Natural Science
Vol.5 No.6A(2013), Article ID:33020,9 pages DOI:10.4236/ns.2013.56A001
Isolation of enriched-yielders and fed-batch production of alkaline protease from the newly isolated Bacillus sp. BHA*
![]()
1Department of Anesthesiology, Medical College of Wisconsin, Milwaukee, USA; #Corresponding Author: bagarwal@mcw.edu, bhawanaa@gmail.com
2Department of Biochemical Engineering and Food Technology, Harcourt Butler Technological Institute, Nawabganj, India
Copyright © 2013 Bhawana Agarwal, Brajesh S. Katiyar. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received 25 April 2013; revised 26 May 2013; accepted 3 June 2013
Keywords: Industrial Enzymes; Alkaline Protease; Feedback Repression; Batch Culture; Fed-Batch Process
ABSTRACT
An alkalophilc and thermophilic Bacillus sp. BHA that produced a thermostable alkaline protease was isolated from decaying protein substrates. The isolate was found to grow in pH range 7 - 11 with an optimum pH 9.0 and temperature up to 55˚C. The activity of alkaline protease of Bacillus sp. BHA (68.98 APU/ml) was found higher than the standard strains of Bacillus amyloliquefaciens MTCC 610 (8.98 APU/ml) and Bacillus subtilis MTCC 8349 (12.14 APU/ml, used in this study, and was comparable (68.98 APU/ml, equivalent to 30.38 APU/mg) to the activity of the commercially produced standard protease procured from Novo Nordisk, Denmark (30.35 APU/mg). Hence, the proteolytic activity produced by this isolate was further investigated in batch and fed-batch process. Sucrose was the best carbon source for the production of protease activity by that isolate. Different organic nitrogen sources (casein, peptone and beef extract) at 1% (w/v) with varying levels of sucrose (1% - 4% w/v) initially repress enzyme synthesis. The duration and extent of repression decreased with increased concentration of sucrose. Maximum protease activity was found in basal medium with 4% (w/v) sucrose and 1% (w/v) yeast extract. Yeast-extract was thought to be an inducer of enzyme synthesis. Further, the basal medium was unique with respect to the enzyme production, as protease production was growth associated with the peak enzyme production being detected at the time of maximum growth. Interestingly, a rise in 34.2% (104.86 APU/ml) of protease activity was detected at incubation temperature of 50˚C and when culture filtrate was assayed at 60˚C, signifying a high temperature stability of the produced protease by this isolate. Additional studies on the enzyme characterization were resulted in recognition of highly significant properties of the activity towards casein at pH 9.0 and stability at high temperature with retention of 96% the enzyme activity at 60˚C. The parametric study under feed intervals had enabled improvement in the maximum protease activities attainable from batch cultures in excess of 21.78% and 26.32% via two feeding strategies. A small continual increase in enzyme activity (132.46 APU/ml during 24 h - 120 h) and enhancement in protease production in excess of 36.84% was observed by fed-batch process than the batch experiment.
1. INTRODUCTION
The global demand for industrial enzymes is estimated to be approximately $2 billion. More than 90% of the sales of industrial enzymes currently used however come from fewer than 30 enzymes [1]. Protease constitutes one of the most significant groups of industrially important enzymes accounting for 60% of the total enzymes market [2]. They form a very large and complex group of enzymes, which differ in properties such as substrate specificity, active site, catalytic mechanism, pH and temperature optima and stability profile [3]. Enzymes in laundry and cleaning products account for 40% of global industrial enzyme market (they are also used in food processing, medical diagnostics and textile processing) of these alkaline proteases used primarily as detergent additive represents about 60% of the global detergent enzyme market for enzyme accounting for approximately 19% of total enzyme demand and more than 25% of industrial enzymes sales [4-6]. With the advent of a wide variety of microbial proteases, and low cost production by fermentation processes of high purity from microbial sources, microbial proteolytic enzymes have replaced the traditional ones form animal and plant sources [7], with bacterial proteases top the list of various industrial enzymes in world wide production and sales.
Alkaline proteases (EC 3.4.21.14) are an important group of enzymes that occupies a large share of the commodity enzyme market and are mainly used in detergent and also in industries like leather, food, photography, pharmaceuticals etc. [8]. Microbial alkaline proteases are reported to share the following properties-A serine residue at the active site, esterolytic activity, alkaline pH optima and sensitivity to organophosphorous reagents such as phenyl-methyl sulphonyl fluoride (PMSF) or Di-isopropyl fluorophosphates (DIFP), generally not inhibited by EDTA. Furthermore, they are not activated by metal ions or reducing agents. The alkaline serine proteases are specific for aromatic or hydrophobic residues such as tyrosine phenylalanine or leucine. These enzymes have requirement for Ca2+ for their stability and optimum activity under alkaline conditions and are found to be functional in detergent components.
Here, we report the isolation of new Bacillus sp. BHA for high potentiality of alkaline protease production by enriched-growth culture method. The batch culture of alkaline protease production by this isolate was found under the control of repression, wherein, in the presence of a readily metabolizable carbon source growth is normal but exo-protease synthesis is switched off. Thus, for the sequential study of stimulatory and inhibitory/repression effect of key culture parameters and to wear-off the time taken for the repression, different nitrogen sources with varying concentrations of carbon sources via different feed compositions and feed rates under fed-batch culture were investigated. The fed-batch method was further optimized for enhanced protease production. The optimized growth of Bacillus sp. BHA under alkaline and high temperature favored the alkalophilic and thermostable characterization of the produced protease.
2. MATERIALS AND METHODS
2.1. Isolation and Identification of Pure-Cultures
Alkaline protease producing strains of Bacillus were isolated from decaying protein substrates (Nutrinugget, Cottage cheese, Gram flour and Pulse) by a stepwise screening using casein media. Following media were used during the course of this investigation (per l): media #1 (M1); Casein broth media (consisted of): Casein 40 g, MgSO4·7H2O 2 g, CaCl2·2H2O 100 μg, Trace element solution 1 ml (per l: ZnSO4·7H2O 8800 mg, FeCl3·7H2O 970 mg, CuSo4·5H2O 270 mg, MnCl2·4H2O 72 mg), pH 10.0, adjusted with NaOH. Medium #2 (M2); M1 with 2% agar, pH 10.0 adjusted with NaOH. Media (M1 and M2) were sterilized at 15 psi for 15 min.
2.2. Isolation of Enriched-Yielders by Growth Cycles
Small amounts of the decaying protein substrates were suspended in 10 ml of sterile distilled water and mixed thoroughly. These suspensions were allowed to settle down for 30 min to obtain a clear suspension. 0.1 ml of 1:100 dilution of the clear suspensions from different samples were plated evenly on the plates of the medium #2 (M2) by spread plate technique under sterile conditions. Plates were incubated at 50˚C for 72 h to allow the growth of each viable organism into a colony. The isolated colonies were selected on the basis of zone of caseinolysis and each selected colony was further subjected to shake flask experiments at 50˚C and 200 rpm in liquid-casein medium (M1) for 24 h. At the end of three successive growth cycles, survived cultures were plated on medium #2 (M2) and were further selected on the basis of largest diameter of caseinolytic zone observed.
2.3. Selection of Strains
Four bacterial cultures, two suitably selected isolated Bacillus species and two other standard strains of Bacillus amyloliquefaciens MTCC 610 and Bacillus subtilis MTCC 8349 procured from Institute of Microbial Technology, Chandigarh were examined for highest producers of alkaline protease. During the course of this investigation screening media consisted of (g/l) MgSO4·7H2O 1, CaCl2·2H2O 0.5, yeast extract 10 and supplemented with 4% (w/v) of different carbon sources e.g. starch, sucrose, maltose, glycerol and citric acid were used.
2.4. Selection of Suitable Fermentation Media
The basal salt solution used was consisted of (g/l): MgSO4·7H2O 1, CaCl2·2H2O 0.5. This was supplemented with different nitrogen sources in presence of varying concentrations of the selected carbon source. Different basal media containing either of casein, peptone or beef extract, each at 1% level was constituted different media. Separate controls of these nitrogen sources 0.5% (w/v), in combination of 0.5% (w/v) yeast extract with 2% (w/v) sucrose were also prepared.
2.5. Cultural Conditions in Batch-Process
Fermentation experiments were carried out in 100 ml each of the above described medium in 250 ml Erlenmeyer flasks. (5% (w/v) and 24 h old raised inoculum (inoculum medium composition (g/l): glucose 10, yeast extract 10, peptone 5, NaCl 3, pH 7 - 8) at 37˚C and 200 rpm was used to initiate the growth of the cultures. The inoculated flasks were kept in incubator shaker at 200 rpm for 120 h at 37˚C. Samples were taken at every 24 h interval started from 0 h, centrifuged at 2500 rpm for 10 min and supernatant were assayed for enzyme activity. Some experiments of batch process were also carried out at incubation temperature of 50˚C and supernatant were assayed at 60˚C for enzyme activity.
2.6. Determination of pH and Thermal Activity
Optimum pH and temperature required for enzyme activity was determined by carrying out the estimations in casein solution of different pH buffers (0.2 M phosphate buffer for pH 6 - 8 and 0.2 M glucine-NaOH buffer for pH 9 - 11) and residual enzyme activity was measured.
2.7. Determination of pH and Thermal Stability
0.5 ml of culture filtrate was pre-incubated with 2 ml 0.2 M glycine-NaOH buffer of different pH (6 - 10) at 60˚C for 10 min, and in other experiments at various temperature range of 30˚C - 90˚C for 10 min. After 10 min the reaction was terminated by keeping the test tubes in ice-bath. The enzymatic assay was carried out by taking 0.5 ml of these samples in 0.2 M glycine-NaOH buffer at pH 9.0 at 37˚C and residual enzyme activity was measured.
2.8. Studies on Feed Rates and Feeding Composition of Carbon and Nitrogen Sources in Fed-Batch Process
Four simultaneous fed-batch experiments in two set ups with different intermittent feeding rates and feeding solutions of different carbon and nitrogen compositions were carried out. Set up 1: the feeding solution consisted of (g/l); sucrose 40, yeast extract 10, was fed intermittently, at the rates of 6 ml and 12 ml per feed at the interval of 3 h with the total numbers of 3 feeds per day. Set up 2: the feeding solution consisted of (g/l); sucrose 80, yeast extract 20, was fed intermittently at the rates of 9 ml and 12 ml per feed at the interval of 3 h with the total numbers of 3 feeds per day.
2.9. Studies on Feed Intervals of Whole Medium in Fed-Batch Process
Experiments were carried out with 50 ml of initial volume (batch mode) to final working volume of 150 ml (fed-batch mode) in Erlenmeyer flasks (500 ml) using two different strategies of feeding. The medium used for these studies was comprised of (g/l): MgSO4·7H2O 1, CaCl2·2H2O 0.5, sucrose 40, yeast extract 10. The duration of each batch operation preceding a fed-batch operation was 24 h. 100 ml of the feeding solution was allowed to feed intermittently at different rates with a volumetric feed of 40 ml per day in both the two strategies. Strategy 1: Two volumetric feed of 40 ml per day was distributed in two feeds with the feeding volume of 20 ml per feed at the interval of 7 h. Strategy 2: the volumetric feed of 40 ml per day was distributed in four feeds at the intervals of 2 h.
2.10. Cultural Conditions in Fed-Batch Process
Erlenmeyer flasks (500 ml) having 50 ml of the medium consisted of basal salt solution (g/l); MgSO4·7H2O 1, CaCl2·2H2O 0.5 with sucrose 20 and yeast extract 5 was used for cultivation of cells in batch culture prior to initiation of fed-batch operation. 5% (w/v) inoculum incubated at 37˚C, 200 rpm for 48 h was used. Fed-batch experiments were started from this initial volume (50 ml) to final variable volumes by intermittent feeding of the solution as described in each set up, under similar cultural conditions. Sampling was done at every 24 h intervals, started from 0 h, centrifuged at 2500 rpm for 10 min and the supernatant so obtained was used for the assay.
2.11. Assay Method for Alkaline Protease
The activity of alkaline protease was estimated by using the modified Anson’s method [9]. The method was based on liberation of tyrosine in trichloroacetic acid (TCA) filtrate and was colorimertically estimated using Folin-ciocalteau reagent. 2 ml aliquot of casein solution (0.5% w/v) was added to a test tube and equilibrated in 37˚C water bath for 5 min. 0.5 ml of properly diluted cultural filtrate was added to the aliquot and was incubated at 37˚C for 10 min. At the end of the incubation period the reaction is terminated by adding 4 ml of 5% (w/v) TCA. For the blank solution 0.5 ml of buffer was substituted for the cultural filtrate. A set of three tubes with 0.5 ml supernatant from blank, control and sample were prepared with the addition of 4 ml of 0.4 M Na2CO3 solution and 0.5 ml two fold diluted Folin-ciocalteau reagent. After 30 min the blue color developed was read at 660 nm on UV visible spectrophotometer (Model UV- 1601) against blank. The values for the controls were subtracted from those of the sample. Enzyme activity in culture filtrate was calculated from the formula: ΔA × D/t × V units/ml, where, ΔA = concentration difference of tyrosine corresponding to difference in absorbance of sample and control. D = dilution factor i.e. factor by which culture filtrate was suitably diluted with buffer. T = timeperiod of incubation (10 min). V = volume of culture filtrate taken for incubation (0.5 ml). One unit of enzyme activity was defined as the amount of enzyme which catalyses the release of Folin positive amino acids equivalent to 1 μmoles of tyrosine from casein per min. under the assay conditions used. Activity was expressed as units per ml of culture filtrate.
2.12. Assay for Protein Content
Protein content was determined by Folin-Lowry method [10] using BSA as standard. Protein reacts with Folin-ciocalteau reagent to give the colored complex due to the reduction of the phosphomolybdate by tyrosine and tryptophone present in the protein.
2.13. Determination of Biomass
30 ml of the fermented broth was first centrifuged at 2500 rpm for 15 min and the whole biomass so obtained was used for plotting standard curve for dry cell mass vs. optical density. The whole cell mass was suspended in 10 ml of distilled water. 90% (9 ml) of that was then dried over night in pre-weighed dish at 50˚C. Different solutions were prepared from rest of 10% (1 ml) cell suspension and optical density in each dilution was measured on spectrophotometer at 600 nm. Dry weight of cells in each dilution was calculated with reference to the dry weight of 90% cell suspension. A standard curve was plotted between dry weights against O.D and the curve was used to determine the dry weight of the unknown cell suspensions.
3. RESULTS AND DISCUSSION
3.1. Isolation and Screening of Bacillus sp.: Development of Enriched-Yielders by Growth Cycles
The search for Bacillus species of high potentiality for alkaline protease production had resulted in development of an isolation method by following an extensive screening of decaying protein substrates, as described in materials and methods. It was considered that only those cultures that had the ability to survive in the presence of 4% (w/v) casein (pH 10.0) would yield high producers of alkaline protease, while relatively poor producers would almost be eliminated through the growth cycles. The survived cultures were then subjected to caseinolysis by plating on casein-agar medium (M2). The data were separately collected for each four sources (Table 1) and were further used for activity screening in shake-flask experiments. From Table 1, it was born out that in subsequent growth cycle treatments number of colonies decreased while percentage of enriched yielders increased. Further, the cultures were selected on the basis of the largest diameter of zone of caseinolysis observed. Every isolate was a single representative of the four samples taken and were designated as isolate #1-4. After qualitative determination on M2 as zone of clearance, further quantitative estimation of alkaline protease production was restored to shake-flask screening of the 4 isolates in M1 medium. Out of the four isolated cultures isolate #1 and #2 afforded comparatively high production profiles under shake-flask level and were further subjected to selection with other standard strains.
3.2. Comparison of Isolated and Standard Cultures with Different Carbon Sources
From the comparative results of isolated and selected
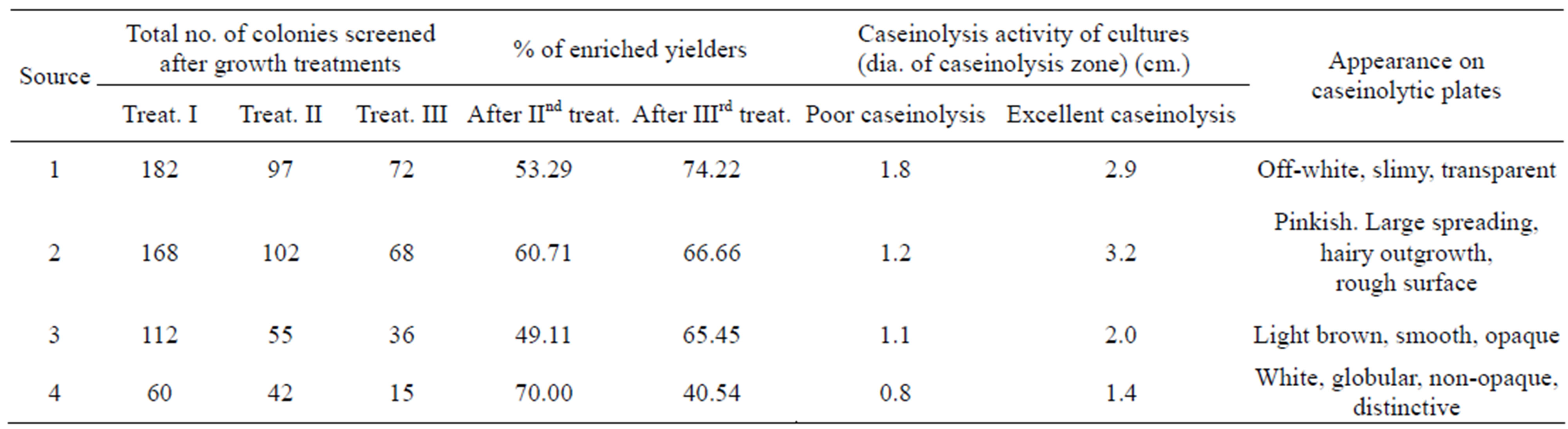
Table 1. Summary of the development of enriched-yielders. The isolated colonies from the degraded protein samples were picked and allowed to grow in shake-culture at 50˚C in liquid casein medium (M1). The survived cultures after three such successive treatments of growth at 50˚C were plated on M1 + agar medium (M2) for 24 h and further selected based on the diameter of zone of caseinolysis. Poor caseinolysis; it shows the smallest diameter of caseinolysis zone observed on casein-agar plates. Excellent caseinolysis; it shows the largest diameter of caseinolytic zone observed on casein-agar plates. They were also characterized based on their appearance on the casein-agar plates.
strains for highest potentiality of alkaline protease producers, isolate #2 showed the highest production profile in all the carbon sources tested; citric acid (6.05 APU/ml), glycerol (8.28 APU/ml), maltose (24.84 APU/ml), and starch (49.67 APU/ml), with the maximum activity in sucrose (68.98 APU/ml) along with the initiation of protease production after 24 h of growth. For isolate #1 starch was found to be the best carbon source with highest activities of (22.0 APU/ml) after 96 h of incubation, while still lower activities were observed by B. amyloliquefaciens MTCC 610 was same (8.28 APU/ml) in different media (except with glycerol of 2.8 APU/ml) with the only difference of peak enzyme production at different hours of incubation. The only utilization of citric acid and glycerol were shown by B. subtilis MTCC 8349 with still lower activities (12.14 APU/ml) and (11.03 APU/ml), respectively at 48 h of incubation. Based on these experiments it was found that isolate #2 gave the best production profile and hence was selected for further fermentation studies under batch and fed-batch processes.
3.3. Identification of Selected Isolate #2
Based on the general and growth characteristics hints given in Bergey’s manual of determinative bacteriology [11], the isolate #2 was appeared as the strain of Bacillus and named as Bacillus sp. BHA. A 5 ml (4.25 × 108 cells/ml) aliquot per 100 ml production medium was served as the inoculums.
3.4. Fermentation Studies in Batch-Process: Effect of Different Nitrogen Sources with Varying Concentrations of Sucrose
Organic nitrogen sources were reported as better producers of alkaline protease than inorganic nitrogen sources [12,13] as was tested here by lesser activities observed with NH4Cl (5.52 APU/ml) and (NH4)H2PO4 (16.56 APU/ml) than yeast extract (68.98 APU/ml) by Bacillus sp. BHA. All these nitrogen sources were taken at 1% (w/v) level. To carry out these studies different organic nitrogen sources (casein, peptone and beef extract) alone and in combination with yeast extract were prepared with various concentrations of sucrose in basal salt solution (as given in materials and methods). With casein at 1% (w/v) level and 1% and 2% of sucrose level, the repression of enzyme activity for up to 72 h was observed, however, with 3% and 4% sucrose level the enzyme activities reaches to plateau after 96 h and 72 h, respectively. With 1% peptone and 1% sucrose there was a severe repression in enzyme activity for up to 120 h of incubation. However, at 2% and 3% sucrose levels there was a large increase in enzyme activity when incubation period was extended from 48 to 120 h. At 4% sucrose level enzyme activity started increasing from 72 h and reached maximum in 120 h. Similarly, with beef extract, there was the repression in enzyme synthesis at 1% sucrose level whereas an exceptional rise in enzyme activities was observed after 96 h of incubation at higher level of sucrose. In general it can be inferred that when sucrose was present in the medium in amounts ranging from 1% to 4% (w/v), protease synthesis was repressed initially. The extent of repression as well as the duration of repression depends on the concentration of carbon source and both decreased with increased concentration of sucrose. Previous repression studies were made with 1% (w/v) of NH4Cl, corn steep liquor (CSL) and cotton seed meal (CSM) and resulted in repression when carbon source was present in media ranging from 1% to 4% (w/v). Further, alkaline protease production in basal medium with casein, peptone and beef extract (each at 1% w/v) were compared with simultaneous controls made by substituting 0.5% (w/v) of these nitrogen sources with yeast extract 0.5% (w/v) along with 2% (w/v) sucrose. From the results so obtained, yeast extract increased the total enzyme activity after 96 h in each case. Higher activities in controls were attributed to either relieved repression because of the low level of nitrogen sources or because of some inducing effect of yeast extract. Finally, as approximately same enzyme activities were obtained in all controls, it was finally concluded that irrespective of other nitrogen sources taken these levels of activities were reached only because of the inducing effect of yeast extract on enzyme production that could be attributed to the presence of some peptides and mixture of amino acids. Further, in basal medium with sucrose (4% w/v) and yeast extract at 1% (w/v) the highest enzyme activity (68.98 APU/ml) was obtained. The protease production was growth associated, with the peak enzyme production being detected at the stage of maximum growth. The production of protease in Bacillus concomitant with growth has been rarely reported [14,15]. Further, fermentation studies were carried out in fed-batch process.
3.5. Effect of pH and Temperature on Enzyme Activity
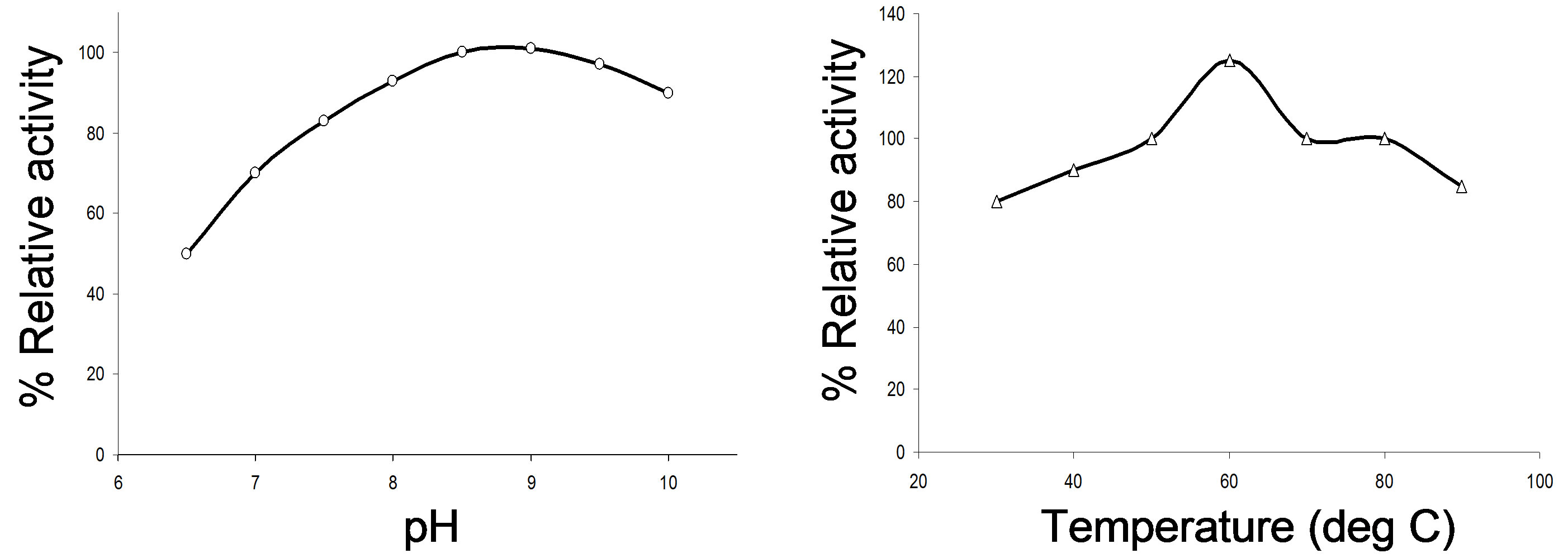
The enzyme exhibited its activity predominantly in the higher pH range (pH 8 - 10) with an optimum pH activity of pH 9.0. At pH range 8.3 - 9.4 about 98% was protease activity was retained (Figure 1(a)). The optimum temperature for enzyme activity was found to be 60˚C as maximum of 133.33% enzyme activity was observed at this temperature, However, it was 100% (or more) as active as at 37˚C at other higher temperatures (50˚C - 80˚C) also (Figure 1(b)).
3.6. Effect of pH and Temperature on Enzyme Stability
In general higher activities were observed when the enzyme was stored at high alkaline pH (8.6 - 9.3). A
 (a)
(a)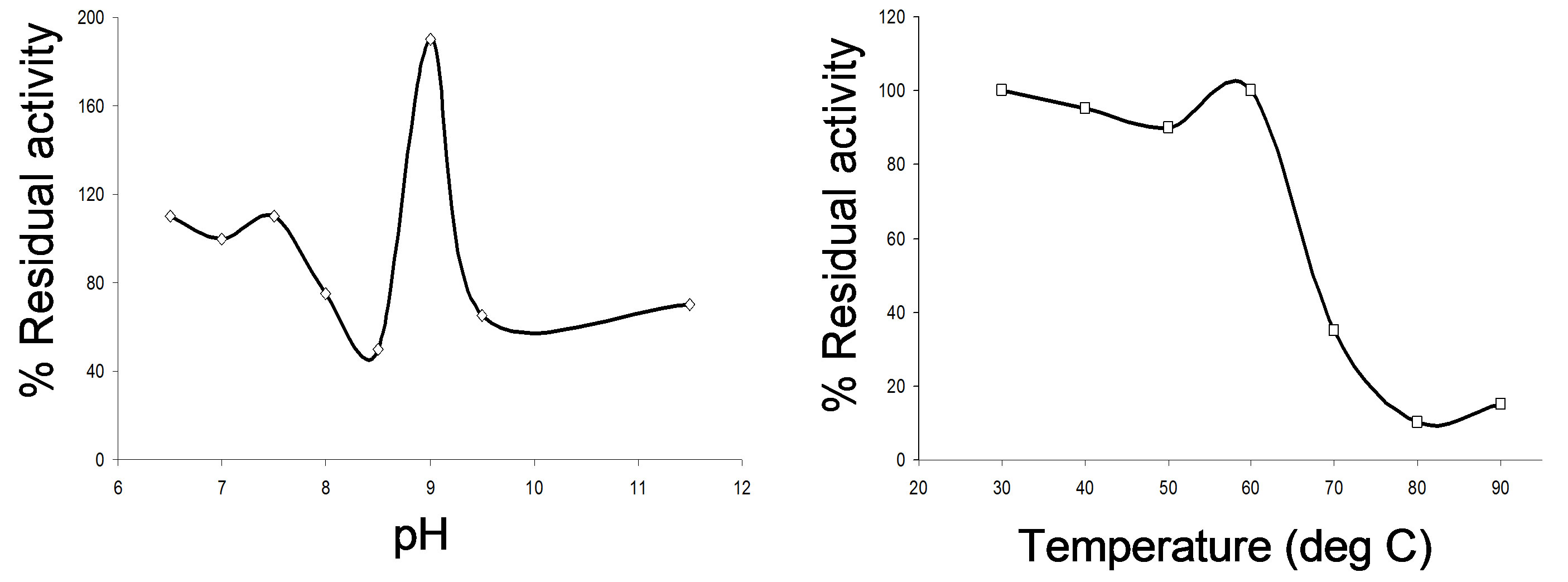 (b)
(b)
Figure 1. Characterization of the alkaline protease produced from Bacillus sp. BHA; effect of pH and temperature on the alkaline protease activity and stability. (a) Effect of pH on the enzyme activity; the relative activities (%) were calculated with reference to the maximal activity found; (b) Effect of temperature on the enzyme activity; enzymatic activities were expressed as % of the activity at 37˚C, pH 10.0; (c) Effect of pH on enzyme stability; stability at pH 7 was taken as 100%; (d) Effect of temperature on the enzyme stability; enzymatic activities after heating at 30˚C for 10 min was taken as 100%.
large increase in activity (86%) was observed when the enzyme was preincubated in buffer oh pH 9.0 (Figure 1(c)). The enzyme was quite stable at temperature range 30˚C - 60˚C and retains 96% activity at 60˚C. A sharp loss of activity was observed at 70˚C (Figure 1(d)).
3.7. Fermentation Studies in Fed-Batch Process: Effect of Different Feed Rates of Carbon and Nitrogen Sources
Starting with the nutrient medium used in batch suspension cultures of Bacillus sp. BHA, the effect of other cultural parameters including the feed rate of carbon and nitrogen sources and feed intervals of whole medium on protease production were investigated in fed-batch process. These experiments were conducted by intermittent feeding of nutrients solution during the course of an otherwise batch operation.
To investigate the repression/induction of protease synthesis and effect of the carbon source (sucrose) on protease production, fed-batch experiments with different feed rates and feeding compositions (as described in material and methods) were conducted. The scheme (Table 2) and results (Tables 3 and 4) show that the increased feed concentration of yeast extract (YE) increased the bulk protease activity while, the specific protease activity (SE) were decreased in each of the different feed rate employed that could be attributed to feed-back inhibition by some of the amino acids generated after proteolysis caused by the synthesized protease. At higher YE concentration in the feed, sucrose utilization was increasingly directed towards cell growth (and less towards protease synthesis), which was shown by increased protease activity but the protease yield based on YE fed (E/YEf) decreased with such an increase. Unlike others the forth fed-batch experiment was marked by a steady decrease in protease activity and was attributed to further repression by excess sucrose fed, which was also indicated by the decreased protease yield based on the sucrose fed (E/Sf).
3.8. Effect of Feed Intervals of Whole Medium
For substantial enhancement of protease synthesis and to optimize the feed concentration (as described in material and methods), the fed-batch experiments were conducted with the initial and final working volume of each
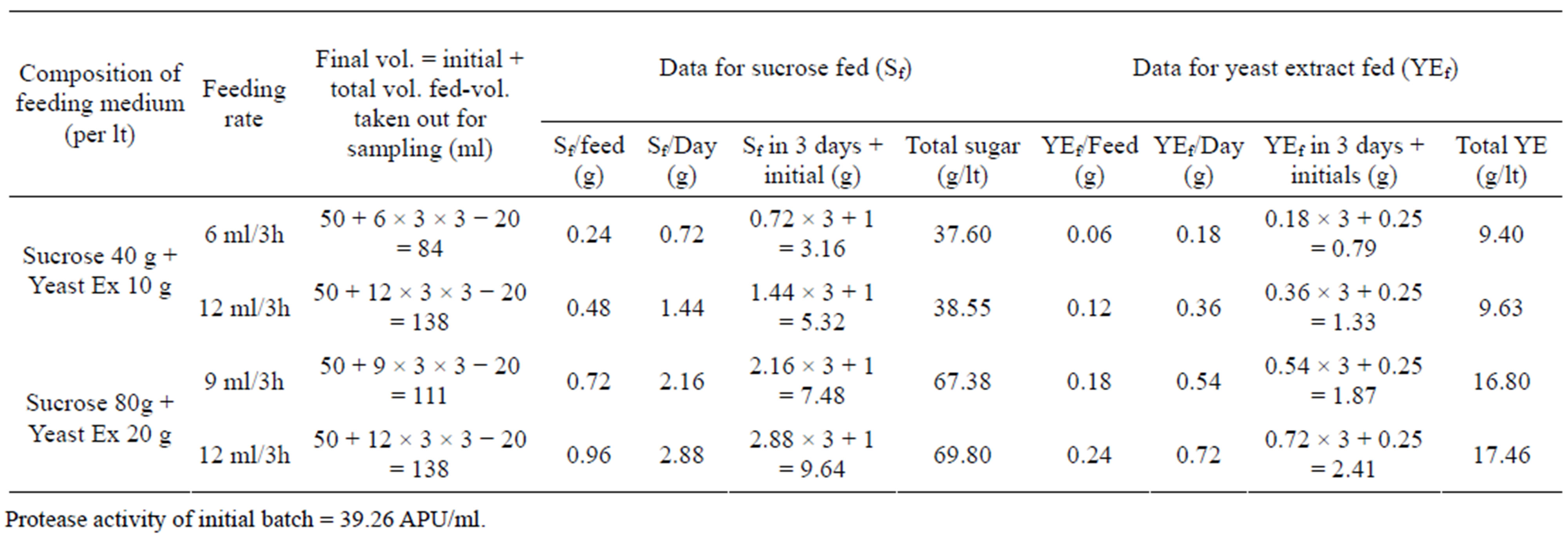
Table 2. Details of the fed-batch process conducted to examine the effects of different feed-rates of carbon and nitrogen sources of the medium with different compositions.
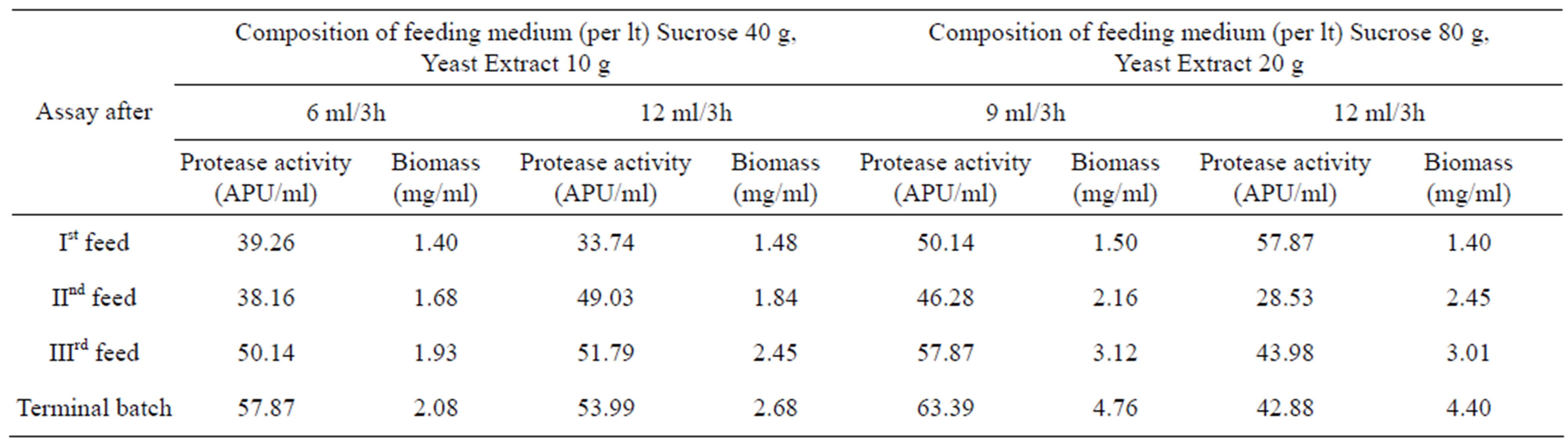
Table 3. Profiles of the concentrations of cell mass and protease activities for the fed-batch process with different feed-rates of the medium under two different feeding compositions.
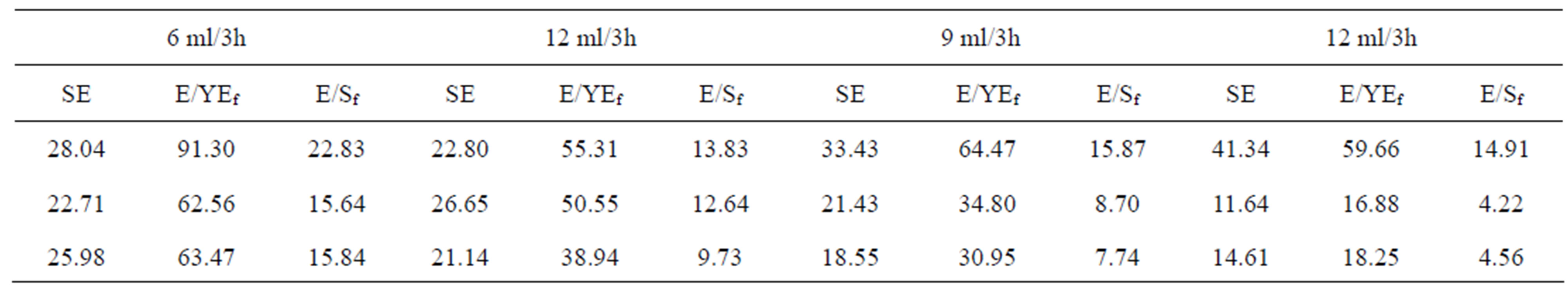
Table 4. Results of the different fed-rates of carbon and nitrogen sources. SE: Specific protease activity based on dry cell mass. E/YEf: Protease yield based on yeast extract fed; E/Sf: Protease yield based on sucrose fed.
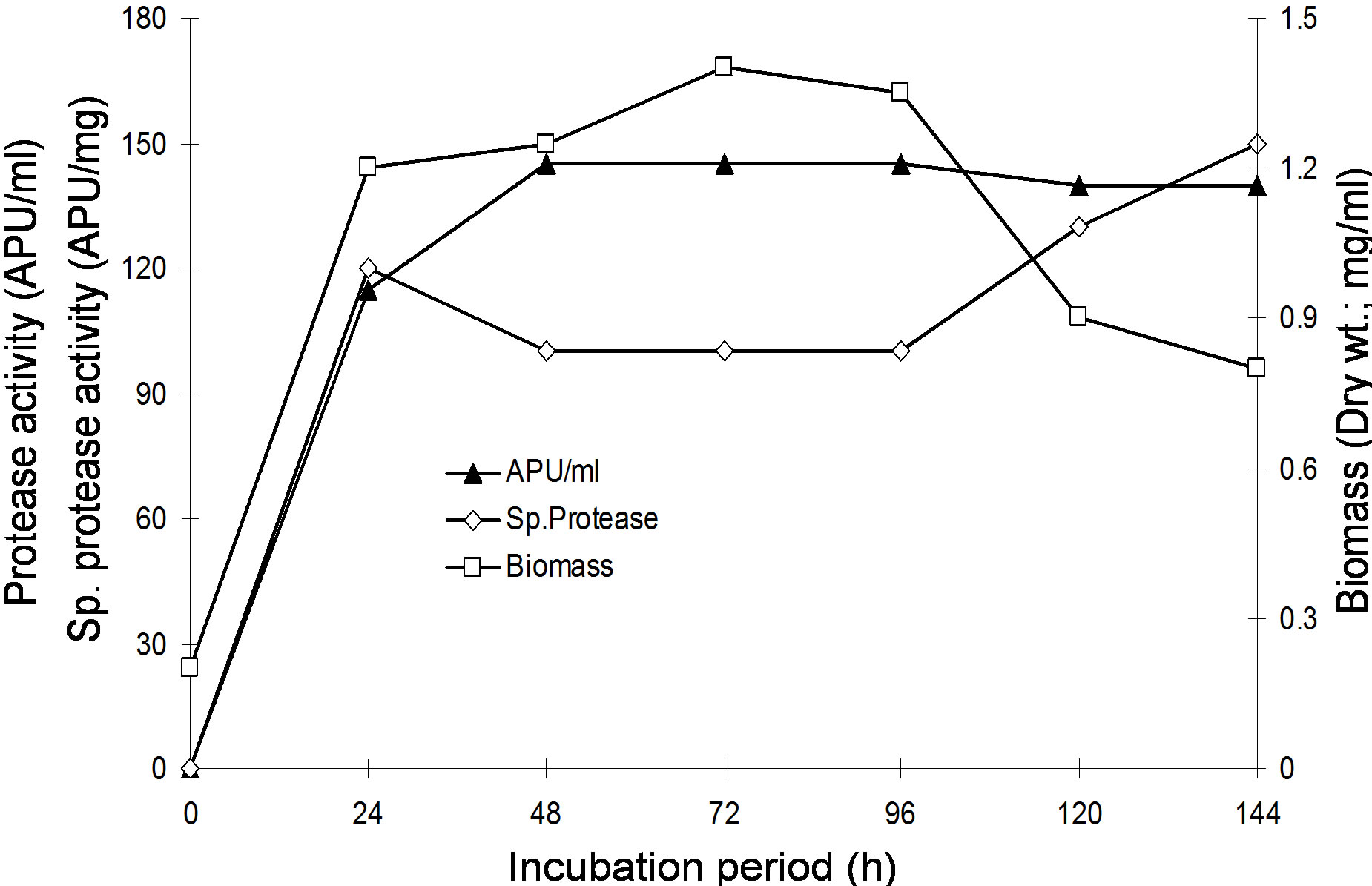
fed-batch operation at 50 ml and 150 ml with the fixed volumetric feed rate of 40 ml/day. A simultaneous batch experiment, under the similar working conditions with a working volume of 150 ml was carried out and the results are summarized in Table 5. The profiles of concentration of cell mass, protease activities and specific protease activities for batch and fed-batch experiments with constant feed rates are presented in Figure 2. The results show that the fed-batch experiment with medium feeding strategy of 20 ml/7h (FB-1) resulted in high protease production. Protease activity of 143.49 APU/ml was obtained only after 48 h of incubation and that much of activity was retained for up to 96 h of incubation period (Figure 2), showing the same percentage utilization of substrate into biomass and product. A further 36.66% increased protease activity was resulted in the next 24 h, while a rise by 15.69% of specific protease activity was attainable during the time-course of the fed-batch experiment. FB-1 resulted in 181% increased protease production rate than FB-2.
When the fed-batch experiments with different medium feeding strategies were compared with the simultaneous batch run (Table 3), it was clearly shown that fed-batch experiments successfully simulated early enzyme production (110.38 APU/ml after 24 h) that might be attributed to some oxygen transfer limitation in batch
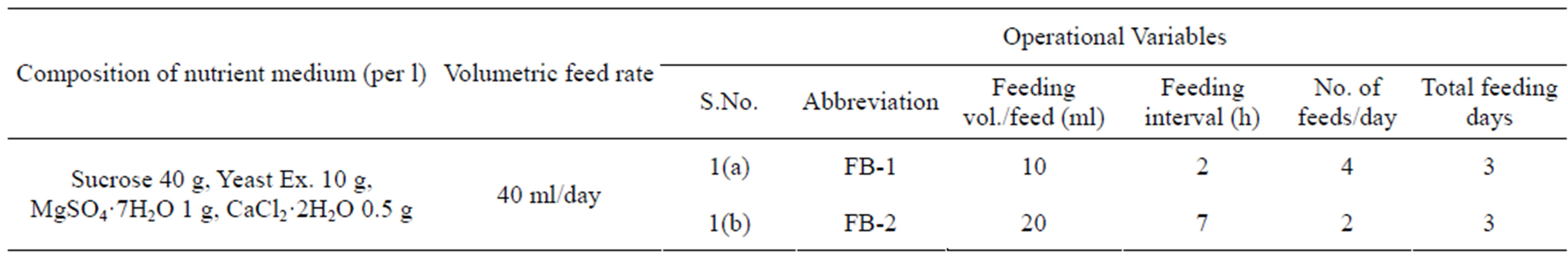
Table 5. Details of operational variables in the fed-batch process for the effect of feed intervals of the whole medium. pH, temperature and agitation speed were maintained at 10.0, 50˚C and 200 rpm, respectively.
 (a)
(a) (b)
(b) (c)
(c)
Figure 2. The growth, protease production and specific protease production during cultivation of the Bacillus sp. BHA. (a) Fed-batch culture with 20 ml of medium fed at the interval of 7 h; (b) Fed-batch culture with 10 ml of medium fed at the interval of 2 h; (c) Batch culture with the same medium composition.
process. Further, the fed-batch cultures showed higher specific enzyme production rates than the ordinary batch process. FB-1 and FB-2 resulted in the rise of protease productivity by 36.84% and 26.32%, respectively as compared to the peak protease activity attainable in batch process. At the terminal batch operation further 11.11%, 15.75% and 10.08% increased protease activities were attainable in FB-1, FB-2 and batch process, respectively.
Thus, we isolated the new Bacillus sp. with high potentiality for alkaline protease production from the degraded protein samples. Four significant enriched-yielders for protease production were screened and compared for the alkaline production. The best isolate (Bacillus sp. BHA) was further screened on different carbon and nitrogen sources and optimum pH (9.0) and temperature (60˚C) for alkaline protease production were determined. The obtained protease had maximum stability at high pH (9.0) and temperature (60˚C). The strain was out of repression at high sucrose (4%) and yeast extract (1%). Fed-batch experiments were largely conducted to investigate the repression of protease synthesis by excess yeast extract and effect of concentration of the carbon source (sucrose) on protease fermentation. Further, fed-batch experiment was optimized for the enhancement of alkaline protease production.
REFERENCES
- Raghvan, R. (2000) International spectrum in: News from abroad. Chemical Weekly, 45, 127-128.
- Cowan (1983) Protein: In industrial enzymology. In: Godfrey, T. and Reichell, K., Eds., Macmillon, London, 352- 374.
- Christic, R.B. (1980) Topics in enzyme and fermentation biotechnology. In: Wiseman, A., Ed., Ellis Horwood, Chishesler, 25-83.
- Mass, F. and Tilburg, R. (1983) The benefit of detergent enzymes under changing washing conditions. Journal of the American Chemical Society, 60, 1672-1675.
- Anisworth, S.J. (1994) Soap and detergent. Chemical & Engineering News, 72, 32-59.
- Gupta, R., Gupta, K. and Saxena, R.K. (1999) Bleach stable alkaline protease from Bacillus sp. Biotechnology Letters, 25, 135-138. doi:10.1023/A:1005478117918
- Loffer, A. (1986) Proteolytic enzymes: Sources and applications. Food Technology, 40, 63-70.
- Anwar, A. and Salemuddin, M. (1998) Alkaline protease: A review. Bioresource Technology, 64, 175-183. doi:10.1016/S0960-8524(97)00182-X
- Nakagawa, Y. (1970) Alkaline proteases from aspergillus. Methods in enzymology. In: Perlmann, G.E. and Lorand, L., Eds., Academic Press, Waltham, 582-583.
- Lowry, O.H., Rosebrough, N.J., Farr, A.L. and Randall, J.R. (1951) Protein measurement with the folin phenol reagent. Journal of Biological Chemistry, 193, 265-275.
- Breed, R.S. (1957) Family XIII bacilaceae fischer, In Bergey’s manual of determinative bacteriology. Baltimore, Williams, 613-634.
- Sinha, N. and Satyanarayana, T., (1991) Alkaline protease production by thermophilic Bacillus licheniformis. Indian Journal of Microbiology, 31, 425-430.
- Outtrup, H. (1995) New alkaline protease from Bacillus DSM 8473. WO-9507350.
- Sandhia, G.S. and Prema, P. (1998) Selection of optimal growth medium for the synthesis of alkaline protease from Bacillus SGP-26. Journal of Scientific & Industrial Research, 57, 629-633.
- Chu, I.M., Lee C. and Li, T.S. (1992) Production and degradation of alkaline protease in batch cultures of Bacillus subtilis ATCC 14416. Enzyme and Microbial Technology, 14, 755-761. doi:10.1016/0141-0229(92)90116-6
NOTES
*This work was supported by the part of the M.S. Thesis from the Department of Biochemical Engineering and Food Technology, Harcourt Butler Technological Institute.