Natural Science Vol.07 No.03(2015), Article ID:54398,13 pages
10.4236/ns.2015.73012
Equilibrium Allele Distribution in Trading Populations
Gilles Saint-Paul
Paris School of Economics, New York University, Abu Dhabi, UAE
Email: gstpaulmail@gmail.com
Copyright © 2015 by author and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 13 February 2015; accepted 3 March 2015; published 4 March 2015
ABSTRACT
This paper extends the results of Saint-Paul (2007) regarding the long-run survival rates of alleles in trading populations, to a more general context where the number of loci is arbitrarily large under general assumptions about sexual reproduction. The central result is that fitness-reducing alleles can survive in a trading population, provided their frequency is not too large. However, the greater the number of loci that matter for fitness, the more stringent the conditions under which these alleles can survive.
Keywords:
Gene-Culture Coevolution, Markets, Division of Labor, Population Genetics

1. Introduction
Can culture affect the genetic makeup of a population? While this question has been dealt with some detail regarding cultural institutions such as cooperation and social norms,1 there is much less work dealing with a key component of culture: markets.2 Do we expect populations who trade for long enough to develop a different distribution of alleles compared with population where individuals remain in relative autarky?
In Saint-Paul [8] ,3 I consider the evolution of the gene pool in a population under alternative economic institutions, and show that alleles that cannot survive natural selection under autarky can survive under trade, because individuals can specialize in activities so as to avoid the fitness disadvantages associated with these alleles. The results are based on a very simplified representation of sexual reproduction, with only one chromo- some (instead of pairs of chromosomes), and only two loci that determine the individual’s productivity at two activities that affect fitness.
This paper generalizes these results for a more general system of sexual reproduction, with an arbitrary number of chromosomes and loci. Its contribution is twofold. First, it provides a set of assumptions under which one can meaningfully state that some alleles dominate their alternatives and eventually eliminate them in the long run. Second, it extends the results in Saint-Paul [8] by characterizing the distribution of alleles for a trading population in a long-run equilibrium (LRE), defined as a stationary distribution of alleles which is also an equilibrium in an economic sense.
The central result is that fitness-reducing alleles can survive in a trading population, provided their frequency is not too large. However, the greater the number of loci that matter for fitness, the more stringent the conditions under which these alleles can survive. That means that in the long run, we expect low alleles to survive only at a relatively small number of loci. Knowing more about the long-run distribution of alleles when their initial distribution does not satisfy the conditions for an LRE would involve analyzing the dynamics, which I do not do here but is an interesting topic for further research.
2. Notations and Genetic Properties of Stationary Populations
A genotype consists of an
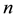 -tuple
-tuple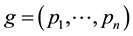 , where
, where
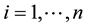 denotes a particular locus, and
denotes a particular locus, and
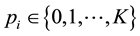 is interpreted as the number of alleles of the “high type” at locus
is interpreted as the number of alleles of the “high type” at locus
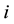 (in the actual world where chromosomes come by pairs, one has
(in the actual world where chromosomes come by pairs, one has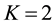 ). Therefore, there are
). Therefore, there are
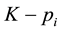 alleles of the “low type” at locus
alleles of the “low type” at locus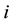 . The set of possible genotypes is denoted by
. The set of possible genotypes is denoted by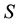 . We will also denote by
. We will also denote by
 the
the
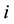 th element
of
th element
of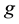 .
.
2.1. The Survival Function
The survival rate of an individual only depends on its genotype, and is denoted
by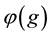 . Note that
the
. Note that
the
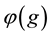 function is not independent of culture. The opportunity to trade and specialize
will dramatically change the
function is not independent of culture. The opportunity to trade and specialize
will dramatically change the
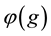 mapping. It
is useful to introduce the genetic improvement operators
mapping. It
is useful to introduce the genetic improvement operators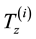 , which, for any genotype
, which, for any genotype
 such that
such that ,
maps it into another genotype
,
maps it into another genotype ,
defined by
,
defined by ,
,
 and
and .
Note that
.
Note that .
.
The survival function is monotonic at locus
 if it satisfies
if it satisfies
 (1)
(1)
Thus, having more of a high allele at locus
 cannot increase mortality, everything else equal. Note that this assumes that the
role played by an allele in mortality has the same sign regardless of what other
alleles are present.
cannot increase mortality, everything else equal. Note that this assumes that the
role played by an allele in mortality has the same sign regardless of what other
alleles are present.
We will say that a locus
 is selective if
is selective if
 (2)
(2)
2.2. The Distribution of Offsprings
We assume a quite general process for transmitting genes to offsprings, which in
particular is compatible with real-world genetics. When genotypes
 and
and
 mate, the fraction of their offsprings with genotype
mate, the fraction of their offsprings with genotype
 is given by a probability distribution function
is given by a probability distribution function .
We shall assume that it satisfies the following properties:
.
We shall assume that it satisfies the following properties:
1. Gene conservation
 (3)
(3)
This says that on average, the number of high alleles at locus
 among offsprings, denoted by
among offsprings, denoted by ,
is equal to the its average between the two parents. For a given pair of parents,
the average among actual off- springs will be different from the parental average.
However, with a continuum of individuals, the law of large number will apply, and
,
is equal to the its average between the two parents. For a given pair of parents,
the average among actual off- springs will be different from the parental average.
However, with a continuum of individuals, the law of large number will apply, and
 will be equal to the population average of the number of H-alleles at
will be equal to the population average of the number of H-alleles at
 among all offsprings of all couples with genotypes
among all offsprings of all couples with genotypes
 and
and .
.
2. Allele independence
 (4)
(4)
This assumption tells us that, among offsprings with the same parental genotypes,
the distribution of other genes among those who have the same number of high alleles
at locus ,
does not depend on that particular number. If that property did not hold, having
many good alleles at one locus could in principle be systematically correlated with
having many bad alleles at another locus, and this complementarity could sustain
a positive amount of mortality-increasing alleles in the long-run, or, conversely,
eliminate mortality-reducing ones.
,
does not depend on that particular number. If that property did not hold, having
many good alleles at one locus could in principle be systematically correlated with
having many bad alleles at another locus, and this complementarity could sustain
a positive amount of mortality-increasing alleles in the long-run, or, conversely,
eliminate mortality-reducing ones.
3. Mixing
For any ,
,
 ,
,
 for
all
for
all
 such that
such that
 (5)
(5)
there exists
 such that
such that ,
,
 for
for ,
and
,
and .
.
The RHS of (5) is the maximum number of H-alleles at locus
 if one inherits
if one inherits
 alleles from each parent; the LHS is the minimum number of H-alleles. That assumptions
says that for any number between these two bounds, there is a positive probability
for a couple
alleles from each parent; the LHS is the minimum number of H-alleles. That assumptions
says that for any number between these two bounds, there is a positive probability
for a couple ,
,
 to
have an offspring with exactly that number. Furthermore, we can pick up that offspring
such that at all other loci, its number of high alleles is between that of its two
parents. Loosely speaking, that means that the distribution of offsprings spans
all possible cases.
to
have an offspring with exactly that number. Furthermore, we can pick up that offspring
such that at all other loci, its number of high alleles is between that of its two
parents. Loosely speaking, that means that the distribution of offsprings spans
all possible cases.
4. Symmetry
 (6)
(6)
5. Monotonicity
For any ,
any
,
any ,
any
,
any
 such that
such that ,
and any
,
and any
 such that
such that

 (7)
(7)
 (8)
(8)
This assumption says that if instead of ,
a genetically improved genotype at locus
,
a genetically improved genotype at locus
 mates with
mates with ,
then holding the alleles at other loci constant, the proportion of H-alleles at
locus
,
then holding the alleles at other loci constant, the proportion of H-alleles at
locus
 improves in a first-order stochastic dominance sense: offsprings are more likely
to have a higher umber of H-alleles at
improves in a first-order stochastic dominance sense: offsprings are more likely
to have a higher umber of H-alleles at .
Formally, applying the
.
Formally, applying the
 operators starting from an initial genotype
operators starting from an initial genotype
 such that
such that ,
allows to compute the marginal distribution of
,
allows to compute the marginal distribution of
 among offsprings holding other alleles constant. The last equality says that the
partial distribution of the genotype at all other loci except
among offsprings holding other alleles constant. The last equality says that the
partial distribution of the genotype at all other loci except
 is invariant when one mates with a genetic improvement of
is invariant when one mates with a genetic improvement of
 at
at
 instead of
instead of .
.
2.3. Demographics
These assumptions allow to write down the demographic evolution equations of each
genotype. We denote by
 total population at date
total population at date
 and by
and by
 the fraction of people with genotype
the fraction of people with genotype .
People mate randomly.
.
People mate randomly.
There are
 matches of types
matches of types
 and
and
 at date
at date .
They produce
.
They produce
 offpsrings, and a frac- tion
offpsrings, and a frac- tion
 of offsprings with genotype
of offsprings with genotype
 reach maturity. Consequently,
reach maturity. Consequently,
 evolves
according to
evolves
according to

Adding all these equations across all possible genotypes we get that

It is also useful to define the population frequency of high alleles at locus :
:

Note that if the gene conservation law holds, then one also has
 (9)
(9)
3. Elimination of Less Fit Alleles
In this section, I provide the basic results regarding the elimination of less fit
alleles. A first lemma, which derives from the random mating and mixing properties,
states that if a genotype exists and if a high allele exists in the population at
locus ,
then we can find another genotype that differs from it only in that it is “improved”
at locus
,
then we can find another genotype that differs from it only in that it is “improved”
at locus ,
unless, of course, the initial genotype has the maximum number of H-alleles at
,
unless, of course, the initial genotype has the maximum number of H-alleles at .
.
LEMMA 1―Assume the mixing property holds. Assume there exists a steady state, a
locus
 and a genotype
and a genotype
 such that in that steady state,
such that in that steady state,
 ,
,
 ,
and
,
and .
Then
.
Then .
.
PROOF―First note that because of random mating there exists a positive measure of matches between two arbitrary genotypes, provided these genotypes are in positive measure in the parent population.
If ,
the mixing property applied at locus
,
the mixing property applied at locus
 implies that offsprings of
implies that offsprings of
 with itself include
with itself include
 with positive probability. Assume
with positive probability. Assume .
Since
.
Since ,
there exists
,
there exists
 such that
such that
 and
and .
We can then iterate the mixing property, by looking at stage
.
We can then iterate the mixing property, by looking at stage
 at the mates between
at the mates between
 and
and ,
starting with
,
starting with .
If at stage
.
If at stage ,
there exists
,
there exists
 such that
such that ,
say
,
say ,
by applying
,
by applying
the mixing property at locus
 we know that among the offsprings between
we know that among the offsprings between
 and
and ,
there exists one
,
there exists one
 such
that
such
that ―implying
―implying
 in steady state―
in steady state― ,
and
,
and
 .
In other words, the “genetic distance” between
.
In other words, the “genetic distance” between
 and
and
 strictly goes down with
strictly goes down with .
Once we have reached the stage where
.
Once we have reached the stage where
 for all
for all ,
we apply the same procedure to locus
,
we apply the same procedure to locus ,
until we have produced an offspring such that
,
until we have produced an offspring such that ,
,
 and
and .
.
At that stage .
Q.E.D.
.
Q.E.D.
The following key result tells us that genes which increase mortality eventually disappear:
PROPOSITION 1―Assume that one of these two conditions holds:
(i) locus
 is selective, OR
is selective, OR
(ii)
 is
monotonic at
is
monotonic at
 and there exists one genotype
and there exists one genotype
 such that
such that
 in steady state,
in steady state,
 ,
and
,
and .
.
Assume (A3) and (A4) holds. Then in any steady state with ,
one must have
,
one must have .
.
PROOF―The frequency of the high allele at
 evolves according to
evolves according to

In steady state, we have that ,
,
 (10)
(10)
and
 (11)
(11)
The term
 can be rewritten as follows:
can be rewritten as follows:

That can be rewritten as:

This formula rests on the fact that all the genotypes such that
 can be deducted by applying the transform
can be deducted by applying the transform
 to all genotypes such that
to all genotypes such that .
.
Furthermore, the allele independence property implies that for
 such that
such that ,
,
 (12)
(12)
where
 is the total fraction of genotypes with
is the total fraction of genotypes with
 among the offsprings of
among the offsprings of
 and
and .4
Note that one must have
.4
Note that one must have
 (13)
(13)
Hence:

Now, if locus
 is selective, then
is selective, then
 is strictly increasing in
is strictly increasing in .
Consequently we have
.
Consequently we have
 (14)
(14)
This inequality rests on the fact that .
It holds with a strict inequality unless all the
.
It holds with a strict inequality unless all the
 but one are equal to zero.
but one are equal to zero.
We now show that unless
 or
or ,
there exists a pair of genotypes
,
there exists a pair of genotypes
 such that
such that ,
,
 ,
and (14) strictly holds. First note that if
,
and (14) strictly holds. First note that if ,
there exists a genotype
,
there exists a genotype
 such that
such that ,
and
,
and .5
Next, note that if there exists
.5
Next, note that if there exists
 such that
such that ,
the mixing property implies that for two parents of the same genotype
,
the mixing property implies that for two parents of the same genotype , there is a
positive probability of having an offpsring
, there is a
positive probability of having an offpsring
 such that
such that ,
for any
,
for any
 between
between
 and
and .
As long as
.
As long as
 and
and ,
there are more than two values of
,
there are more than two values of
 that satisfy that property. Consequently, there are at least two strictly positive
values of
that satisfy that property. Consequently, there are at least two strictly positive
values of ,
and one can take
,
and one can take .
.
Thus, if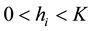 ,
it must be that there exists a pair
,
it must be that there exists a pair
 such that
such that ,
,
 ,
and (14) strictly holds.
,
and (14) strictly holds.
Alternatively, consider the case where
 is monotonic. Then (14) also holds. Furthermore, assume there exists
is monotonic. Then (14) also holds. Furthermore, assume there exists
 such that
such that ,
,
 ,
and
,
and .
Let
.
Let .
Then (14) will hold with strict inequality for
.
Then (14) will hold with strict inequality for ,
,
 such
that
such
that ,
,
 ,
,
 and
and .
If
.
If ,
,
taking
 and applying the mixing property to locus
and applying the mixing property to locus ,
generates both offsprings with
,
generates both offsprings with
 and
and
 implying
that
implying
that
 and
and .
If
.
If ,
the Lemma implies that
,
the Lemma implies that .
Taking
.
Taking
 and
and
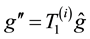 then generates both offsprings with
then generates both offsprings with
 and
and
 implying that
implying that
 and
and .
Thus, we can again pick up a genotype
.
Thus, we can again pick up a genotype
 and a pair
and a pair
 such that
such that ,
,
 and
(14) strictly holds.
and
(14) strictly holds.
From (14), we get that
 (15)
(15)
Once again, there exists a pair
 such that
such that ,
,
 ,
and (15) strictly holds. The RHS can be rewritten
,
and (15) strictly holds. The RHS can be rewritten

where the first step derives from (13) and the second one from (12).
Inequality (15) means that the fitness of the high alleles in the gene pool of the
offsprings of
 and
and
 is higher than the average fitness of the offsprings as individuals, because those
with more H-alleles at
is higher than the average fitness of the offsprings as individuals, because those
with more H-alleles at
 live longer. In order to get that, the allele invariance property is needed. Otherwise,
it could be that the offsprings of
live longer. In order to get that, the allele invariance property is needed. Otherwise,
it could be that the offsprings of ,
,
 that
have a high
that
have a high
 have a lower fitness than the others because they are systematically poorly endowed
at other loci.
have a lower fitness than the others because they are systematically poorly endowed
at other loci.
Going back to (11), we see that
 (16)
(16)
where the strict inequality comes from the fact that
 for at least one pair
for at least one pair
 such that
such that .
.
We now have
 (17)
(17)
where we have applied gene conservation and
 is defined as
is defined as

Observe that
 can be rewritten as
can be rewritten as

Furthermore, one can write
 where
where .
Iterating the monotonicity property, we find that
.
Iterating the monotonicity property, we find that
 is nonincreasing in
is nonincreasing in ,
while
,
while
 does not depend on
does not depend on .
We then have that
.
We then have that

Since
 is monotonic, the term in brackets is nonpositive. Thus, the sum is nondecreasing
in
is monotonic, the term in brackets is nonpositive. Thus, the sum is nondecreasing
in ,
while the last term is constant in
,
while the last term is constant in .
Therefore, the LHS is nondecreasing in
.
Therefore, the LHS is nondecreasing in ,
for any
,
for any
 such that
such that .
Summing this property across these
.
Summing this property across these ’s,
we also find that
’s,
we also find that
 is nondecreasing in
is nondecreasing in .
Roughly, that property means that the average mortality of offsprings improves when
one parent is genetically enhanced at locus
.
Roughly, that property means that the average mortality of offsprings improves when
one parent is genetically enhanced at locus .
The monotonicity property is needed to get that. Otherwise, it could be that parents
with more H-alleles at
.
The monotonicity property is needed to get that. Otherwise, it could be that parents
with more H-alleles at ,
everything else equal, have an
,
everything else equal, have an
 systematically biased toward high-mortality genotypes.
systematically biased toward high-mortality genotypes.
Let us now go back to (17), which we can rewrite

For a given ,
we have that
,
we have that ,
that
,
that
 increases with
increases with
 and that
and that
 weakly increases with
weakly increases with .
Thus, once again, we have the following inequality:
.
Thus, once again, we have the following inequality:

Consequently,

where the steady-state condition (10) has been used to derive the first term.
By virtue of (16), (3) and (6), the last term in that formula must be equal to ,
so that
,
so that .
(16) then implies that
.
(16) then implies that ,
which is a contradiction. Hence, it must be that either
,
which is a contradiction. Hence, it must be that either
 or
or .
Q.E.D.
.
Q.E.D.
The last set of inequalities tell us that since parents who have a greater
 have children with a higher fitness, these parents’ children tend to increase the
survival rate of the high allele at
have children with a higher fitness, these parents’ children tend to increase the
survival rate of the high allele at
 relative to average. Since, in addition, the survival rate of the high allele at
relative to average. Since, in addition, the survival rate of the high allele at
 among their children is greater than their children’s average survival rate, these
two effects together imply that the fitness of the high allele at
among their children is greater than their children’s average survival rate, these
two effects together imply that the fitness of the high allele at
 is strictly higher than average. But that cannot be in steady state, unless
is strictly higher than average. But that cannot be in steady state, unless
 or
or .
.
4. Autarky
We now describe how an individual’s genotype
 affects his/her productivity at various activities, depending on the ecomic setting.
affects his/her productivity at various activities, depending on the ecomic setting.
The alleles present at a given locus
 determine the individual’s productivity at a corresponding activity denoted by the
same index
determine the individual’s productivity at a corresponding activity denoted by the
same index .
This productivity is a strictly increasing function
.
This productivity is a strictly increasing function
 of
of ,
the number of H-alleles at locus
,
the number of H-alleles at locus .
Any individual has a total time endowment equal to 1. The time allocation constraint
of genotype
.
Any individual has a total time endowment equal to 1. The time allocation constraint
of genotype
 is therefore given by
is therefore given by
 (18)
(18)
where
 is the individual’s output in activity
is the individual’s output in activity .
.
Finally the individual’s fitness is

where
 is the individual’s consumption of activity
is the individual’s consumption of activity ,
and
,
and
 is the “utility function”, which is concave in each argument, and satisfies the
“Inada conditions”:
is the “utility function”, which is concave in each argument, and satisfies the
“Inada conditions”: ,
, .
.
Under autarky, we have ,
and the following result holds:
,
and the following result holds:
PROPOSITION 2―Under autarky, all loci are selective. Therefore, in any steady state
such that ,
,
 ,
all individuals are of genotype
,
all individuals are of genotype ,
i.e. the H-allele is fixed at all locations.
,
i.e. the H-allele is fixed at all locations.
Proof―Type
 has a more favorable time budget constraint than type
has a more favorable time budget constraint than type .
Therefore, it achieves a higher fitness. The rest follows from the previous subsection.
Q.E.D.
.
Therefore, it achieves a higher fitness. The rest follows from the previous subsection.
Q.E.D.
Note that the case
 is not of interest: it means that the high allele does not exist at that locus.
is not of interest: it means that the high allele does not exist at that locus.
5. Trade
Let us now look at the trade case. Each good
 is traded at price
is traded at price .
We assume the following normalization for the price vector
.
We assume the following normalization for the price vector

 (19)
(19)
People allocate their time between the various activities so as to maximize their
income ,
,
subject to the time allocation constraint (18). Their demand vector is the one which
maximizes
 subject to their budget constraint:
subject to their budget constraint:

Types with lower incomes must achieve lower fitness and therefore disappear in the long run.
Furthermore,
 must
be monotonic at all loci. The reason is that the vector
must
be monotonic at all loci. The reason is that the vector
 supplied by a geno- type
supplied by a geno- type
 can also be supplied by genotype
can also be supplied by genotype .
On the other hand, all loci need not be selective, as genotypes with fewer H-alleles
at locus
.
On the other hand, all loci need not be selective, as genotypes with fewer H-alleles
at locus
 may achieve the same income as fitter genotypes, by just specializing.
may achieve the same income as fitter genotypes, by just specializing.
Define a long-run equilibrium (LRE), as a stationary state such that the economy is in equilibrium, i.e. each genotype sets its supply and demand as just described, and markets clear for each good. The following proposition generalizes the results derived for the two-loci case in Saint-Paul (2007).
PROPOSITION 3―(i) In any LRE such that ,
,
 ,
a given type only supplies goods corresponding to loci in their genotype where they
have the highest number of
,
a given type only supplies goods corresponding to loci in their genotype where they
have the highest number of
 -alleles:
-alleles:

(ii) In any LRE such that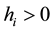 ,
the price vector is
,
the price vector is
 such that
such that
 (20)
(20)
(iii) In any LRE, there exists a locus
 such that
such that ,
i.e. allele H is fixed at locus
,
i.e. allele H is fixed at locus .
.
Proof of (i)―Iterating the mixing property with appropriately chosen parents, one
can easily show that if ,
,
 ,
in steady state there exists a strictly positive supply of genotypes with a arbitrary,
strictly positive number of H-alleles
,
in steady state there exists a strictly positive supply of genotypes with a arbitrary,
strictly positive number of H-alleles
 at each locus
at each locus .
In particular, there exists a strictly positive mass of the best genotype
.
In particular, there exists a strictly positive mass of the best genotype . Next, note
that if
. Next, note
that if ,
then genotype
,
then genotype
 achieves higher fitness than
achieves higher fitness than ,
and hence
,
and hence .
.
Assume there exists a genotype
 such that
such that
 for
for
 such that
such that
 Clearly, the plan
Clearly, the plan
 achieves a strictly higher income level and is feasible (i.e. satisfies (18)) for
achieves a strictly higher income level and is feasible (i.e. satisfies (18)) for .
Consequently,
.
Consequently,
 ,
implying
,
implying .
But, given that
.
But, given that
 is monotonic, Proposition 1, under assumption (ii), would then imply that
is monotonic, Proposition 1, under assumption (ii), would then imply that , which makes
it impossible for
, which makes
it impossible for
 to exist. Consequently, any type
to exist. Consequently, any type
 only supplies goods where it has an
only supplies goods where it has an
 -allele.
-allele.
Proof of (ii)―The price vector defined by (20) is the one which makes type
 indifferent between all activities. Assume there exists an LRE with a different
price vector. Then there exists a pair of goods
indifferent between all activities. Assume there exists an LRE with a different
price vector. Then there exists a pair of goods
 such that
such that
 (21)
(21)
and
 since more income is yielded for type
since more income is yielded for type
 by offering good
by offering good
 than good
than good .
.
Since
 satisfies the Inada conditions, the demand for good
satisfies the Inada conditions, the demand for good
 is strictly positive; since
is strictly positive; since
 does not
does not
supply good ,
there exists
,
there exists
 such that
such that
 and
and .
By virtue of (i),
.
By virtue of (i), .
Further- more,
.
Further- more,
 ,
otherwise
,
otherwise
 would prefer to supply
would prefer to supply
 instead of
instead of
 as well.
as well.
The income of type
 is
is .
The supply vector
.
The supply vector
 is feasible for type
is feasible for type ,
since
,
since
 is more productive than
is more productive than
 at all activities. The supply vector
at all activities. The supply vector
 defined by
defined by ,
,
 ,
,
 ,
,
 ,
,
 also
satisfies (18) for
also
satisfies (18) for .
Therefore,
.
Therefore,

where the last inequality comes from (21). But, this cannot hold since it again
implies
 Consequently, there exists
Consequently, there exists
 such that
such that
 Furthermore, as
Furthermore, as ,
,
iterating Lemma 1 implies that .
Monotonicity of
.
Monotonicity of
 then implies that (ii) in Proposition 1 is satisfied. Consequently,
then implies that (ii) in Proposition 1 is satisfied. Consequently, . But that contradicts
the requirement that
. But that contradicts
the requirement that .
Q.E.D.
.
Q.E.D.
Proof of (iii)―Suppose not; then by iterating the mixing property with appropriately
chosen parents, one can prove that .
But that contradicts (i). Q.E.D.
.
But that contradicts (i). Q.E.D.
The preceding proposition tells us what properties an LRE must necessarily have,
but does not tell us whether an LRE exists and whether, as in the preceding analysis,
one can construct equilibria with a positive level of some
 -alleles.
We now establish a result which tells us that an LRE exists with a strictly positive
proportion of L-alleles, provided these alleles are not too frequent.
-alleles.
We now establish a result which tells us that an LRE exists with a strictly positive
proportion of L-alleles, provided these alleles are not too frequent.
To do so, for any subset
 of
of
 we define
we define
 as
as

 is the set of all
is the set of all
genotypes such that their loci saturated with
 -alleles
(which define the activities at which they can possibly specialize) are all in
-alleles
(which define the activities at which they can possibly specialize) are all in .
.
PROPOSITION 4―Let
 be the inverse demand function for the fitness maximization problem of an individual
with income R facing price vector
be the inverse demand function for the fitness maximization problem of an individual
with income R facing price vector

 Let
Let

Then there exists an LRE with a distribution
 of genotypes if and only if this distribution satisfies the following property:
of genotypes if and only if this distribution satisfies the following property:
 (22)
(22)
Proof―We first prove that this condition is necessary. The RHS of (22) is the total
time supplied by genotypes in
 (relative to the population's total time). Proposition 3, (i) implies that it must
be allocated among goods
(relative to the population's total time). Proposition 3, (i) implies that it must
be allocated among goods
 such that
such that ,
i.e. among goods in
,
i.e. among goods in .
It also implies that in any candidate equilibrium, income per
.
It also implies that in any candidate equilibrium, income per
capita (equal to the income of any genotype) must be equal to
 Thus,
Thus,
 is
the per capita amount of good
is
the per capita amount of good
 consumed and produced in any candidate equilibrium. The LHS of (22) is therefore
the total time input needed to produce all the goods in
consumed and produced in any candidate equilibrium. The LHS of (22) is therefore
the total time input needed to produce all the goods in .
It must be greater than or equal to its RHS, since genotypes in
.
It must be greater than or equal to its RHS, since genotypes in
 cannot produce any other good. Otherwise, supply would exceed demand. Note that
(22) applied to
cannot produce any other good. Otherwise, supply would exceed demand. Note that
(22) applied to

implies that one
 -allele
is fixed (the RHS is then the total supply of all genotypes
-allele
is fixed (the RHS is then the total supply of all genotypes
 such that
such that ).
Also, (22) applied to
).
Also, (22) applied to
 boils down to Walras’ law, since it is equivalent to
boils down to Walras’ law, since it is equivalent to ,
and by Walras law
,
and by Walras law .
.
Let us now prove sufficiency. In order to do so, we construct a set of functions ,
representing the share of time of genotype
,
representing the share of time of genotype
 devoted to activity
devoted to activity ,
such that:
,
such that:
 (23)
(23)
 (24)
(24)
 (25)
(25)

6One can trivially check that such an allocation exists, since one
 -allele
is fixed, all genotypes have at least one locus where
-allele
is fixed, all genotypes have at least one locus where .
.
If we are able to construct such functions, then this is indeed an equilibrium,
since supply equals demand for all goods, and since the price vector in (20) implies
that a genotype is indifferent between supplying all the goods at which it has

 -alleles.
-alleles.
To construct the ,
we use the following algorithm. We start from any arbitrary allocation
,
we use the following algorithm. We start from any arbitrary allocation
 satisfying (23) and (25). This defines the initial stage.6 Then we move
from stage
satisfying (23) and (25). This defines the initial stage.6 Then we move
from stage
 to stage
to stage
 as follows. At the beginning of stage
as follows. At the beginning of stage ,
the set
,
the set
 can be partitioned into three subsets:
can be partitioned into three subsets:

That is, those goods for which supply equals demand, those for which there is excess
demand, and those for which there is excess supply. Note that since ,
,
 is
empty if and only if
is
empty if and only if
 is empty. If
is empty. If ,
then we have an equilibrium, and the algorithm stops.
,
then we have an equilibrium, and the algorithm stops.
Assume therefore that it is not the case. Then neither
 nor
nor
 is empty. We now distinguish two cases.
is empty. We now distinguish two cases.
Case A. Assume there exists a partition
 of
of
 such that
such that

and:
 (P)
(P)
That is, people who do produce goods in
 cannot produce goods in
cannot produce goods in .
.
For any good ,
let
,
let

Clearly, one has ,
for all
,
for all .
We then have
.
We then have

This strict inequality comes from the fact that
 and from the fact that
and from the fact that
 is non-empty.
is non-empty.
Furthermore,

This is because if
 and
and ,
then
,
then ,
implying
,
implying .
Therefore
.
Therefore
 .
.
Interverting, we get

Now, note that,
 ,
, :
if I produce one good in
:
if I produce one good in ,
all my loci with
,
all my loci with

 -alleles
are also in
-alleles
are also in .
Consequently,
.
Consequently,

which clearly violates assumption (22). Case A is therefore ruled out.
Case B. Assume then that there exists no such partition. We can construct a chain
of
 goods
goods
 such that the following property holds:
such that the following property holds:
PROPERTY Q:
(a)

(b)

(c) ,
for
,
for

(d) ,
,
 ,
,
 and
and

To construct such a chain, proceed as follows. We will write
 if (
if ( ,
, and
and ).
In this case, we call
).
In this case, we call
 the set of genotypes that satisfy this property:
the set of genotypes that satisfy this property: .
Property (d) implies that the chain we want to construct is such that
.
Property (d) implies that the chain we want to construct is such that

Start from a set .
As property (P) is violated, there exists
.
As property (P) is violated, there exists ,
,
 ,
such that
,
such that

If ,
stop the procedure there, and take
,
stop the procedure there, and take .
.
If not, then .
Add
.
Add
 to
to :
: .
Since
.
Since ,
it must be that
,
it must be that ,
, .
Use again the fact that (P) is violated. There exists
.
Use again the fact that (P) is violated. There exists ,
,
 ,
such that
,
such that .
Given that either
.
Given that either
 or
or ,
there exists a chain of length
,
there exists a chain of length ,
,
 such
that
such
that ,
,
 ,
,
 ,
and
,
and .
If
.
If ,
we use that chain and stop the procedure. Otherwise, we add
,
we use that chain and stop the procedure. Otherwise, we add
 to
to ,
and iterate again.
,
and iterate again.
More generally, at each iteration ,
there is a set
,
there is a set
 such that
such that ,
and such that for all
,
and such that for all ,
there exists a chain
,
there exists a chain
 such that
such that ,
,
 ,
and
,
and
 (The chain property)
(The chain property) Because (P) is violated, there exists
Because (P) is violated, there exists
 and
and ,
such that
,
such that .
Let
.
Let
 be the chain corresponding to
be the chain corresponding to .
If
.
If ,
we use the chain
,
we use the chain
 and stop the procedure. If
and stop the procedure. If ,
we use
,
we use
 and iterate the procedure. As the new member
and iterate the procedure. As the new member
 is connected to
is connected to
 via the chain
via the chain ,
,
 still
satisfies the chain property. As
still
satisfies the chain property. As ,
it also satisfies
,
it also satisfies .
As the number of elements in
.
As the number of elements in
 goes up by one unit at each iteration, one must find an
goes up by one unit at each iteration, one must find an
 in
in
 a finite number of iterations.
a finite number of iterations.
Next, we can use such a chain to construct a new allocation of labor for stage .
Let
.
Let

and

Define the new allocation as follows:

The new allocation clearly still satisfies (23) (as ),
and (25) (as
),
and (25) (as
 ).
Futhermore, one has
).
Futhermore, one has ,
as
,
as .
Finally, for
.
Finally, for ,
, .
Hence, all markets that were in equilibrium remain so. Furthermore, as
.
Hence, all markets that were in equilibrium remain so. Furthermore, as , market
, market
 weakly remains in excess supply, and similarly market
weakly remains in excess supply, and similarly market
 weakly remains in excess demand. Therefore:
weakly remains in excess demand. Therefore:

Finally, we note that either
(i)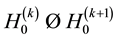 ,
which will be true provided
,
which will be true provided
 or
or
 .
In such cases the new allocation restores equilibrium in market
.
In such cases the new allocation restores equilibrium in market
 (resp.
(resp. ).
).
(ii) Or, the chain
 and its associated chain of genotypes
and its associated chain of genotypes
 no longer satisfy Q; that is the case if
no longer satisfy Q; that is the case if
 for some
for some ,
in which case
,
in which case .
In such a case, we have constructed a new allocation such that
.
In such a case, we have constructed a new allocation such that
 has increased by at least one unit, and which satisfies (23) and (25).
has increased by at least one unit, and which satisfies (23) and (25).
Thus, at each stage, the quantity
 strictly increases. As it is bounded, the proce- dure cannot go on forever, and
the only case in which one cannot iterate it is if
strictly increases. As it is bounded, the proce- dure cannot go on forever, and
the only case in which one cannot iterate it is if .
This proves
.
This proves
the existence of equilibrium. Q.E.D.
Clearly, conditions (22) are pretty stringent, so that it is not straightforward to construct an equilibrium.
However for
 close enough to 1, i.e.
close enough to 1, i.e.
 small
enough when
small
enough when ,
they are clearly satisfied, since
,
they are clearly satisfied, since
 appears on the RHS only for
appears on the RHS only for ,
in which case (22) is always satisfied with equality, due to Walras’ law:
,
in which case (22) is always satisfied with equality, due to Walras’ law: . Therefore there
always exist equilibria with a strictly positive fraction of genotypes with
. Therefore there
always exist equilibria with a strictly positive fraction of genotypes with
 -alleles,
provided this fraction is small enough.
-alleles,
provided this fraction is small enough.
Note that the greater the number of loci, the greater the number of conditions that
must hold. Intuitively, it suggests that the equilibrium fraction of
 -alleles
must become smaller. Intuitively, if the initial distribution of alleles in the
population is such that (22) is violated, we expect a number of H-alleles to eventually
become fixed, which is equivalent to a reduction in
-alleles
must become smaller. Intuitively, if the initial distribution of alleles in the
population is such that (22) is violated, we expect a number of H-alleles to eventually
become fixed, which is equivalent to a reduction in .
The process would continue until
.
The process would continue until
 is small enough for the number of relevant activities not to be too large, so that
(22) holds.
is small enough for the number of relevant activities not to be too large, so that
(22) holds.
References
- Cavalli-Sforza, L. and Feldman, M.W. (1981) Cultural Transmission and Evolution. Princeton University Press, Princeton.
- Lumsden, C.J. and Wilson, E.O. (1981) Genes, Mind, and Culture: The Coevolutionary Process. Harvard University Press, Cambridge.
- Gintis, H. (2003) The Hitchhiker’s Guide to Altruism: Gene-Culture Coevolution and the Internalization of Norms. Journal of Theoretical Biology, 220, 407-418. http://dx.doi.org/10.1006/jtbi.2003.3104
- Boyd, R. and Richerson, P.J. (1985) Culture and the Evolutionary Process. University of Chicago Press, Chicago.
- Hirshleifer, J. (1977) Economics from a Biological Viewpoint. Journal of Law and Economics, 20, 1-52. http://dx.doi.org/10.1086/466891
- Robson, A. (2001) The Biological Basis of Economic Behavior. Journal of Economic Literature, 29, 11-33. http://dx.doi.org/10.1257/jel.39.1.11
- Seabright, P. (2004) The Company of Strangers: A Natural History of Economic Life. Princeton University Press, Princeton.
- Saint-Paul, G. (2007) On Market Forces and Human Evolution. Journal of Theoretical Biology, 247, 397-412. http://dx.doi.org/10.1016/j.jtbi.2007.03.021
- Horan, R.D., Bulte, E.H. and Shogren, J.F. (2009) How Trade Saved Humanity from Biological Extinction: An Economic Theory of Neanderthal Extinction. Journal of Economic Behavior and Organization, 58, 1-29. http://dx.doi.org/10.1016/j.jebo.2004.03.009
- Hammerstein, P., Ed. (2003) Genetic and Cultural Evolution of Cooperation. MIT Press, Cambridge.
- Bowles, S. and Hammerstein, P. (2003) Does Market Theory Apply to Biology. In Hammerstein, P., Ed., Genetic and Cultural Evolution of Cooperation, MIT Press, Cambridge, 153-165.
NOTES

1In this line of work, grouyp selection often plays an important role. See for example Cavalli-Sforza and Feldman [1] ; Lumsden and Wilson [2] ; Gintis [3] , Boyd and Richerson [4] .
2Interesting surveys on interactions between the economic and biological spheres include Hirshleifer [5] , Robson [6] , and Seabright [7] .
3I addition to this, the most closely related paper is Horan et al. [9] . A related literature (see Hammerstein [10] , and in particular Bowles and Hammerstein [11] ), studies the rise of markets and specialization in animal societies, but does not draw this paper’s implications for the gene pool.
4If ,
we can write down the same steps using the smallest value of
,
we can write down the same steps using the smallest value of
 such that
such that
 as a benchmark.
as a benchmark.
5The only other possibility is to only have genotypes such
 and such that
and such that ,
but random mating and mixing imply that they will produce offsprings such that
,
but random mating and mixing imply that they will produce offsprings such that .
.