American Journal of Plant Sciences
Vol.07 No.02(2016), Article ID:63306,8 pages
10.4236/ajps.2016.72025
ISSR Fingerprinting to Ascertain the Genetic Relationship of Curcuma sp. of Tripura
Kishan Saha1, Rabindra Kumar Sinha1, Surajit Basak2, Sangram Sinha1*
1Department of Botany, Tripura University, Suryamaninagar, India
2Department of Molecular Biology and Bioinformatics, Tripura University, Suryamaninagar, India

Copyright © 2016 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 21 December 2015; accepted 31 January 2016; published 3 February 2016
ABSTRACT
Molecular fingerprints of four different species of Curcuma, viz., C. amada, C. caesia, C. longa and C. zedoaria, found in Tripura were developed using Inter Simple Sequence Repeats. Twenty ISSR primers generated 116 loci amplified in the range of 200 - 5000 bp with an average of 5.8 alleles and 1.6 effective alleles per locus. The percentage of polymorphic band was found to be 86.29 with an average of 5.15 per primer. Based on UPGMA algorithm these four species are placed in two different clusters that validate the classification based on external and internal morphological characters. The polymorphic ISSR markers generated from this study will be useful for understanding the genetic relationship of different species of the genus Curcuma.
Keywords:
Curcuma, Genetic Diversity, Inter Simple Sequence Repeats, Polymerase Chain Reaction, Taxonomy, Zingiberaceae

1. Introduction
The genus Curcuma belonging to the family Zingiberaceae comprises ca. 80 species and shows the widespread distribution from tropical Asia to Australia and South pacific region [1] . The highest diversity of Curcuma has been found in India and Thailand and about 40 species are indigenous to India [2] . Different species of Curcuma have immense medicinal value and have been extensively used in indigenous system of medicine [3] - [6] . It is now well documented that the position of the spike, presence of coma bract and the color of bract in Curcuma are the major distinctive traits for delineation of species [7] . However, variation in the position of the spikes and bract color has also been noted in some species of Curcuma [8] . The state of Tripura situated in the sub Himalayan region of North East India is one of the hotspot of Indo-Burma biodiversity region of the world [9] [10] . Previously three species of Curcuma, viz., Curcuma amada, C. longa (=C. domestica) and C. zedoaria were reported from Tripura [11] . In our recent survey, we have identified another species C. caesia from West Tripura. To study the evolutionary history of a species knowledge of genetic variation is a prerequisite and it is essential to characterize the plants genetically in order to have a sustainable conservation programme [12] . DNA based molecular markers show differences in nucleotide sequence of DNA and are now used as powerful tools in the field of Plant breeding, Taxonomy, Physiology and Genetic engineering [13] . ISSR markers are technically simpler than other markers. These markers are mostly dominant except a few which are codominant in nature. In this technique, unlimited number of primers can be synthesized and the advantage lies in long primer length and stringent annealing temperature [14] . In higher plants, Inter Simple Sequence Repeat or ISSR markers are therefore frequently used because they are known to be abundant, very reproducible and highly polymorphic [14] [15] . Moreover, the ISSR based molecular fingerprinting technique is a good alternative to AFLP when tested on Curcuma species [16] [17] . Till now, there is no report on the genetic relationship of Curcuma species grown in diverse habitats of Tripura. An attempt has, therefore, been undertaken for molecular characterization of four species of wild and cultivated Curcuma grown in the state of Tripura using ISSR markers.
2. Materials and Methods
2.1. Plant Material and DNA Extraction
Rhizomes of four species of Curcuma, viz., C. amada Roxb., C. longa L., C. zedoaria (Christm.) Roscoe and C. caesia Roxb. found in wild state were collected from different geographical locations of Tripura (Table 1) and grown in the experimental garden of Department of Botany, Tripura University for experimental purposes. In addition to these, rhizomes of two populations of cultivated C. longa were also grown in the experimental garden for the present study. Total genomic DNA was extracted according to the manufacturer’s protocol (DNeasy® Plant Mini Kit-Qiagen, part no.69104). DNA concentration was determined using the Nanodrop 2000C spectrophotometer (Thermo Scientific-USA) and qualitative study was performed in 1.5% Agarose gel.
2.2. ISSR Analysis
For the genetic diversity study of four different Curcuma species, 20 ISSR markers (Sigma Aldrich, India) were chosen (Table 2). PCR amplification was performed using a mixture of 25 µl containing genomic DNA (30 ng/µl), dNTPs 10mM (Qiagen), 25 mM of MgCl2 (Sigma), 10× Taq buffer (Sigma), 10 µM primer and 2.5 Unit of Taq Polymerase (Sigma). PCR amplification was carried out in a Thermal Cycler (Applied Biosystems, Gene Amp* PCR System 9700). PCR was performed at an initial temperature of 94˚C for 4 minutes for complete denaturation. The second step consisted of 44 cycles having three ranges of temperature: 94˚C for 1 minutes, 50˚C for 1.30 minutes for primer annealing and 72˚C for primer extension, followed by 72˚C for 10 minutes. All amplified reactions were repeated at least two times for confirmation. The amplified products were visualized using 2% Agarose gel electrophoresis and scanned through a gel documentation system.
2.3. Data Analysis
The amplified fragments obtained from the ISSR profile were scored as binary data (1/0 for the presence or ab-

Table 1. Different species of Curcuma collected from different locations of Tripura.
Table 2. Total number of amplified fragments generated by PCR using ISSR primers.
sence) of each fragment. Only clear and reproducible bands were taken into account; the intensity of the bands was not considered. The numbers of polymorphic and monomorphic bands were determined for each primer in all species studied. Polymorphic Information Content (PIC) was computed using the formula
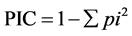
where pi is the frequency of ith allele at a given locus [18] and Marker Index (MI) was calculated [19] . The number of observed alleles, mean number of effective alleles [20] , Nei’s [21] gene diversity index (H) and Shannon index [22] were calculated using the POPGENE software [23] . The level of similarity between the species was established using DICE’s coefficient [24] . Similarity coefficients were used to construct the dendrogram using the SAHN subroutine through the NTSYS pc (Numerical Taxonomy System, 2.21q version) [25] . Further, Principal Coordinate Analysis (PCA) was performed with modules of STAND, CORR and EIGEN of NTSYS pc using the Euclidean distances with the help of NTSYS pc-2.21q software.
3. Results
Twenty ISSR primers that were used to characterize the genetic diversity among the species yielded 116 fragments with an average of 5.8 alleles and 1.6 effective alleles per locus (Table 3). In the present observation, it was found that out of the total amplified products, 13 bands were monomorphic and 103 were polymorphic and these were amplified in the range of 200 - 5000 bp. Maximum number of bands were recorded in HB 12, 825, UBC 873 and 807. However, the average number of polymorphic bands obtained per primer was 5.15. The per-
Table 3. Degree of polymorphism and polymorphic information content for ISSR primers in four species of Curcuma.
NSB: Number of score band, PB: polymorphic bands, MB: monomorphic bands, PPB: Percentage polymorphic band, PIC: average polymorphic information content for polymorphic bands, MI: marker index = POL (%) × PIC.
centage of ISSR polymorphic bands were found to range from 66.67 - 100, whereas the highest and lowest PIC values for ISSR primers were recorded in HB (0.49) and 809 (0.19) with an average of 0.34. The MI value on the other hand, was maximum in HB (48.56) and minimum in 809 (12.54). Screening of genetic diversity at interspecific and intraspecific levels have been done using POPGENE software and the average values of the observed number of alleles (na), effective number of alleles (ne) Nei’s gene diversity index (h) and the mean Shannon index (I) were found to be 1.86,1.60, 0.34 and 0.50 respectively (Table 4).
In the present study, genetic relationship among the four species of Curcuma shows two clusters expressed as UPGMA dendrogram using SAHN Neighbor Joining tree (Figure 1). The coefficients on the X axis represent the similarity indices of the different species chosen for the study. Based on UPGMA clustering, the genotype of C. zedoaria and C. caesia belongs to one cluster and that of C. amada and C. longa in a separate cluster. Dice’s coefficient showed that C. zedoaria and C. caesia were related to each other with a similarity value of 0.6379 whereas the similarity value between C. longa and C. amada was found to be 0.5593 (Table 5). PCA was analyzed on the basis of ISSR data which shows that the first 3 coordinate components accounted for 38.54%, 23.67% and 18.72% variation (Figure 2).

Figure 1. Dendrogram representing the genetic variability of Curcuma sp. using Dice similarity coefficient.
Table 4. Results of polymorphic primers screening in four species of Curcuma.
na = observed number of alleles; ne = effective number of alleles; h = Nei’s gene diversity; I = Shannon’s information index.

Figure 2. Principal coordinate analysis (PCA) map for the species of Curcuma.
Table 5. Dice similarity coefficient among the species of Curcuma.
4. Discussion
In Tripura so far we have recorded four species of Curcuma and they differ in morphological and anatomical characters to a certain extent [26] . A priori, key to the species identification in Curcuma was based on external and internal morphological characters, but relying on morphological characters alone in species delineation has its limitations. While majority of the morphological characters of C. caesia and C. zedoaria are more or less similar, the flower color and the internal anatomy of rhizome differs and the cortical zone of the rhizome of C. caesia shows bluish green color. C. longa is used mostly as an important spice and so, is extensively cultivated throughout Tripura. However, C. longa was also found in the wild state but remain restricted to the higher altitude of Jampui hills of Tripura. Morphologically C. longa and C. amada are almost similar but their rhizomes differ in color and odor [11] . The rhizome of C. longa is deep orange yellow in color and that of C. amada is pale yellow having the aroma of mango. ISSR cluster analyses reveal the presence of two distinct clusters in the wild and the cultivated Curcuma species studied; cluster I represents C. caesia and C. zedoaria and cluster II includes C. longa and C. amada and, the results thus obtained are not in full agreement with previous findings [6] [16] [17] . The genetic diversity of different species of Curcuma from the North Eastern region of India was also assessed [6] using ISSR fingerprinting but the formation of an independent cluster of C. caesia alone as was reported could not be ascertained in our present study even after repeated experimental trials. The presence of C. caesia and C. zedoaria in the same cluster and their similarity indices indicate that they might have arisen from a common ancestor inspite of their diverse ecological habitats. PCA depicts the variability among the species of Curcuma and three principal components with Eigen value greater than 1 extracted a cumulative of 80.83% variation. In all the taxa studied, there are sequence specific profiles (Figure 3) and the dendrogram shows that the genome of each species is not exactly the same. The somatic chromosome number of C. amada (=42) and C. longa (=63) differs (unpublished) due to the difference in their ploidy level, but differences in somatic chromosome number does not affect the similarity indices between the two species as is evident from the experimental data. Out of the two populations of cultivated C. longa3 (population-II) is genetically closer to that of C. longa1 found in the wild state. This resemblance suggests that probably C. longa1 found in the wild state escaped
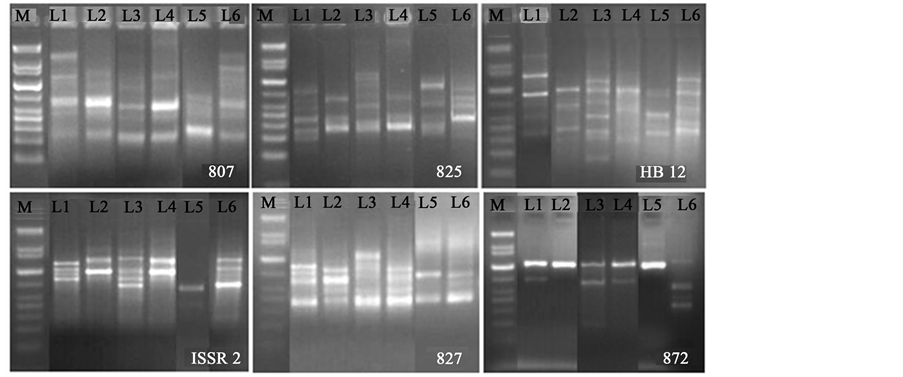
Figure 3. ISSR fingerprints of four species of Curcuma with primer 807, 825, HB12, ISSR2, 827 and 872 respectively M-1kb plus DNA ladder (Qiagen), L1-L6 represents, Curcuma amada, Curcuma longa1, Curcuma longa2, Curcuma longa3 and Curcuma zedoaria and Curcuma caesia.
earlier from the cultivated form. The genetical distance between C. longa2 and C. longa3 may be attributed to varietal distinction. Taken together, our findings support the taxonomic key to the identification of taxon at species level.
5. Conclusion
The molecular profiling of four species of Curcuma validates the conventional taxonomic interpretation. Interspecific and intraspecific variation observed with respect to degree of polymorphism, number of alleles observed, number of effective alleles, Nei’s gene diversity and Shannon’s information index are all indicators ascertaining genetic diversity of Curcuma species in Tripura. Thus, ISSR fingerprint can be used not only as an effective parameter to assess the genetic relationship between the species of Curcuma but also provides additional support for establishing the taxonomic position of a species.
Acknowledgements
The first author is grateful to University Grants Commission, New Delhi for providing BSR fellowship. The authors thankfully acknowledge the support provided by Dr. B. Dholakia, NCL, Pune and Dr. A.K. Banerjee, IISER, Pune for carrying out this work.
Cite this paper
KishanSaha,Rabindra KumarSinha,SurajitBasak,SangramSinha, (2016) ISSR Fingerprinting to Ascertain the Genetic Relationship of Curcuma sp. of Tripura. American Journal of Plant Sciences,07,259-266. doi: 10.4236/ajps.2016.72025
References
- 1. Skornickova, J.L., Otakar, S., Jarolimova, V., Sabu, M., Fer, T., Travnicek, P. and Suda, J. (2007) Chromosome Numbers and Genome Size Variation in Indian Species of Curcuma (Zingiberaceae). Annals of Botany, 100, 505-526.
http://dx.doi.org/10.1093/aob/mcm144 - 2. Velayudhan, K.C., Muralidharan, V.K., Amalraj, V.A., Gautam, P.L., Mandal, S. and Kumar, D. (1999) Curcuma Genetic Resources. Scientific Monograph No. 4, National Bureau of Plant Genetic Resources, New Delhi.
- 3. Mridula, K.R. and Jayachandran, B.K. (2001) Quality of Mango Ginger (Curcuma amada Roxb.) as Influenced by Mineral Nutrition. Journal of Tropical Agriculture, 39, 182-183.
- 4. Matsuda, H., Tewtrakul, S., Morikawa, T., Nakamura, A. and Yoshikawa, M. (2004) Anti-Allergic Principles from Thai Zedoary: Structural Requirements of Curcuminoids for Inhibition of Degranulation and Effect on the Release of TNF-Alpha and IL-4 in RBL-2H3 Cells. Bioorganic & Medicinal Chemistry, 12, 5891-5898.
http://dx.doi.org/10.1016/j.bmc.2004.08.027 - 5. Sarma, G.C. and Kalita, S. (2005) Zingiberaceae of Assam: Their Taxonomy and Utilization. Ph.D. Thesis, Botany Department, Gauhati University, Assam.
- 6. Das, A., Kesari, V., Satyanarayana, V.M., Parida, A. and Rangan, L. (2011) Genetic Relationship of Curcuma Species from North East India Using PCR Based Markers. Molecular Biotecnology, 49, 65-76.
http://dx.doi.org/10.1007/s12033-011-9379-5 - 7. Larsen, K. and Smith, R.M. (1978). A New Species of Curcuma from Thailand. Notes Royal Botanical Garden. Edinburgh, 30, 269-272.
- 8. Sabu, M. (1991) A Taxonomic and Phylogenetic Study of South Indian Zingiberaceae. Ph.D. Thesis, Department of Botany, University of Calicut.
- 9. Myers, N., Mittermier, R.A., Mittermier, C.G., Fonseca da, G.A.B. and Kent, J. (2000) Biodiversity Hotspots for Conservation Priorities. Nature, 40, 853-858.
http://dx.doi.org/10.1038/35002501 - 10. Mao, A.A., Hynniewta, T.M. and Sanjappa, M. (2009) Plant Wealth of Northeast India with Reference to Ethnobotany. Indian Journal of Traditional Knowledge, 8, 96-103.
- 11. Deb, D.B. (1981) The Flora of Tripura. Vol. I & Vol. II, Today and Tomorrow’s Publ., New Delhi.
- 12. Islam, M.A., Meister, A., Schubert, V., Kloppstech, K. and Esch, E. (2006) Genetic Diversity and Cytogenetic Analyses in Curcuma zedoaria (Christm.) Roscoe from Bangladesh. Genetic Resources and Crop Evolution, 54, 149-156.
http://dx.doi.org/10.1007/s10722-005-2639-0 - 13. Kesawat, M.S. and Daskumar, B. (2009) Molecular Markers: It’s Application in Crop Improvement. Journal of Crop Science and Biotechnology, 12, 169-181.
http://dx.doi.org/10.1007/s12892-009-0124-6 - 14. Zietkiewicz, E., Rafalski, A. and Labuda, D. (1994) Genome Fingerprinting by Simple Sequence Repeat (SSR)-Anchored Polymerase Chain Reaction Amplification. Genomics, 20, 176-183.
http://dx.doi.org/10.1006/geno.1994.1151 - 15. Bornet, B. and Branchard, M. (2004) Use of ISSR Fingerprints to Detect Microsatellites and Genetic Diversity in Several Related Brassica taxa and Arabidopsis thaliana. Hereditas, 140, 245-248.
http://dx.doi.org/10.1111/j.1601-5223.2004.01737.x - 16. Syamkumar, S. and Sasikumar, B. (2007) Molecular Marker Based Genetic Diversity Analysis of Curcuma Species from India. Scientia Horticulturae, 112, 235-241.
http://dx.doi.org/10.1016/j.scienta.2006.12.021 - 17. Singh, S., Panda, M.K. and Nayak, S. (2012) Evalution of Genetic Diversity in Turmeric (Curcuma longa L.) Using RAPD and ISSR Markers. Industrial Crops and Products, 37, 284-291.
http://dx.doi.org/10.1016/j.indcrop.2011.12.022 - 18. Roldan-Ruiz, I., Dendauw, J., Van Bockstaele, E., Depicker, A. and De Loose, M. (2000) AFLP Markers Reveal High Polymorphic Rates in Ryegrasses (Lolium spp.). Molecular Breeding, 6, 125-134.
http://dx.doi.org/10.1023/A:1009680614564 - 19. Powell, W., Morgante, M., Andre, C., Hanafey, M., Vogel, J., Tingey, S. and Rafalski, A. (1996) The Comparison of RFLP, RAPD, AFLP and SSR (Microsatellite) Markers for Germplasm Analysis. Molecular Breeding, 2, 225-238.
http://dx.doi.org/10.1007/BF00564200 - 20. Kimura, M. and Crow, J.F. (1964) The Number of Alleles That Can Be Maintained in a Finite Population. Genetics, 49, 725-738.
- 21. Nei, M. (1973) Analysis of Gene Diversity in Subdivided Populations. Proceedings of the National Academy of Sciences of the United States of America, 70, 3321-3323.
http://dx.doi.org/10.1073/pnas.70.12.3321 - 22. Lewontin, R.C. (1972) Testing the Theory of Natural Selection. Nature, 236, 181-182.
http://dx.doi.org/10.1038/236181a0 - 23. Yeh, F.C., Yang, R.C., Boyle, T.B.J., Ye, Z.H. and Mao, J.X. (1997) POPGENE, the User Friendly Shareware for Population Genetic Analysis. Molecular Biology and Biotechnology Centre, University of Alberta, Alberta.
- 24. Dice, L.R. (1945) Measures of the Amount of Ecologic Association between Species. Ecology, 26, 297-302.
http://dx.doi.org/10.2307/1932409 - 25. Rohlf, F.J. (2002) NTSYSPC. Numerical Taxonomy System. Exter Software, New York.
- 26. Prain, D. (1963) Bengal Plants. Vol. I & Vol. II Reprinted Edition, Botanical Survey of India, Kolkata.
NOTES

*Corresponding author.