American Journal of Plant Sciences
Vol.4 No.12C(2013), Article ID:40794,6 pages DOI:10.4236/ajps.2013.412A3001
Quantitative Detection of Inositol Hexakisphosphate (InsP6) in Crop Plants Using Polyacrylamide Gel Electrophoresis (PAGE)
![]()
Department of Applied Science, University of Arkansas at Little Rock, Little Rock, USA.
Email: *mvkhodakovsk@ualr.edu
Copyright © 2013 Mohammad Alimohammadi et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received July 30th, 2013; revised August 31st, 2013; accepted September 16th, 2013
Keywords: Inositol Hexakisphosphate; Polyacrylamide Gel Electrophoresis; High-Performance Liquid Chromatography; Nutrient Deficiency
ABSTRACT
Inositol phosphates are essential for cell development and signaling in all living organisms. Inositol hexakisphosphate (InsP6) is the most abundant phosphoinositol in both plants and animals. While the concentration of inorganic phosphorous (Pi) is often limited in soil, some plants overcome this limitation by creating a phosphate reservoir that serves as a source of Pi during phosphate deficiency. Although this strategy benefits plant development and signaling under adverse environmental conditions, excessive accumulation of Pi in crop plants has raised serious concerns about its toxicity and ill effects on human health. Consumption of crop plants with high InsP6 content or food products made from these crops is found to reduce nutrient intake significantly by way of chelating essential metal cations in human and livestock fed by such plants. Therefore, it is necessary to determine InsP6 contents in crop plants. Several methods have been developed for the screening and detection of InsP6 in plants. These detection methods however, are complex, labor-intensive, and often provide inaccurate results. We have developed a fast, reliable, and cost-effective method for the detection and quantification of InsP6 in plants using polyacrylamide gel electrophoresis (PAGE) with potential applications in industry, quality control labs, and research projects.
1. Introduction
Phytic acid or phytate (inositol hexakisphosphate, InsP6), is one of the major components of seeds, leaves, and fruits in many plant species [1-3]. It is considered as the reservoir for phosphorus storage in plants and is used in case of phosphate deficiency in soil. Phytate is primarily presented as phytin in combination with K+, Ca2+, Mg2+ or Zn2+ and is deposited in plant cells together with proteins [4]. The formation of phytate occurs both in seeds and vegetative tissue [5]. Biosynthesis of fundamental molecules, such as starch, lipids and proteins, during developmental processes in plant cells is followed by the accumulation of phytate in plant tissues [6]. The mechanism by which phytate is synthesized in plants is not well understood. Several attempts have been made, howeverto study the mechanism of the InsP6 biosynthetic pathway in plants [7-9]. Although more investigations are needed to explain the possible functions of phytate in plants, excessive accumulation of phytate is found to reduce availability of important nutritional compounds when consumed by human or used as animal feed [10]. In human and animals that only have one stomach in their digestive tract, phosphorous is not readily bioavailable. In these organisms, consumption of food products such as legumes, grains, and cereals with high-phytate concentration not only affects bioavailability of phosphorous, but also phytic acid binding to iron, zinc and calcium molecules and makes these minerals unavailable for the body (Figure 1). Phytic acid also inhibits essential digestive enzymes such as pepsin and amylase [11]. These enzymes are required to break down proteins and starch respectively. A diet rich in phytic acid will induce
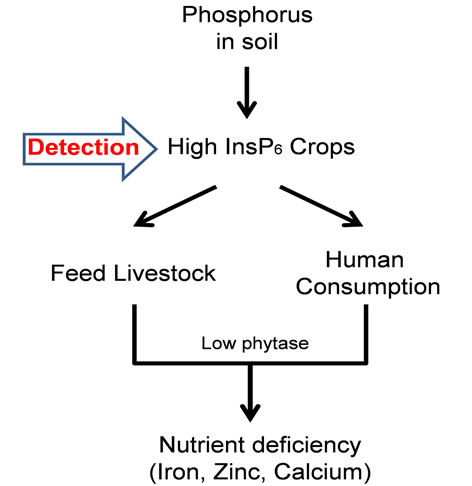
Figure 1. Schematic representation of the effect of high InsP6 crops on nutrient uptake in human and animals.
toxicity and negatively affect nutrition uptake and as a result cause several health problems including nutrient deficiencies, digestive problems and tooth decay.
All of these factors suggest that a reliable and efficient method for the quantitative detection of phytate in crop plants is of utmost importance. This will facilitate identification of high-phytate crop plants and limit or prevent their application in the food industry. The methods that are currently used for detection of phytate are complex, labor-intensive and involve enzymatic assays, high-performance liquid chromatography (HPLC), and radioactive labeling. Enzymatic assays involve extraction and phytase treatment of InsP6 followed by measurement of inorganic phosphorous (Pi) liberated during the enzymatic reaction. These assays are not reliable and do not give an accurate insight into the total phytate content in the tested samples. The HPLC method is more accurate but is complicated and difficult to perform because it is labor-intensive and often requires radioactive labeling. Metabolic radio-labeling of plant materials is expensive, cumbersome, and hazardous. Therefore, a simple, easy to perform method that is fast and reliable giving more accurate quantitation is needed to establish phytate contents in crop plants.
In this study, we have used polyacrylamide gel electrophoresis (PAGE) to separate and quantitatively visualize the InsP6 contents in plant samples. This method can be used as a tool for rapid screening of high-phytate crop plants. Our approach in developing this method is especially valuable because it can precisely detect and quantify InsP6 molecules in plant samples directly without dealing with complications of hydrolysis with phytase and then determine the Pi or the labor-intensive HPLC method involving the risks of working with radio-hazard materials. The simplicity and accuracy of this method suggest its potential as an effective alternative method for the detection and quantification of phytic acid in various plant tissues. To our knowledge, this is the first attempt to implement and quantify the phytate content in plants using PAGE.
2. Materials and Methods
2.1. Plant Growth Condition
Tomato (cv. Micro-Tom), rice (cv. Millie), and tobacco (cv. Petit Havana) plants were germinated in pots containing 25% sand and 75% Sun Gro Redi-earth “Plug and Seedling” Mix (Sun Gro Horticulture, Bellevue, WA) in a growth chamber with light intensity of 600 µmol∙m−2∙s−1 for 16 h light (28˚C) and 8 h dark (22˚C). Tissue samples were collected from the plants 1 month after germination and used for the detection of InsP6 using PAGE.
2.2. Extraction of Phytate from Plant Samples
Tissue samples were prepared using a modified version of the phosphoinositol extraction method described by Stevenson-Paulik et al. [6] Leaf tissue (200 mg ± 5) was weighed, transferred to 2 mL screw-cap tubes containing 1.0 mm Zirconia/Silica beads (BioSpec Inc. Bartsville, OK), and kept on ice. 500 µL of 0.4 M HCl were added to each tube. The tissue was homogenized using a MiniBead Beater (BioSpec Inc. Bartsville, OK) for 2 min. The homogenized tissue was then boiled for 5 min and disrupted again for 2 min. The leaf extracts were then centrifuged for 10 min at 13,000 g. The supernatant was transferred to new 1.5 mL microfuge tubes. Tubes containing supernatant were snap-frozen in liquid nitrogen and then thawed at room temperature. The samples were then centrifuged again at 13,000 g for 5 min. Following centrifugation, the supernatant was transferred to a fresh 1.5 mL microfuge tube and used for phytate detection by PAGE.
2.3. Polyacrylamide Gel Electrophoresis, Staining and Visualization
Phytate was detected on a 8 × 8.5 × 0.1 cm polyacrylamide gel prepared by modification of the method originally described by Losito et al. [12]. The polyacrylamide gel consisted of 33.3% resolving gel in TBE (3.7 mL 40% Acrylamide/Bis-acrylamide (Acr/Bis) (19:1); 0.5 mL 10 × TBE (pH 8.0); 0.3 mL ddH2O; 32 µL 10% APS; 3.5 µL TEMED) topped with a stacking gel (1 mL of 16% Acr/Bis (19:1); 120 µL 5 × TBE (pH 6.0); 69.5 µL ddH2O; 8.5 µL 10% APS; 1 µL TEMED). The gel was pre-run for 20 min. at 100 V/10 mA in 1× TBE buffer. Twenty microliters of the extract were mixed with 5 µL of 6× orange G loading dye (10 mM Tris-HCl pH 7.0; 1 mM EDTA; 30% glycerol; 0.1% Orange G) and loaded onto the gel. The gels were run at 100 V/10mA until the dye front reached 2 cm from the bottom of the gel. Gels were stained with a filtered toluidine blue (Sigma, MO, USA) staining solution (20% methanol; 2% glycerol; 0.05% Toluidine Blue) for 30 minutes. It was then de-stained for 1 hour with several changes of the same solution without the dye. Gel images were recorded during exposure to visible light using a gel imaging system (Bio-Rad, Hercules, CA).
2.4. Data Analysis
Phytate concentration was quantified by using the gel images in a GelQuant.NET software (biochemlabsolutions.com). The phytate concentration was expressed in nmol InsP6 per 20 µL of tissue extract based on the standard curve. Volume Density − 115.36 = nmol InsP6 × 59271 obtained over a dynamic range of 0 - 5 nmol InsP6 (R2 = 0.994). The values were then converted to nmol∙g−1 fresh weight of tissue.
3. Results and Discussion
Phosphorous is an essential element for plants. It plays a critical role in the formation of multiple cell components [13]. Plants use various mechanisms to accumulate and re-utilize inorganic phosphorous (Pi) to protect themselves against Pi deficiency in the soil [14]. Accumulation of Pi in plants in the form of organic phosphate takes the form of phytate when it combines with inositol at all six carbon positions (inositol hexkisphosphate, InsP6). Although phosphate is required for functionality of cell components and in signaling pathways, high concentrations of phytate in crop plants have always been controversial. Recent studies suggest that InsP6 can regulate carcinogenesis by inducing programmed cell death and could be used therefore for the treatment of various types of cancers [15-18]. On the other hand, it has been found that phytate prevents absorption of critical minerals and metal ions and therefore accumulation of phytate can cause cytotoxicity. Thus, the consumption of crop plants with high phytate content in the daily diet could lead to nutrient deficiency related diseases [19-21]. This is especially problematic among children at early stages of development. Phosphorous is essential for bone development in children. However, around 80 percent of phosphrous found in grains is found in the form of phytate which is unusable for the body [22]. Consumption of phytate rich products will cause calcium to bind with phytic acid and produce insoluble compounds. This will cause problem in absorption of calcium which is required for bone development. Phytate can also block absorption of iron and zinc which can cause serious nutritional problems. To avoid these consequences, consumption of phytate rich products should be reduced in daily diet. This can be achieved by selecting crop plants with low level of phytic acid in food industry. Considering these facts, a reliable and accurate method for the detection of high phytate crops would be helpful in preventing potentially toxic products from hitting the market.
Losito et al. [12] described a method of separating standard InsP6 and inositol pyrophosphates by PAGEbased gel electrophoresis. However, the efforts by the authors to apply this technique to detect inositol hexakisphosphate or its pyrophosphate derivatives in mammalian or yeast cells failed perhaps due to the poor sensitivity of the detection method as mammalian cells contain much lower amounts of these compounds than plants. We attempted to develop a new application of the PAGEbased separation of inositol phosphates and established a method for detecting phytate in plants. Our new application essentially uses PAGE to separate and visualize phytate in crop plants and shows promising results in the detection and quantification of phytate in various plant species.
We optimized this method for the extraction of phytate in tomato, rice, and tobacco samples. All of these plant species have significance as model crops in both agriculture and research. Our results demonstrated that phytate can be accurately visualized and quantified by using this new method.
As a first step, we validated the technique by application of different concentrations of standard InsP6 (0 - 5 nmol) and successfully visualized on the gel (Figure 2(a)). Commercially available phytate (Sigma, MO) was used as standard InsP6. Volume density was determined based on the visualized band intensity corresponding to each concentration of phytate loaded on the gel. This value was incorporated to create a standard curve that was used as a reference for the quantification of phytate in plant samples (Figure 2(b)).
In the next step, phytate was extracted from leaves of tomato, rice, and tobacco plants and was separated using PAGE as described in materials and methods. Gel images were captured using a gel imaging system after staining the gels with toluidine blue. The images clearly revealed the phytate content of the tested samples and demonstrated differences in phytate levels in the three different crop plants that were studied (Figure 3(a)).
The identity of the visualized band as phytate was confirmed by its sensitivity to phytase. To achieve this goal, tissue extracts and standard InsP6 were treated with phytase and incubated at 37˚C for 1 hour prior to loading on the gel and were compared with the samples that were not treated with the enzyme (Figure 3(a)). Phytase is the primary enzyme that can facilitate degradation of phytic acid to lower forms of inositol phosphates and inositol [23]. Therefore, incubation with phytase would breakdown the phytate to inositol derivatives which would
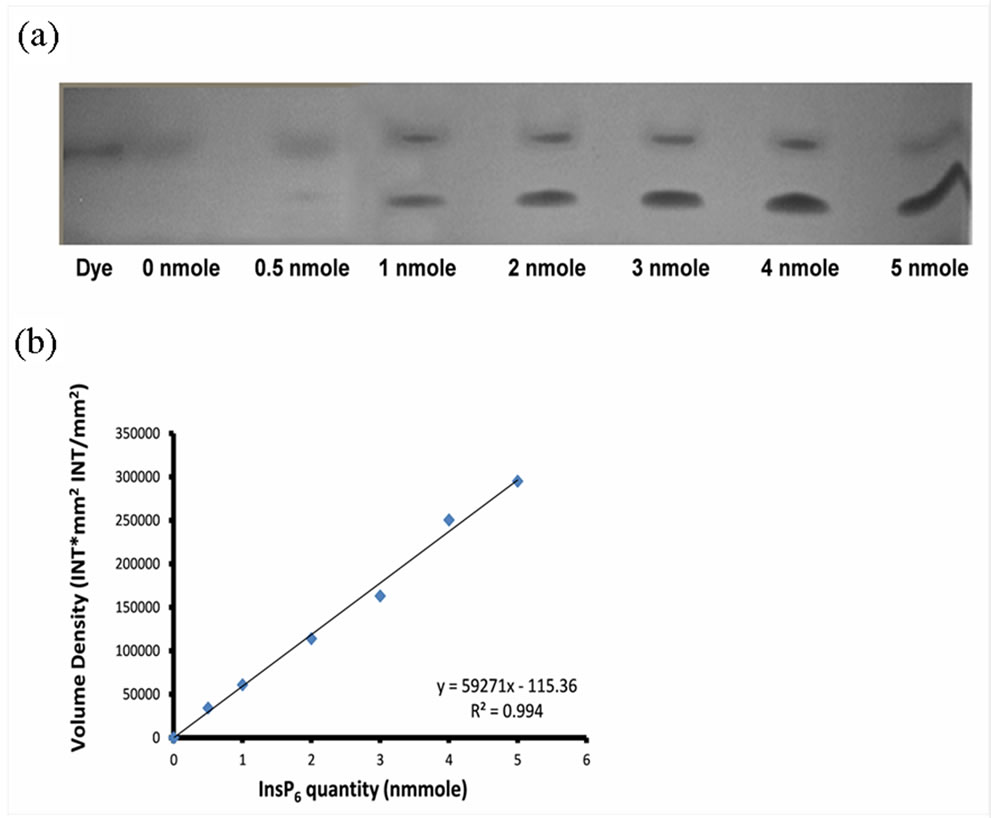
Figure 2. (a) Visualization of standard InsP6 using PAGE. Different concentrations of InsP6 (0 - 5 nmoles) were run on polyacrylamide gel. Gels were stained by toluidine blue and exposed to visible light using gel imaging system; (b) Standard curve based on the volume density obtained from gel images of InsP6 standards.
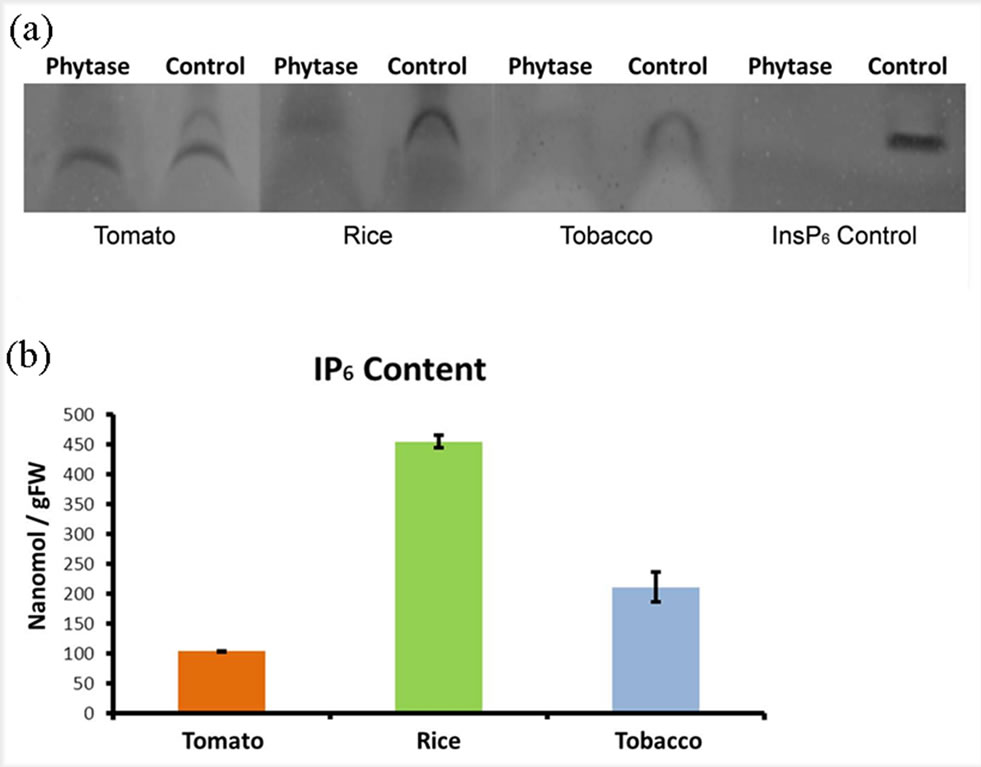
Figure 3. Visualization of InsP6 in tomato, rice and tobacco leaf samples using PAGE. (a) Samples were run directly onto the gel (control samples) or were pre-treated with phytase (phytase). InsP6 was used as standard (InsP6). For phytase treatment, tissue extracts were treated with phytase and incubated at 37˚C for 1 hour prior to loading onto gel and were compared with samples that were not treated with phytase. Data is a representative of three experiments performed independently; (b) Total amount of InsP6 in tomato, rice and tobacco leaf samples. Results are based on the volume density of bands obtained from gel. Three biological replicates were used for each experiment. Leaf samples were collected after 4 weeks of cultivation in a growth chamber with light intensity of 600 µmol∙m−2∙s−1 with 16 h light (28˚C) and 8 h dark (22˚C).
result in disappearance of the phytate band on the gel. Results revealed that observed bands on the gel were indeed related to InsP6 as treatment with the enzyme led to degradation of InsP6 and disappearance of the band (Figure 3(a)). The bands, however, were not straight and were bent at the bottom part of the gel. This can be explained by the fact that the total plant cell extract was loaded onto the gel. These extracts perhaps contained high salts and proteins which could have affected the running conditions of the gel but did not affect phytase digestion and overall quantitation.
Finally, gel images were used to determine the quantity of phytate in various plant samples. Data analysis using the standard curve showed that phytate contents vary among plant species and thus this method can be used to quantitate phytate contents in various crop plants (Figure 3(b)).
In conclusion, the phytate detection method described in this study can effectively facilitate the screening of crop plants for their InsP6 content. The detection of phytate by PAGE has several advantages over other methods which would make it a preferred choice for research studies and also for the mass detection and quantification of phytate in crop plants in quality test laboratories. This method is rapid, cost-effective, and reliable. It is also safer to use and more environmentally-friendly for it does not involve radioactive materials.
Further preparatory modifications in the phytate extraction method to minimize salt and protein contents are likely to minimize distortions in the bands on the gel. The PAGE-based method can also be an ideal substitute for the detection of InsP6 and other inositol polyphosphates in organisms including animal and human cells where InsP6 is present at much lower concentration. This would require an improved staining procedure which is more sensitive than toluidine blue. Other cationic dyes that have higher affinity for InsP6 and that fluoresce might enhance the sensitivity of detection.
REFERENCES
- D. Oberleas, “Phytates,” In: Toxicants Occurring Naturally in Foods, 2nd Edition, National Academy of Sciences, Washington DC, 1973, pp. 363-371.
- F. A. Loewus, “Structure and Occurrence of Inositol in Plants,” In: D. J. Morri, W. F. Boss and F. A. Loewus, Eds., Inositol Metabolism in Plants, Wiley-Liss Inc., New York, 1990, pp. 1-11.
- V. Raboy, “Biochemistry and Genetics of Phytic Acid Synthesis,” In: D. J. Morré, W. F. Boss and F. A. Loewus, Eds., Inositol Metabolism in Plants, Wiley-Liss Inc., New York, 1990, pp. 55-76.
- J. N. A. Lott, “Accumulation of Seed Reserves of Phosphorus and Other Minerals,” In: D. R. Murray, Ed., Seed Physiology, Academic Press, 1984, pp. 139-166.
- J. N. A. Lott, J. S. Greenwood and G. D. Batten, “Mechanisms and Regulation of Mineral Nutrient Storage during Seed Development,” In: J. Kigel and G. Galili, Eds., Seed Development and Germination, Marcel Dekker, New York, 1995, pp. 215-235.
- J. Stevenson-Paulik, R. J. Bastidas, S. T. Chiou, R. A. Frye and J. D. York, “Generation of Phytate-Free Seeds in Arabidopsis through Disruption of Inositol Polyphosphate Kinases,” Proceedings of the National Academy of Sciences, Vol. 102, No. 35, 2005, pp. 12612-12617. http://dx.doi.org/10.1073/pnas.0504172102
- J. S. Greenwood and J. D. Bewley, “Subcellular Distribution of Phytin in the Endosperm of Developing Castor Bean: A Possibility for Its Synthesis in the Cytoplasm Prior to Deposition within Protein Bodies,” Planta, Vol. 160, No. 2, 1984, pp. 113-120. http://dx.doi.org/10.1007/BF00392859
- L. R. Stephens and R. F. Irvine, “Stepwise Phosphorylation of Myo-Inositol Leading to Myo-Inositol Hexakisphosphate in Dictyostelium,” Nature, Vol. 346, No. 6284, 1990, pp. 580-583. http://dx.doi.org/10.1038/346580a0
- J. van der Kaay, J. Wesseling and P. J. van Haastert, “Nucleus-Associated Phosphorylation of Ins(1,4,5)P3 to InsP6 in Dictyostelium,” Biochemical Journal, Vol. 312, No. 3, 1995, pp. 911-917.
- M. Singh and A. D. Krikorian, “Inhibition of Trypsin Activity in Vitro by Phytate,” Journal of Agricultural and Food Chemistry, Vol. 30, No. 4, 1982, pp. 799-800. http://dx.doi.org/10.1021/jf00112a049
- S. R. Tannenbaum and V. R. Young, “Vitamins and Minerals,” In: O. R. Fennema, Ed., Food Chemistry, 2nd Edition, Marcel Dekker, New York, 1985.
- O. Losito, Z. Szijgyarto, A. C. Resnick and A. Saiardi, “Inositol Pyrophosphates and Their Unique Metabolic Complexity: Analysis by Gel Electrophoresis,” PLoS One, Vol. 4, No. 5, 2009, p. e5580. http://dx.doi.org/10.1371/journal.pone.0005580
- H. Marschner, “Mineral Nutrition of Higher Plants,” 2nd Edition, Academic Press, London, 1995.
- T. J. Chiou and S. I. Lin, “Signaling Network in Sensing Phosphate Availability in Plants,” Annual Review of Plant Biology, Vol. 62, No. 1, 2011, pp. 185-206. http://dx.doi.org/10.1146/annurev-arplant-042110-103849
- J. P. Singh, K. Selvendiran, M. Banu, R. Padmavathia and D. Sakthisekaran, “Protective Role of Apigenin on the Status of Lipid Peroxidation and Antioxidant Defense against Hepatocarcinogenesis in Wistar Albino Rats,” Phytomedicine, Vol. 11, No. 4, 2004, pp. 309-314. http://dx.doi.org/10.1078/0944711041495254
- A. M. Shamsuddin, G. Y. Yang and I. Vucenik, “Novel Anti-Cancer Functions of IP6; Growth Inhibition and Differentiation of Human Mammary Cancer Cell Lines in Vitro,” Anticancer Research, Vol. 16, No. 6A, 1996, pp. 3287-3292.
- M. Hirose, S. Fukushima, K. Imaida, N. Ito and T. Shirai, “Modifying Effects of Phytic Acid and G-Oryzanol on the Promotion Stage of Rat,” Carcinogenesis, Vol. 19, 1999, pp. 3665-3670.
- R. Agarwal, H. Mumtaz and N. Ali, “Role of Inositol Polyphosphates in Programmed Cell Death,” Molecular and Cellular Biochemistry, Vol. 328, No. 1-2, 2009, pp. 155-165. http://dx.doi.org/10.1007/s11010-009-0085-6
- L. R. McDowell, “Minerals in Animal and Human Nutrition,” Academic Press, San Diego, 1992.
- M. Gillooly, T. H. Bothwell, J. D. Torrance, A. P. MacPhail, D. P. Derman, W. R. Bezwoda, W. Mills, R. W. Charlton and F. Mayet, “The Effects of Organic Acids, Phytates and Polyphenols on the Absorption of Iron from Vegetables,” British Journal of Nutrition, Vol. 49 No. 3, 1983, pp. 331-342. http://dx.doi.org/10.1079/BJN19830042
- P. R. Heaney, C. M. Weaver and M. L. Fitzsimmons, “Soybean Phytate Content: Effect on Calcium Absorption,” American Journal of Clinical Nutrition, Vol. 53, No. 3, 1991, pp. 745-747.
- K. Johansen and H. D. Poulsen, “Substitution of Inorganic Phosphorus in Pig Diets by Microbial Phytase Supplementation—A Review,” Pig News and Information, Vol. 24, No. 3, 2003, pp. 77N-82N.
- L. Barrientos, J. J. Scott and P. P. N. Murthy, “Specificity of Hydrolysis of Phytic Acid by Alkaline Phytase from Lily Pollen,” Plant Physiology, Vol. 106, No. 4, 1994, pp. 1489-1495. http://dx.doi.org/10.1104/pp.106.4.1489
NOTES
*Corresponding author.