kneissel, M., Rauner, M., Hofbauer, G., Stupphann, D., et al. (2007) Bone Structure and Metabolism in a Rodent Model of Male Senile Osteoporosis. Experimental Gerontology, 42, 1099-1108.http://dx.doi.org/10.1016/j.exger.2007.08.008
NOTES

*Corresponding author.