Open Journal of Veterinary Medicine
Vol.05 No.10(2015), Article ID:60415,6 pages
10.4236/ojvm.2015.510028
VEGF System in Dog’s Thymus― Temporal Expression
Fernanda Rodrigues Agreste1, Pedro Primo Bombonato1, Kaique Nogueira2*, Anna Clara dos Santos2, André Rodrigues da Cunha Barreto-Vianna2, Eduardo Maurício Mendes de Lima2
1Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, Brasil
2Faculdade de Agronomia e Medicina Veterinária, Universidade de Brasília, Brasília, Brasil
Email: *kiqnogueira@gmail.com
Copyright © 2015 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 4 September 2015; accepted 18 October 2015; published 21 October 2015
ABSTRACT
Given the importance of the thymus to the immune system and the limited knowledge about its development and involution processes, this study evaluated the expression of the vascular endothelial growth factor, a key angiogenic factor for the formation of vascular tissue bed and modulation of functions related to cell proliferation and migration, hormone synthesis, and immunomodulation. In this study, thymus samples of dog fetuses (30, 40, 50 and 60 days old), and young (180 days old) and adult (365 days old) dogs were submitted to real-time PCR and Western blot. Time- dependent changes were observed for proteins and genes associated with the development of the vascular system, such as VEGF-A, VEGF-C, Flt-1, KDR, FLT-4, and EG-VEGF. This result demonstrates that these system components play different roles during development and involution of the thymus. Possible functions, such as the modulatory effect on vasculogenesis and microenvironment, influence the proliferation and differentiation of thymocytes, T cells maturation, cell-cell interaction and hormone secretion.
Keywords:
Angiogenesis Factor, EG-VEGF, Immune System, Thymus, Vascular Endothelian Growth Factor

1. Introduction
The thymus modulates the activity of other lymphoid organs. Therefore, knowing the mechanisms involved in its development, maturation, and involution is of paramount importance to understand the general activity of the lymphoid system [1] .
The vascular endothelial growth factor (VEGF) is a key angiogenic factor for the formation of vascular tissue bed and modulation of functions related to cell proliferation and migration, hormone synthesis, and immunomodulation. Knowledge of the VEGF expression in the thymus is critical to understand the intercellular interactions in the organ [2] .
The understanding of the vascular behavior, its structure and interferences that occur along the thymic angiogenesis and involution can help to elucidate certain conditions. Autoimmune diseases, thymic tumors and persistence, and other diseases linked to impaired immune systems are some of these peculiarities, which play a very important role in understanding thymic pathophysiology and morphophysiology.
The objective was to determine thymus vascular behavior during development and involution by characterizing the gene and protein temporal expression of EG-VEGF system in dogs’ thymus of different ages using the Western blot and real-time PCR techniques.
2. Materials and Methods
The thymuses of 36 (18 male and 18 female) dogs were collected at the Veterinary Hospital of the College of Veterinary and Animal Science of Universidade de São Paulo and Center for Disease Control of São Paulo. The animals were divided into 6 groups according to age, as follows:
・ Group G1: 30-day-old fetuses (n = 6);
・ Group G2: 40-day-old fetuses (n = 6);
・ Group G3: 50-day-old fetuses (n = 6);
・ Group G4: 60-day-old fetuses (n = 6);
・ Group G5: young dogs, 180 days old (n = 6);
・ Group G6: adult dogs, 365 days old (n = 6).
The thymuses were dissected, fragmented and stored in a freezer at −80˚C until further use. The samples were used to perform Western blot (all groups) and Real Time PCR (for G1, G4, G5 and G6 groups). The Ethics Committee on Animal Use of the Faculty of Veterinary and Animal Science of the Universidade de Sao Paulo approved the study under Protocol #995/2006.
2.1. Real-Time PCR
The total RNA of the samples was extracted following the protocol of the RNA Spin GE® Kit and quantified by spectroscopy (Eppendorf® BioPhotometer). Superscript III (Invitrogen, Carlsbad CA-USA) was used in the reverse transcription step. After transcription, the cDNA samples were kept in freezer at −20˚C until use in real- time PCR.
The sequences used for the preparation of primers (Table 1) are available in the NCBI public database (http://www.ncbi.nlm.nih.gov). The constitutively-expressed GAPDH (glyceraldehyde phosphate dehydrogenase) canine gene was used as internal control of the amplification reactions to normalize the results obtained for the target genes.
The efficiency of the target genes was calculated using the “LinRegPCR” (Ruijter and Ramakers, 2003) software, having as average efficiency the individual amplification curve of each sample. The GAPDH efficiency was calculated using E = 10(−1/slope) [3] [4] where E = efficiency and slope = value obtained from the standard curve generated by the ABI 7500 equipment. Real-Time PCR was performed in the ABIPrism 7500 equipment (Applied Biosystems, Foster, USA).
2.2. Western Blot
Tissue proteins were extracted and homogenized with lysis buffer and protease and phosphatase inhibitors, centrifuged (10,000 RPM, 30 min) and the supernatant was collected to analyze thymus proteins.
The total protein concentration was determined using the Bradford method (1976) (Protein Assay Kit, Bio- Rad, California, USA). After denaturing (five minutes at 95˚C), equal amounts of protein were separated by polyacrylamide gel electrophoresis (8% - 12% SDS-PAGE) and transferred to a nitrocellulose membrane (Amersham Biosciences, Piscataway, NJ, USA). Subsequently, the membrane was blocked in 5% bovine serum albumin (BSA), diluted in Tris buffered saline solution (TBS-T [20 mmol/L Tris/HCl (pH 7.4) and 500 mmol/L

Table 1. Sequences of oligonucleotide initiators used to design the primers.
NaCl] at room temperature for two hours to prevent non-specific binding.
Sequentially, the membrane was incubated with primary antibodies for about 12 hours at 4˚C (Table 2), followed by another incubation with the secondary antibody specific for the primary antibody source during one hour, at room temperature. Protein expression was detected by a chemiluminescence detection system with ECL and developed using photographic film (Film RX_A IBF Filmes do Brasil, São Paulo, Brazil). The intensity of the bands was quantified by the ImageJ software, version 1.47 (NIH imagej.nih.gov/ij, USA).
The antibody binding complex was broken and the membrane was marked again with beta-actin (anti-mouse, SC81178, Santa Cruz Biotechnology, 1:1000), as a constitutive protein to normalize the data obtained by Western blot.
2.3. Statistical Analysis
Data presented as standard deviation and standard error of the mean, were submitted to the Kolmogorov-Smir- nov normality test and when not normally distributed, transformed into logarithms. Statistical analysis of the data expression evaluated during development and involution of the thymus between different age groups, one-way ANOVA at p < 0.05 significance level was applied using the Minitab software.
3. Results
3.1. VEGF mRNA Expression in Thymus of Dogs
The relative expression of VEGF system in the thymus of dogs showed significant time-dependent changes throughout the development and involution processes.
The VEGF-A mRNA expression in the thymus remained constant during fetal development, with a gradual and significant increase in the involution phase (Figure 1(a)), whereas the VEGF-C mRNA expression remained low during development, with no significant increase after birth (Figure 1(b)).
The Flt-1 and VEGF-A mRNA expression remained constant during thymus development; increased in the involution phase, and significantly (p < 0.05) in the later stage of this phase for 365-day-old dogs (Figure 1(c)).
The Flt-4 expression in the thymus was lower compared with other receptors, remaining constant during development, increasing significantly (p < 0.05) during the onset of the involution phase, decreasing again in the later stages of this phase (Figure 1(d)).
The KDR mRNA expression in the thymus increased during development and involution stages, and significantly (p < 0.05) in 365-day-old animals (Figure 1(e)).
Similarly, the relative expression of VEGF and EG-VEGF systems was significantly time-dependent (p <
Table 2. Used antibodies, manufacturer and dilution.
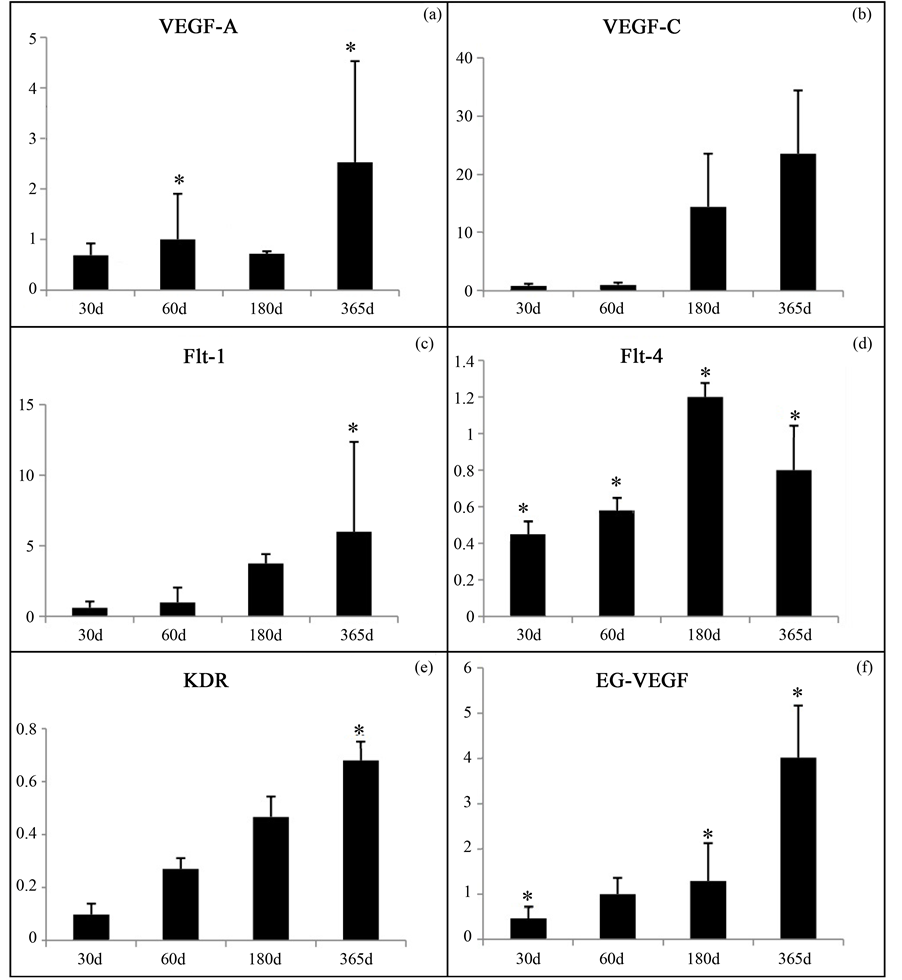
Figure 1. VEGF-A, VEGF-C, Flt-1, Flt-4, KDR and EG-VEGF mRNA expression (AU) in canine thymus at different stages of development and involution. (*p < 0.05).
0.05) during development and involution stages (Figure 1(f)).
3.2. VEGF Protein Expression
Protein expression of all elements of the VEGF system was detected in both development and involution stages.
VEGF-A, VEGF-C and Flt-1 mRNA expression increased over the evaluation period (Figures 2(a)-(c)). Flt-4 also increased until early thymic involution stage, decreasing significantly in 365-day-old dogs (Figure 2(d)). The KDR increased over both stages (Figure 2(e)). Finally, the EG-VEGF displayed high values throughout the development stage, decreasing in the involution stage (Figure 2(f)).
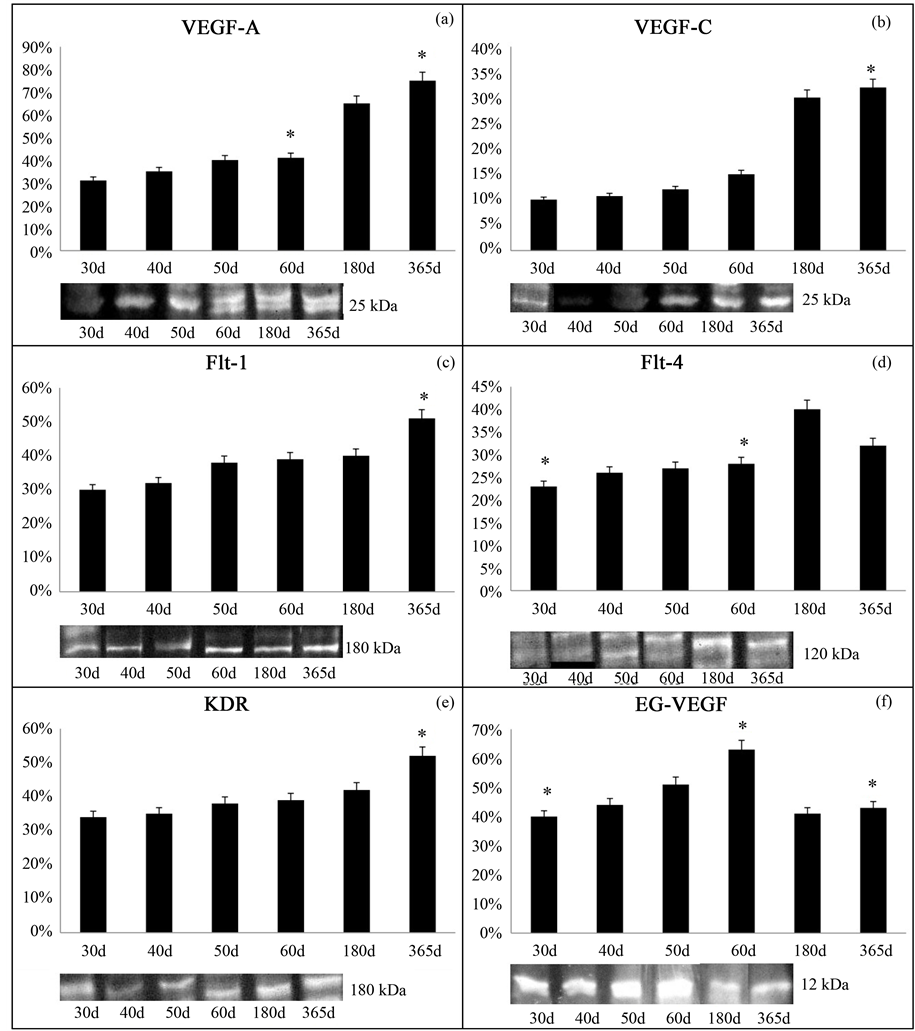
Figure 2. VEGF-A, VEGF-C, Flt-1, Flt-4, KDR and EG-VEGF immunoblots, densities in relation to beta-actin in canine thymus at different stages of development and involution, and molecular weight of each target (kDa). (*p < 0.05).
4. Discussion
VEGF is a mediator of relations between monocytes and endothelial cells, as well as relations between them and the cells of the thymic stroma, especially in the postnatal period. The thymus vascularization is considered the third anatomical compartment of the organ, performing additional functions in thymic development, and responsible for carrying the lymphocyte precursors, derived from the bone marrow, into the body. The reduced expression of VEGF in the epithelial cells leads to a complete change in the vascular pattern [5] . The comparison of these results with those of Agreste et al. (2013) [6] , shows that the expression of the protein and the VEGF-A mRNA in canine thymus system not associated only with cells of the vascular endothelium of the organ, but it may also be related negatively to vascularization.
Protein expression of the VEGF-A system of canine thymus resembled the findings of Cimpean et al. (2008) [7] in humans, which corroborates the hypothesis of active participation of thymic corpuscles in thymopoiesis. Cimpean et al. (2008) [7] observed immunostaining for VEGF-A and its Flt-1 and KDR receptors in human thymic corpuscles of individuals aged from 1 month to 50 years old, in both normal and abnormal physiological conditions. These authors also state that, under normal after birth conditions, VEGF expression decreases rapidly. The immunostaining for this growth factor and its receptors supports the hypothesis that the thymic corpuscles are active structures in the maturation and selection process of lymphocytes in the postnatal period [8] . Furthermore, in the neonatal period, the murine thymus had a higher VEGF expression than in thymus of adult animals, which was also observed in the evaluated canine thymuses [5] .
The protein and mRNA expression of the VEGF system in canine thymus determined by real-time PCR and Western Blot techniques contributed to the better characterization of organ development in the species at different evolution stages. Thus confirming that the expression in the thymus tends to follow the time-dependent growth curve of the organ.
It is possible that the cell populations that express VEGF and its receptors in the thymus are modified after birth. However, specific studies using double labeling/tagging have not been conducted yet. Furthermore, it has been hypothesized that thymopoiesis is regulated via VEGF by both the hematopoietic and epithelial compartment of the thymus [9] .
Thus, mRNA expression displayed a time-dependent profile during development and involution. On the other hand, VEGF protein expression was observed in thymic epithelial cells and thymic corpuscles in all stages. This result reveals that the VEGF-A protein expression in the canine thymus is similar to humans, supporting the hypothesis of active participation of the thymic corpuscles in [7] .
The expression of VEGF-A and VEGFR-1/Flt-1 and VEGFR-2/KDR receptors increased during placental development in the placenta of Pecari tajacu (Santos, 2014). Unlike the VEGF-A expression in the canine thymus, which was higher during the involution stage. On the contrary, the expression of Flt-4, VEGF-C, and D receptor, did not follow the VEGF-A expression pattern, and the decreasing Flt-4 expression during thymic involution is due to its high occurrence in the endothelium. After this phase, the expression of this molecule becomes restricted to lymphatic endothelium in most tissues [10] [11] . Therefore, VEGF-A, Flt-1, and KDR followed the vascular thymus curve and the increasing expression during the organ involution phase might be due to hypoxia. During the involution stage, the blood supply decreased in the organ, increasing the expression of VEGF-A and its receptors. This increase in response to hypoxia revealed the opposite direction of the VEGF-A system when the vascularization of the thymus decreased.
VEGF-C and D exerted angiogenic effect via KDR and Flt-4, and lymphangiogenic, via Flt-4. The absolute or relative expression levels of these receptors led to believe that VEGF-C/D promoted the angiogenic or lymphan- giogenic effect. The relationship between VEGF-C and these receptors may explain the increased expression of this factor in the thymus involution stage [9] [12] -[14] .
In human and mouse placenta, EG-VEGF played an important role in placentation, proliferation, migration and selective fenestration in the endothelial cells of endocrine glands [15] . However, EG-VEGF expression in other organs such as liver, kidney and intestine suggested that EG-VEGF was not a factor limited to endocrine glands [16] [17] .
5. Conclusion
The results suggest that the VEGF system plays different roles during development and involution of the thymus. Possible functions include a modulatory effect on the microenvironment and vasculogenesis, influencing the pro- liferation and differentiation of thymocytes, maturation of T cells, cell-cell interaction and hormone secretion.
Conflict of Interest
None.
Authors’ Contribution
AFR, PPB, ACS and KN collected and analyzed the data and drafted the manuscript. ARCBV and EMML conceived the project, supervised data collected, oversaw composition of the manuscript and obtained funding. All authors read and approved the final manuscript.
Cite this paper
Fernanda RodriguesAgreste,Pedro PrimoBombonato,KaiqueNogueira,Anna Clara dosSantos,André Rodrigues da CunhaBarreto-Vianna,Eduardo Maurício Mendes deLima, (2015) VEGF System in Dog’s Thymus—Temporal Expression. Open Journal of Veterinary Medicine,05,211-217. doi: 10.4236/ojvm.2015.510028
References
- 1. Sinkora, J., Rehakova, Z., Sinkora, M., Cukrowska, B. and Tlaskalova-Hogenova, H. (2002) Early Development of Immune System in Pigs. Veterinary Immunology, 87, 301-306.
http://dx.doi.org/10.1016/S0165-2427(02)00056-9 - 2. Kiseleva, E.P., Krylov, A.V., Lyudyno, V.I. and Suvorov, A.N. (2005) Effect of VEGF on Mouse Thymocyte Proliferation and Apoptosis in Vitro. Bulletin of Experimental Biology and Medicine, 139, 576-579.
http://dx.doi.org/10.1007/s10517-005-0349-4 - 3. Bustin, S.A. (2000) Absolute Quantification of mRNA Using Real-Time Reverse Transcription Polymerase Chain Reaction Assays. Journal of Molecular Endocrinology, 25, 169-193.
http://dx.doi.org/10.1677/jme.0.0250169 - 4. Rasmussen, R. (2001) Quantification on the Light Cycler. In: Meuer, S., Wittwer, C. and Nakagawara, K., Eds., Rapid Cycle Real-Time PCR, Methods and Applications, Springer Press, Heidelberg, 21-34.
http://dx.doi.org/10.1007/978-3-642-59524-0_3 - 5. Cuddihy, A.R., Ge, S., Zhu, J., et al. (2009) VEGF-Mediated Cross-Talk within the Neonatal Murine Thymus. Blood, 113, 2723-2731.
http://dx.doi.org/10.1182/blood-2008-06-162040 - 6. Agreste, F.R., Moriconi, P.R., Barreto-Vianna, A.R.C., Santos-Leonardo, A., Bombonato, P.P. and Lima, E.M.M. (2013) Is the Vascular Growth Curve of Thymus of Dogs (Canis familiaris) Time-Dependent? Australian Journal of Basic and Applied Science, 7, 334-342.
- 7. Cimpean, A.M., Raica, M., Encica, S., Cornea, R. and Bocan, V. (2008) Immunohistochemical Expression of Vascular Endothelial Growth Factor A (VEGF), and Its Receptors (VEGFR1, 2) in Normal and Pathologic Conditions of the Human Thymus. Anatomischer Anzeiger, 190, 238-245.
http://dx.doi.org/10.1016/j.aanat.2007.05.003 - 8. Watanabe, N., Wang, Y.H., Lee, H.K., et al. (2005) Hassall’s Corpuscles Instruct Dendritic Cells to Induce CD4+CD25+ Regulatory T Cells in Human Thymus. Nature, 436, 1181-1185.
http://dx.doi.org/10.1038/nature03886 - 9. Olsson, A.K., Dimberg, A., Kreuger, J. and Claesson-Welsh, L. (2006) VEGF Receptor Signalling? In Control of Vascular Function. Nature Reviews: Molecular Cell Biology, 7, 359-371.
http://dx.doi.org/10.1038/nrm1911 - 10. Ji, R.C. (2009) Lymph Node Lymphangiogenesis, a New Concept for Modulating Tumor Metastasis and Inflammatory Process. Histology and Histopathology, 24, 377-384.
- 11. Sabattini, S. and Bettini, G. (2009) An Immunohistochemical Analysis of Canine Haemangioma and Haemangiosarcoma. Journal of Comparative Pathology, 140, 158-168.
http://dx.doi.org/10.1016/j.jcpa.2008.10.006 - 12. Shibuya, M. (2001) Structure and Dual Function of Vascular Endothelial Growth Factor Receptor-1 (Flt-1). The International Journal Biochemistry & Cell Biology, 33, 409-420.
http://dx.doi.org/10.1016/S1357-2725(01)00026-7 - 13. Takahashi, H. and Shibuya, M. (2005) The Vascular Endothelial Growth Factor (VEGF)/VEGF Receptor System and Its Role under Physiological and Pathological Conditions. Clinical Science, 109, 227-241.
http://dx.doi.org/10.1042/CS20040370 - 14. Ribatti, D., Nico, B., Crivellato, E., Roccaro, A.M. and Vacca, A. (2007) The History of the Angiogenic Switch Concept. Leukemia, 21, 44-52.
http://dx.doi.org/10.1038/sj.leu.2404402 - 15. LeCouter, J., Kowalski, J., Foster, J., Hass, P., Zhang, Z., Dillard-Telm, L. and Ferrara, N. (2001) Identification of an Angiogenic Mitogen Selective for Endocrine Gland Endothelium. Nature, 412, 877-884.
http://dx.doi.org/10.1038/35091000 - 16. Hoffmann, P., Feige, J.J. and Alfaidy, N. (2006) Expression and Oxygen Regulation of Endocrine Gland-Derived Vascular Endothelial Growth Factor/Prokineticin-1 and Its Receptors in Human Placenta during Early Pregnancy. Endocrinology, 147, 1675-1684.
http://dx.doi.org/10.1210/en.2005-0912 - 17. Hoffmann, P., Feige, J.J. and Alfaidy, N. (2007) Placental Expression of EG-VEGF and Its Receptors PKR1 (Prokineticin Receptor-1) and PKR2 throughout Mouse Gestation. Placenta, 28, 1049-1058.
http://dx.doi.org/10.1016/j.placenta.2007.03.008
NOTES

*Corresponding author.