Open Journal of Veterinary Medicine
Vol.4 No.1(2014), Article ID:41943,5 pages DOI:10.4236/ojvm.2014.41001
Newcastle Disease Virus Isolation and Its Prevalence in Uganda Poultry Farms
1Central Diagnostic Laboratory, Department of Pharmacy Clinics and Comparative Medicine, College of Veterinary Medicine Animal Resources and Biosecurity, Makerere University, Kampala, Uganda
2Indian Veterinary Research Institute (IVRI), Central Avian Research Institute (CARI), Izatnagar, India
Email: *kicelandy@gmail.com
Copyright © 2014 Keneth Iceland Kasozi et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. In accordance of the Creative Commons Attribution License all Copyrights © 2014 are reserved for SCIRP and the owner of the intellectual property Keneth Iceland Kasozi et al. All Copyright © 2014 are guarded by law and by SCIRP as a guardian.
Received December 17, 2013; revised January 2, 2014; accepted January 8, 2014
KEYWORDS
Newcastle Disease Virus; Chicken Embryo Virus Isolation; Poultry Farmers in Uganda; Avain Viruses in Africa; Animal Production Challenges
ABSTRACT
The present research work was carried out to isolate and identify Newcastle disease virus (NDV) by using haemagglutination inhibition (HI) test and HA-HI virus isolation, embryonated eggs (EE) and chicken embryo fibroblasts (CEF). A total of 95 clinical (blood, tracheal and cloacal swabs) and post-mortem (brain, lung, colon and spleen) samples were collected from chickens of field outbreaks of suspected Newcastle disease virus (NDV). The HI and HA-HI were employed to detect NDV in tissue homogenates of all the clinical and post-mortem samples as well as laboratory samples (AF and ICF). Among the four different types of post-mortem samples, virus isolation rate was found to be low in body organs. In CEF cell culture system, the rate of virus isolation from all the aforesaid samples was found to be at 100% with the exception of serum samples; while in tracheal and cloacal swabs, it was at 90%; while in serum, it was at 10%, in all clinical cases. The isolation rate of NDV was higher in CEF culture system (66.7%) compared to that of avian embryos (33.3%). Samples were inoculated and the allantoic fluid (AF) of the dead embryos and the infected culture fluid (ICF) of the CEF were harvested at 24 to 96 hours of the post-infection, respectively, which revealed that the virulent strain of NDV is highly prevalent in the region. The prevalence of NDV was established at 1.1%, 2.1% and 4.2% using HA-HI, EE, and CEF methods. Rapid detection and identification of the virus are crucial for the effective control of the disease as conventional diagnostic methods such as virus isolation on embryonated eggs followed by serological identification in haemagglutination-inhibition test are laborious and time-consuming. The speed of the diagnosis can be considerably increased by using methods based on molecular biology, e.g. reverse transcription—polymerase chain reaction. However, the genetic variability of APMV-1 isolates should be considered carefully as the potential cause for false negative results of genetic-based laboratory tests.
1. Introduction
Newcastle disease virus (NDV), also known as avian pneumoencephalitis, is an important production disease of poultry having a serious impact on national and international markets worldwide. NDV is a non-segmented, negative-strand RNA virus belonging to genus Avulavirus in the family Paramyxoviridae. Virulent NDV strains [1] cause a severe respiratory and neurologic disease in poultry worldwide and naturally occurring avirulent NDV strains have been successfully used as vaccines for more than 70 years [1]. Development of reverse genetic systems has allowed manipulation of the genome of NDV [2-4] and generation of recombinant NDVs that express foreign proteins from added genes [5,6]. Several groups have used NDV reverse genetic systems to develop vaccines against AIV subtypes H5 [7,8] and H7 [9].
The ND virus is shed during the incubation period of the disease, the clinical phase and for a period during recovery. The rate of spread of NDV is determined by the organs in which the virus multiples. NDV is present in air respired through the trachea, as well as the cloacal discharges in infected fowls. Live poultry, hatcheries, carcasses and offal [10], poultry shows and markets are important in the spread of the virus, but the current practice of moving exotic birds around the world has helped the international transport of the disease.
Background in Uganda
A prospective study of risk factors associated with outbreaks of Newcastle disease (ND) in indigenous freerange chickens was carried out in three agro-ecological zones in eastern Uganda. Multivariate survival analyses were performed to identify the risk factors for Newcastle disease outbreaks. Although the survival time [11] against ND was longer (>100 days) in the dry compared to the rainy season, none of the factors investigated was significantly associated with ND [11]. However, purchasing of restocking chickens [11] from the market and neighborhood (hazard ratio [HR] = 1.79; 95% confidence interval [CI] 0.89 - 3.60), the presence of migratory wild birds (HR = 1.70; 95% CI 0.65 - 4.48) and being in agro-ecological zone 1 (HR = 1.48; 95% CI 0.66 - 3.36) showed a positive but non-significant association with risk for ND [11]. In another study, housed chickens attained 100% protection to Newcastle disease with Mean ND antibody HI log2 titre of 6.19 ± 1.3 compared to 89% protection by unhoused chickens with Mean ND antibody HI log2 titre of 5.40 ± 2.5. The benefits and adaptability of these poultry health interventions are discussed in view of the rural poultry management system in Uganda [12].
2. Materials and Methods
A study poultry population of 95 chicken, submitted at the college of veterinary medicine animal resources and biosecurity (CoVAB) by farmers from June to September 2013 was included in the study. The farmers came from central Uganda (Kampala, Mukono, Kayunga, Kiboga, Mpigi, Wakiso, Rakai and Kyankwanzi districts) and all chicken presented was screened for virus shadding and antibody titers by use of the HA-HI tests followed by virus isolation from the tissues using 11-day-old chick embryonated eggs and chicken embryonated fibroblasts (CEF). Serum samples representing 100%, were collected from all chicken, using the wing veins. The serum samples were tested for ND antibody titres to establish the pre-vaccination sero-status using the Haemagglutination Inhibition (HI) test [13]. The serum samples were stored at 4˚C before testing for protection against ND by the HI test [13]. Virus isolation was further carried out from cloacal, trachea, spleen, lung, liver and heart following post mortem from suspected suspected ND cases.
2.1. Virus Identification
HA activity detected in bacteriologically sterile fluids harvested from inoculated eggs was due to the presence of any of the ten subtypes of APMV (including NDV) or 16 haemagglutinin subtypes of influenza A viruses. NDV was confirmed by the use of specific antiserum in a haemagglutination inhibition (HI) test by using chicken antiserum that had been prepared against one of the strains of NDV [14]. The antibody levels of serum samples collected from chickens vaccinated with NDVs were evaluated by hemagglutination (HA), HA inhibition (HI), virus neutralization (VN) and enzyme-linked immunesorbent assays (ELISA) using standard protocols [15].
2.2. Biological Characterization of NDV
The pathogenicities of the strain were evaluated by the mean death time (MDT) test in embryonated chicken eggs. According to the OIE guidelines, an NDV strain [1,3] is considered lentogenic or avirulent, if the MDT value is >90 h [16].
2.3. Quality Control
Since chicken sera rarely give nonspecific positive reactions in the HI test and any pretreatment of the sera was unnecessary since all serum samples handled were from chicken only for the HA-HI tests. Chicken blood was taken from unvaccinated birds monitored regularly and shown to be free from antibodies to NDV. The chicken blood cells were washed three times in phosphate buffered solution (PBS) before use as a 1% (packed cell v/v) suspension.
NDV harvesting was carried out in a biosafety cabinet (BSC 2) to avoid spread of the virus in the environment and protective gear was used to protect personnel handling the samples. The validity of results was assessed against a negative control serum, which did not give a titre >1/4 (>2 2 or >log 2 when expressed as the reciprocal), and a positive control serum [14] for which the titre was within one dilution of the known titre.
3. Results
3.1. Hemagglutination-Hemagglutination Inhibition (HA-HI) and NDV Sero-Prevalence and It’s Isolation
Clinical cases were submitted to the central diagnostic laboratory (CDL) from which postmortem (PM) chicken samples were collected for ND virus isolation whenever judged appropriate by the pathologists CoVAB. The prevalence of NDV by embryonated egg isolation was found to be at 2.1% and the sero-prevalence of the disease established at 1.1% using HA-HI titers. Viral isolation using CEF was over 60% than using EE (40%) which gave a viral prevalence of 4.2% and 2.1 % respectively. Only cases that presented a history of Newcastle were considered for the isolation especially after gathering the relevant clinical history on the farm management and progress of the disease and the results are summarized in Table 1.
3.2. Viral Characterization and Isolation from the Submitted Chicken Birds
Hemagglutination (HA) was carried out on allantoic fluid (AF) and the virulence of the viral strain in the region was found to be the virulent strain of NDV being prevalent in the region. Following inoculation of both clinical and PM samples, the allantoic fluid was harvested and hemagglutination (HA) tests carried out for viral isolation. AF of HA titers 1 < 8 were subjected to second passage, and the activity repeated from which the virus was further confirmed.
Viral characterization showed that the prevalent strain was the virulent strain of NDV as it had a mean death time less than 90 h [16] as shown in Table 2.
4. Discussion
All the samples analyzed in this study were from the central region due to its proximity to the central diagnos-
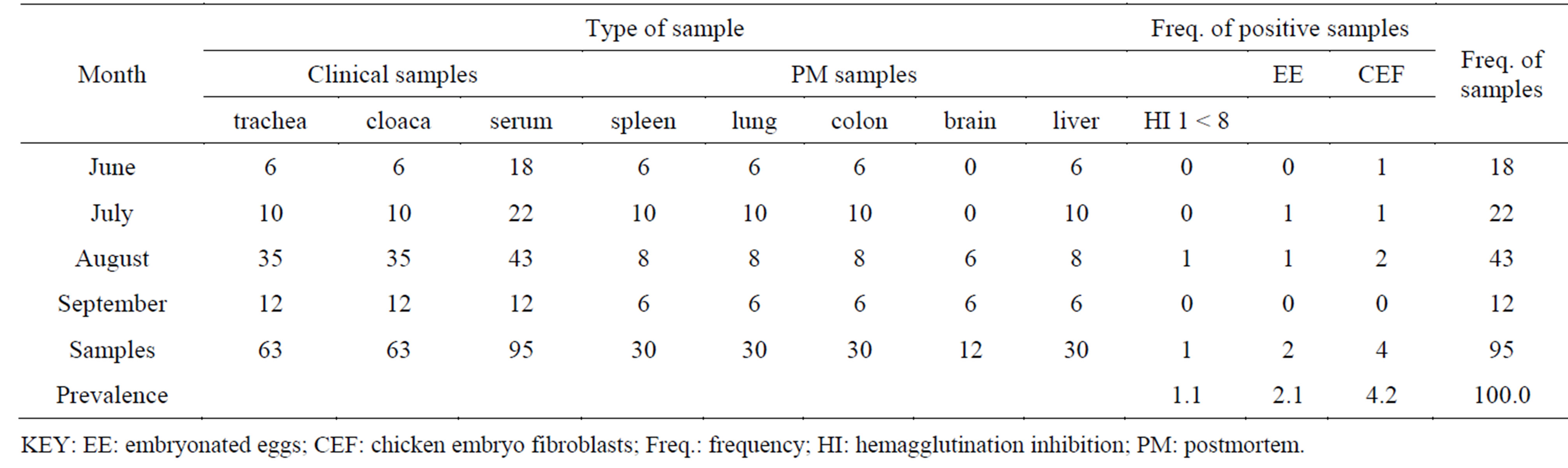
Table 1. Frequency of samples submitted, HI and virus isolation prevalence.

Table 2. Detection of Newcastle disease virus from the clinical and post-mortem samples of naturally infected chickens.
tic laboratory at the CoVAB. The sero-preverance of ND virus within the study area was found to be at 1.1%, virus isolation by embryonated eggs at 2.1%, and chicken embryo fibroblasts (CEF) at 4.2%. This would be due to the good vaccination status of most of the farms within the study area accounting for the low titers registered in this study. Simillar results among the clinical and postmortem samples i.e. inoculum of cloacal swab and colon (intestine) showed HA and HI activities (Tables 1 and 2) just as it was in [17]. This might be due to the presence of low concentrations of free NDV in both types of samples during early and advanced stage of the disease. The anti-NDV hyperimmune serum [17] revealed complete inhibition of the 4 haemagglutination unit of each isolate of viruses isolated from broiler and layer chickens present in the laboratory samples (AF and ICF), which were the breeds of chicken analyzed for this study. The HI test results of the present study directly support the findings of [18,19] who confirmed NDV by HI test. The failure of virus isolation from the blood samples at postinfection might be due to the lodgment of the virus in different tissues of the infected birds of post infection. The reduced rate of virus isolation in avian embryo may be due to the presence of maternal antibody.
In vaccinated flocks that were being monitored serologically, it was possible to identify anamnestic responses as the result of a challenge infection with field virus [20], but great care was exercised as variations would occur from other causes. The spread of ND virus in communities has also been propagated by vectors such as P. cuprina and M. domestica [21], F. canicularis and Fannia femoralis (Stein) [22] which are highly prevalent in many developing countries especially the tropics thus contributing to the high incidence of NDV within these communities. Viral isolation was highest (80%) in in both cloacal and tracheal over that in the body tissues (20%), thus leading to high rates of shading and spread of the infection within communities. In the HI test, some level of cross-reactivity was observed among the various avian paramyxovirus serotypes. Cross-reactivity was observed between APMV-1 and APMV-3 viruses (particularly with the psittacine variant of APMV-3 [14], commonly isolated from pet or exotic birds) or APMV-7. The risk of mistyping an isolate was greatly reduced by using a panel of reference sera or monoclonal antibodies (MAbs) specific for APMV-1, APMV-3 and APMV-7 [14].
5. Conclusion and Recommendations
Newcastle disease is one of the most important infectious diseases of poultry. The rapid detection and the identification of the virus are crucial for the effective control of the disease. Conventional diagnostic methods such as the virus isolation on embryonated eggs followed by the serological identification and the haemagglutination-inhibition test are laborious and time-consuming. The speed of the diagnosis can be considerably increased by using methods based on the molecular biology, e.g. reverse transcription—polymerase chain reaction (RT-PCR).
At present, RT-PCR-based techniques for the detection and typing (pathotyping and genotyping) of APMV-1 RNA in the allantoic fluid of inoculated fowl eggs are becoming increasingly common in diagnostic laboratories. However, the genetic variability of APMV-1 isolates should be considered carefully as potential cause for false negative results of genetic-based laboratory tests.
Acknowledgements
The study was carried out as part of the Japanese Technical Assistance to Improve the National Diagnostic Capacity for Animal Disease Control in Uganda. The study was funded in part by the Japan International Cooperation Agency.
REFERENCES
- B. Nayak, S. N. Rout, S. Kumar, M. S. Khalil, M. M. Fouda, L. E. Ahmed, K. C. Earhart, D. R. Perez, P. L. Collins and S. K. Samal, “Immunization of Chickens with Newcastle Disease Virus Expressing H5 Hemagglutinin Protects against Highly Pathogenic H5N1 Avian Influenza Viruses,” PLoS One, Vol. 4, No. 8, 2009, Article ID: e6509. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2716524/
- S. Krishnamurthy, Z. Huang and S. K. Samal, “Recovery of a Virulent Strain of Newcastle Disease Virus from Cloned cDNA: Expression of a Foreign Gene Results in Growth Retardation and Attenuation,” Virology, Vol. 278, No. 1, 2000, pp. 168-182. http://www.ncbi.nlm.nih.gov/pubmed/11112492 http://dx.doi.org/10.1006/viro.2000.0618
- T. Nakaya, J. Cros, M. S. Park, Y. Nakaya, H. Zheng, et al., “Recombinant Newcastle Disease Virus as a Vaccine Vector,” Journal of Virology, Vol. 75, No. 1, 2001, pp. 1868-1873.
- Z. Huang, S. Elankumaran, A. Panda and S. K. Samal, “Recombinant Immunization of Primates with a Newcastle Disease Virus-Vectored Vaccine Via the Respiratory Tract Induces a High Titer of Serum Neutralizing Antibodies against Highly Pathogenic Avian Influenza Virus,” Journal of Virology, Vol. 8, No. 1, 2003, pp. 11560- 11568.
- J. M. DiNapoli, L. Yang, A. Suguitan Jr., S. Elankumaran, D. W. Dorward, et al., “Immunization of Primates with a Newcastle Disease Virus-Vectored Vaccine Via the Respiratory Tract Induces a High Titer of Serum Neutralizing Antibodies against Highly Pathogenic Avian Influenza Virus,” Journal of Virology, Vol. 8, No. 1, 2007, 11560- 11568. http://dx.doi.org/10.1128/JVI.00713-07
- Z. Huang, S. Elankumaran, A. S. Yunus and S. K. Samal, “A Recombinant Newcastle Disease Virus (NDV) Expressing VP2 Protein of Infectio Us Bursal Disease Virus (IBDV) Protects against NDV and IBDV,” Journal of Virology, Vol. 7, No. 8, 2004, pp. 10054-10063. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC514986/
- J. Ge, G. Deng, Z. Wen, G. Tian, Y. Wang, et al., “Newcastle Disease Virus Based Live Attenuated Vaccine Completely Protects Chickens and Mice from l Ethal Challenge of Homologous and Heterologous H5N1 Avian Influenza Viruses,” Journal of Virology, Vol. 81, No. 1, 2007, pp. 150-158. http://dx.doi.org/10.1128/JVI.01514-06
- J. Veits, D. Wiesner, W. Fuchs, B. Hoffmann, H. Granzow, et al., “Newcastle Disease Virus Expressing H5 Hemagglutinin Gene Protects Chickens against Newcastle Disease and Avian Influenza,” Proceedings of the National Academy of Sciences of the United States of America, Vol. 103, No. 21, 2006, pp. 8197-8202. http://dx.doi.org/10.1073/pnas.0602461103
- J. Steel, S. V. Burmakina, C. Thomas, E. Spackman, A. García-Sastre, D. E. Swayne and P. Palese, “A Combination in-ovo Vaccine for Avian Influenza Virus and Newcastle Disease Virus,” 2006. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2394284/
- Newcastle, “Newcastle Disease,” University of Reading, Reading, 2002. http://www.veeru.reading.ac.uk/comp2/Poultryweb/disease/newc/newc1.htm
- M. O. Otim, E. K. Kabagambe, G. M. Mukiibi, H. Christensen and M. Bisgaard, “A Study of Risk Factors Associated with Newcastle Disease Epidemics in Village Free-Range Chickens in Uganda,” Tropical Animal Health and Production, Vol. 39, No. 1, 2007, pp. 27-35. http://www.ncbi.nlm.nih.gov/pubmed/17941485
- J. Illango, G. Mukiibi-Muka, P. P. Abila, G. Musisi and A. Etoori, “The Value of the Newcastle Disease I-2 Thermostable Vaccine Use in the Rural Free-Range Poultry Management System in Uganda,” 2012.
- W. H. Allan and R. E. Gough, “A Standard Heamagglutination Inhibition Test for Newcastle Disease,” Veterinary Records, Vol. 95, No. 7, 1974, pp. 147-149. http://dx.doi.org/10.1136/vr.95.7.147
- OIE, “Version Adopted by the World Assembly of Delegates of the OIE,” Newcastle Disease, 2012. www.oie.int/fileadmin/Home/eng/.../2.03.14_NEWCASTLE_DIS.pdf
- J. C. Hierholzer and R. A. Killington, “Virus Isolation and Quantitation,” In: B. W. Mahy and H. I. Kangro, Eds., Virology Methods Manual Academic Press, London, 1996, pp. 25-46.
- OIE, “Manual of Diagnostic Tests and Vaccines for Terrestrial Animals,” Newcastle Disease, 2004. http://web.oie.int/fr/normes/mmanual/A_00038.htm
- M. H. Haque, M. T. Hossain, M. T. Islam, M. A. Zinnah, M. S. R. Khan and M. A. Islam, “Isolation and Detection of Newcastle Disease Virus from Field Outbreaks in Broiler and Layer Chickens by Reverse Transcription Polymerase Chain Reaction,” 2012. www.banglajol.info/index.php/BJVM/article/download/9618/7133
- T. B. Manin, L. O. Shcherbakova, I. A. Bochkov, V. V. El’nikov, I. P. Pchelkina, S. K. Starov and V. V. Drygin, “Characteristics of Field Isolates of Newcastle Disease Virus Isolated in the Course of Outbreaks in the Poultry Plant in the Leningrad Region in 2000,” Voprosy Virusologii, Vol. 47, No. 6, 2002, pp. 41-43.
- B. S. Seal, M. G. Wise, J. C. Pedersen, D. A. Senne, R. Alvarez, M. S. Scott, D. J. King, Q. Yu and D. R. Kapczynski, “Genomic Sequences of Low-Virulence Avian Paramyxovirus-1 (Newcastle Disease Virus) Isolates Obtained from Live-Bird Markets in North America Not Related to Commonly Utilized Commercial Vaccine Strains,” Veterinary Microbiology, Vol. 106, No. 1-2, 2005, pp. 7- 16. http://www.ncbi.nlm.nih.gov/pubmed/15737469. http://dx.doi.org/10.1016/j.vetmic.2004.11.013
- D. J. Alexander and W. H. Allan, “Newcastle Disease Virus Pathotypes,” Avian Pathology, Vol. 3, No. 4, 1974, pp. 269-278.
- S. Chakrabarti, D. J. King, C. J. Cardona, C. Afonso, D. R. Kuney, D. Swayne and A. C. Gerry, “Detection and Isolation of Exotic Newcastle Disease Virus from FieldCollected Flies,” Journal of Medical Entomology, Vol. 44, No. 5, 2007, pp. 840-844. http://www.bioone.org/doi/abs/10.1603/0022-2585%282007%2944%5B840:DAIOEN%5D2.0.CO%3B2
- W. M. Rogoff, E. C. Carbrey, R. A. Bram, T. B. Clark and G. H. Gretz, “Transmission of Newcastle Disease Virus by Insects: Detection in Wild Fannia spp. (Diptera: Muscidae),” Journal of Medical Entomology, Vol. 12, No. 2, 1975, pp. 225-227. http://www.researchgate.net/publication/5928524_Detection_and_isolation_of_exotic_Newcastle_disease_virus_from_field-collected_flies
NOTES
*Corresponding author.