World Journal of Neuroscience
Vol.04 No.05(2014), Article ID:50779,10 pages
10.4236/wjns.2014.45043
Thymosin β4 Improves Neurological Outcome and Enhances Induced Oligodendrogenesis in the Rat after ICH
Dongmei Yang1, Yuxia Han1, Michael Chopp2,3, Donald M. Seyfried1*
1Departments of Neurosurgery, Henry Ford Health System, Detroit, MI, USA
2Departments of Neurology, Henry Ford Health System, Detroit, MI, USA
3Department of Physics, Oakland University, Rochester, MI, USA
Email: *nsdos@neuro.hfh.edu
Copyright © 2014 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 18 August 2014; revised 19 September 2014; accepted 25 September 2014
ABSTRACT
Thymosin β4 (Tβ4), a G-actin binding protein, has diverse biological functions. This study tested the effects of Tβ4 on oligodendrogenesis in a rat model of intracerebral hemorrhage (ICH). ICH was induced by stereotactic injection of 100 µm of autologous blood into the striatum in 32 male Wistar rats. The rats were randomly divided into four groups: 1) saline control group (n = 8); 2) 3 mg/kg Tβ4-treated group (n = 8); 3) 6 mg/kg Tβ4-treated group (n = 8); and 4) 12 mg/kg Tβ4- treated group (n = 8). Tβ4 or saline was administered intraperitoneally starting at 24 h post ICH and then every 3 days for 4 additional doses. The neurological functional outcome was evaluated by behavioral tests (i.e., modified Neurological Severity Score and corner turn test) at multiple time points after ICH. Animals were sacrificed at 28 days post ICH, and histological studies were completed. Tβ4 treatment improved neurological functional recovery significantly and increased actively proliferating oligodendrocytic progenitor cells and myelinating oligodendrocytes in the ICH-affected brain tissue, compared with the saline-treated group. The high-dose treatment of Tβ4 showed better restorative effects compared with the low-dose treatment. Tβ4 treatment enhanced ICH-induced oligodendrogenesis that may contribute to the enhanced functional recovery after ICH. Further investigation is warranted to determine the associated underlying mechanisms of Tβ4 treatment for ICH.
Keywords:
Thymosin β4, Neurogenesis, Oligodendrogenesis, Intracerebral Hemorrhage

1. Introduction
Current clinical treatment strategies for intracerebral hemorrhage (ICH), a particularly devastating form of stroke, emphasize improved functional recovery of the patient [1] . The neurological functional deficits after ICH are caused by damage to both gray matter (neurons and astrocytes) and white matter (axons and myelin-pro- ducing oligodendrocytes) [2] [3] . Injured oligodendrocytes do not form new myelin sheaths and mature oligodendrocytes do not proliferate in the adult brain [4] . Loss of oligodendrocytes and their myelin impairs axonal function [5] . Therefore, increased oligodendrogenesis could be a crucial step during the recovery process as new oligodendrocytes from oligodendrocytic precursor cells (OPCs) are required to form myelin sheaths for sprouting axons after ICH [2] [6] .
Thymosin β4 (Tβ4) has multiple biological functions in addition to its actin-binding properties [7] - [10] . It inhibits inflammation and apoptosis, and promotes regeneration in the skin, cornea and heart [11] [12] . In murine models of experimentally induced myocardial infarction (MI), exogenous Tβ4 promotes the survival of cardiomyocytes and restores cardiac function [13] . These cardioprotective effects may be attributed to Tβ4 stimulating vasculogenesis, angiogenesis and arteriogenesis by mobilizing, recruiting and promoting the differentiation of progenitor cells [14] . Tβ4 is widely distributed in the mammalian central nervous system [15] [16] . It is expressed in most neural cell types of the developing brain and in a subset of neurons and microglia [17] - [19] . Exogenous Tβ4 has been demonstrated to benefit the recovery of animals after ischemic stroke, traumatic brain injury (TBI), and experimental autoimmune encephalomyelitis (EAE) by stimulating OPCs [18] [20] [21] . In this study, we employ an animal model of ICH to investigate the effect of Tβ4 on functional recovery and particularly on oligodendrogenesis.
2. Materials and Methods
2.1. Experimental Model
All studies were approved by the Institutional Animal Care and Use Committee of Henry Ford Hospital (IACUC No. 1243). Young adult male Wistar rats (n = 32, weighing 270 - 330 g) were used. Primary ICH was induced by direct infusion of autologous whole blood into the striatum region adjacent to the subventricular zone (SVZ) [22] - [24] . The autologous blood-injection model of ICH has been well-defined and reproducible in our laboratory.
2.2. Experimental Protocols
At 24 h post-ICH, the animals were randomly assigned into four groups: 1) saline control group (n = 8); 2) 3 mg/kg Tβ4-treated group (n = 8); 3) 6 mg/kg Tβ4-treated group (n = 8); and 4) 12 mg/kg Tβ4-treated group (n = 8). The Tβ4 provided by RegeneRx Biopharmaceuticals, Inc. (Bethesda, MD) is a synthetic copy of the naturally occurring 43-amino acid peptide. Tβ4 was dissolved in saline and administered intraperitoneally starting at 24 h post-ICH and then every 3 days at 4 additional identical doses. The same volume of saline was used as a control group using the identical protocol as those treated with Tβ4. This dose of 6 mg/kg Tβ4 has been shown to increase neurogenesis in ischemic brain and TBI in a previous animal study [25] [26] .
To detect proliferating cells, 5-bromo-2-deoxyuridine (BrdU, 100 mg/kg; Sigma-Aldrich Co., St. Louis, MO) was injected intraperitoneally into rats once daily for 14 days, starting 1 day after injury. All rats were sacrificed at 28 days after ICH. For testing HP expression, 6 additional animals were added. Three rats were treated with 12 mg/kg Tβ4 and 3 rats were treated with saline. These rats were sacrificed at 7 days for ELISA.
2.3. Neurobehavioral Testing
Functional outcome was assessed using a cornering test and mNSS at days 1, 4, 7, 14, 21 and 28 after ICH. The cornering test measures the number of times that an animal turns to the right or left when placed in a corner (the normal state being 50%) [27] . The mNSS-a composite of motor, sensory, balance, and reflex measures-is used to assess neurological functions using a 0 - 18 scale, with higher scores implying greater neurological injury [28] .
2.4. Immunohistochemistry
At the 4th week post ICH and after completed functional tests, the animals were anesthetized using ketamine (44 mg/kg) and xylazine (13 mg/kg intraperitoneally) and perfused transcardially with PBS, followed by 4% paraformaldehyde in PBS. The intact brain was immediately removed and immersed in fixative overnight. The brains were embedded in paraffin and were cut into seven equally spaced (2 mm) coronal blocks.
Six micrometer-thick sections from the block (bregma 0.1 mm to - 0.86 mm) were labeled histochemically for BrdU (a marker for proliferation cells), 2″, 3″-cyclic nucleotide 3′-phosphodiesterase (CNPase, a marker of mature oligodendrocytes), and chondroitin sulfate proteoglycan (NG-2, a marker of oligodendrocyte progenitor cells).
Briefly, tissue sections were deparaffinized in changes of xylene and rehydrated in decreasing concentrations of ethanol. Sections were boiled in 10 mM citric acid buffer (pH 6) for 10 min. After washing with PBS, sections were incubated in 2.4 N HCl for 20 min at 37˚C. Sections were then incubated with 1% BSA containing 0.3% Triton-X-100 in PBS, followed by incubation with BrdU (1:200, Monoclonal, incubated overnight at 4˚C [Dako, Carpinteria, CA]), CNPase (1:200, Monoclonal, incubated for 2 days at 4˚C [Chemicon, Temecula, CA]), and NG-2 (1:800, Polyclonal rabbit, incubated overnight at 4˚C [Chemicon, Temecula, CA]). Fluorescein isothiocyanate (FITC)-conjugated antibody (1:200 [Jackson ImmunoResearch Laboratories, Inc., West Grove, PA]) was added to sections at room temperature for 1 h. Double immunostaining for CNPase and BrdU in rat brain sections was conducted by first staining the sections for CNPase as described above. Next, the sections were treated with 2 N HCl for 1 h, neutralized for 10 min in 0.1 M sodium borate, and incubated in a rat anti-BrdU primary antibody at 1:200 (Maine Biotechnology Services) overnight at 4˚C. The sections then were incubated with Cy3-conjugated anti-rat antibody (1:200 dilution [Jackson ImmunoResearch Laboratories, Inc.]) for 1 h at room temperature. For NG-2 and BrdU staining, NG-2 expression was resolved as described above with a FITC conjugated secondary antibody at 1:200, and then the sections were stained for cells that had incorporated BrdU as mentioned. Each of the steps was followed by three 5-min rinses in PBS. Tissue sections were mounted with Vectashield mounting medium (Vector Laboratories, Inc.).
After staining, the series of six slides at various levels from the same block were used for semiquantitative measurements. Quantitative measurements of immunostaining were performed by an observer blinded to the individual treatment status of the animals. Regions of interest (ROIs) were defined in the SVZ, at the dorsal end of the lateral ventricle, as a zone with 100 µm width and 800 µm length, and in the subcortical striatum with 200 µm width in the peri-hemorrhagic striatum for counting BrdU + cells, CNPase + cells, NG-2 + cells, CNPase + BrdU + cells, and NG-2 + BrdU + cells positive signals. Images were collected with a 40× objective using a CoolSNAP color camera (Photometrics) interfaced with a MCID system.
For quantitation of vascular endothelial cell (VEC) proliferation, 8 fields from the subcortical striatum were digitized under a 40× objective. The percentage of BrdU-positive VECs to a total of VECs found in 10 enlarged vessels located in the subcortical striatum was measured in each section using the MCID imaging analysis system.
2.5. HP Expression
Animals for HP expression were sacrificed at 7 days after ICH. At sacrifice, all animals were intracardially perfused with ice-cold PBS and the brain tissue immediately frozen in dry ice and stored in a −80 ˚C freezer prior to protein extraction. The Hp levels in the hematomal boundary zone and contralateral striatum were determined using a rat Hp ELISA kit (ICL, Newberg, OR) according to the manufacturer's instructions.
3. Results
3.1. Neurobehavioral Tests
To determine whether Tβ4 treatment regulates functional outcome after ICH in rats, we performed the corner turn and mNSS tests (Figure 1). All rats had normal scores for both neurobehavioral tests at baseline (Pre-ICH) and displayed similar neurological impairments at day 1 after ICH. No significant differences were found between treatment groups prior to 4 days post-ICH. Corner testing scores for all Tβ4 groups improved marginally at 1 week and significantly at 2, 3 and 4 weeks post-ICH compared to the saline-treated group. Significantly improved mNSS scores were found at weeks 1 to 4 in the Tβ4-treated groups compared with the saline-treated group. The 12 mg/kg Tβ4 group appeared to show mildly greater improvement compared with the 3 mg/kg Tβ4 group in the corner test at 4 weeks post-ICH, but these findings were not statistically significant. Improvement
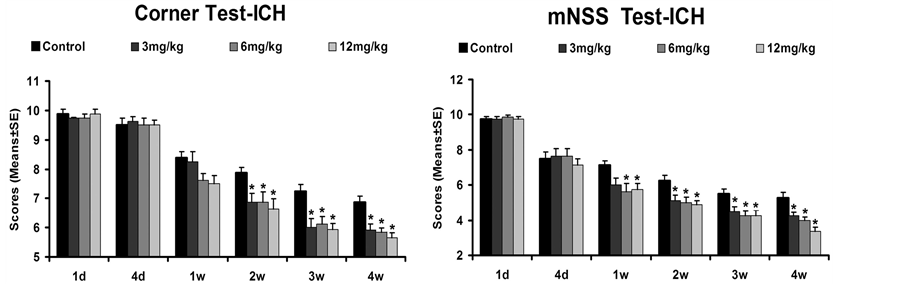
Figure 1. The effect of Tβ4 on functional recovery after ICH. Quantitative bar graph results of corner test (left panel) and modified neurological severity score (mNSS, right panel) are presented. Early Tβ4 treatment (starting at 24 h after ICH) significantly reduces corner turn at Days 14 - 28 and mNSS test scores at Days 7 - 28 compared with the saline-treated control group (p < 0.05). Significantly lower mNSS scores were shown in the 12 mg/kg Tβ4 group compared with the 3 mg/kg Tβ4 group (p < 0.05). Data represent mean ± SD.
of mNSS was significant at 4 weeks in the 12 mg/kg Tβ4 group when compared to the 3 mg/kg Tβ4 group.
3.2. Cell Proliferation
Staining with BrdU, an analog of thymidine, is commonly used to detect proliferating cells. Figure 2 demonstrates that a statistically significant effect on the number of BrdU+ cells was noted in the SVZ and the boundary zone around the hematoma at 4 weeks after ICH, compared with results in the control group. Marginally increased cell proliferation was noted in the 12 mg/kg Tβ4 group compared with the 3 mg/kg Tβ4 group. Angiogenesis may stimulate neurogenesis and enhances functional recovery after ICH. Enlarged and thin-walled vessels, containing BrdU immunoreactive positive endothelial cells are indicative of angiogenesis. Therefore, in the present study, BrdU-positive endothelial cells within a total of 8 enlarged and thin walled vessels located in the subcortical striatume were counted in each section. Figure 3 shows that BrdU-positive vascular endothelial cells (VECs) increased significantly in the Tβ4 treatment groups compared with control animals. However, there was no significant difference in VEC proliferation between the treatment groups. These findings suggest that Tβ4 promotes endothelial cell proliferation and angiogenesis.
3.3. Treatment with Tβ4 Increases OPCs and Mature Oligodendrocytes
NG-2 (a marker of OPCs) and CNPase (a marker of myelinating oligodendrocytes) were measured to address whether Tβ4 treatment affects OPCs and mature oligodendrocytes. Significant increases in actively proliferating OPCs were demonstrated within the peri-hematomal area and the SVZ at 4 weeks after ICH, when compared with the saline-treated group. Although no significant difference was detected among the 3 dose groups, the high-dose treatment (12 mg/kg) showed the greatest augmentation in the number of OPCs. Treatment with Tβ4 significantly increased the number of mature oligodendrocytes only in the peri-hematomal area region but not in the SVZ. Similarly, no significant difference was detected among the 3 dose groups in the number of CNPase- positive signals. To investigate whether the increasing OPCs and mature oligodendrocytes is correlated with functional outcome in rats, Pearson’s correlation coefficient was performed to analyze the relationship between the mNSS score and the number of NG2-positive and CNPase-positive signals at 4 weeks after ICH at the high-dose treatment group. Our data showed that the score of mNSS was significantly inversely correlated with them in the striatum adjacent to the hematoma (Figure 4).
3.4. Tβ4 Treatment Increases Oligodendrogenesis in the Brain of ICH Rats
Double immunostaining revealed that many NG2-positive cells in the peri-hematomal area and the SVZ were BrdU positive (Figure 5), indicating that these NG2-positive cells are within the cell cycle. Tβ4 enhanced this proliferation. Some CNPase-positive cells exhibited BrdU immuno-reactivity within the peri-hematomal area after Tβ4 treatment (Figure 6). Since the mature oligodendrocytes are post-mitotic and unable to proliferate
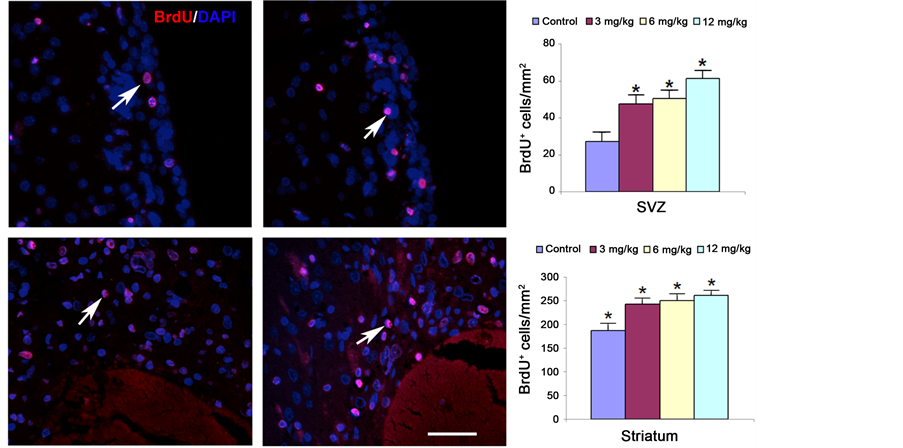
Figure 2. The effect of Tβ4 on BrdU-positive cell number in the ipsilateral SVZ (upper row) and peri-hematoma (lower row) after ICH. There are increased BrdU cells in the 3 mg/kg, 6 mg/kg, and 12 mg/kg (middle column) of Tβ4-treated rats compared to the saline control (left column) in both areas (right column). No significant difference is observed among the different dose treatment groups (right column).
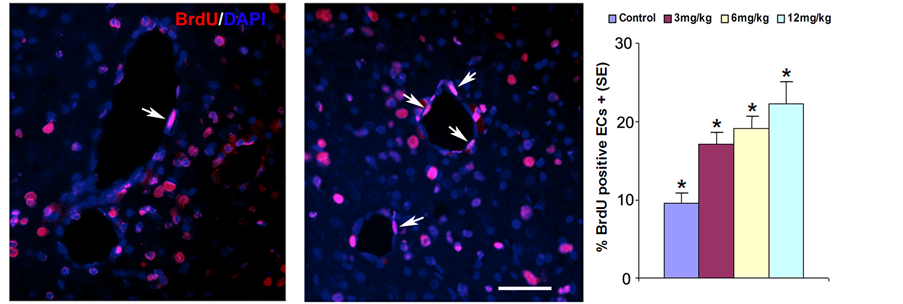
Figure 3. Vascular BrdU-positive endothelial cell (Ec) expression is increased in striatum (middle panel) of the Tβ4-treated rats when compared to saline control (left panel). Quantitative data (right panel) show significantly increased percentage of vascular endothelial cells compared to the saline control group.
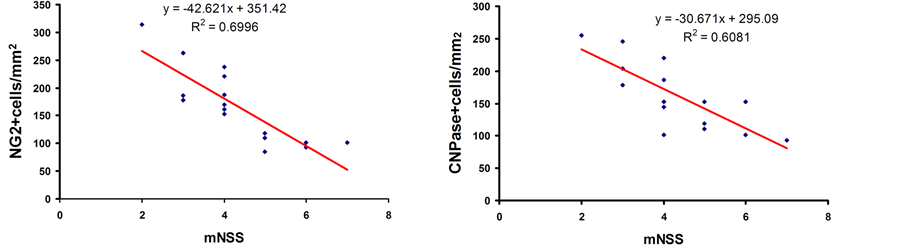
Figure 4. Correlation of functional recovery with proliferating OPCs and mature oligodendrocytes. The graph shows that the score of mNSS is significantly and inversely correlated with the number of NG2-positive and CNPase-positive signals at 4 weeks after ICH at the high-dose treatment group (p < 0.01).
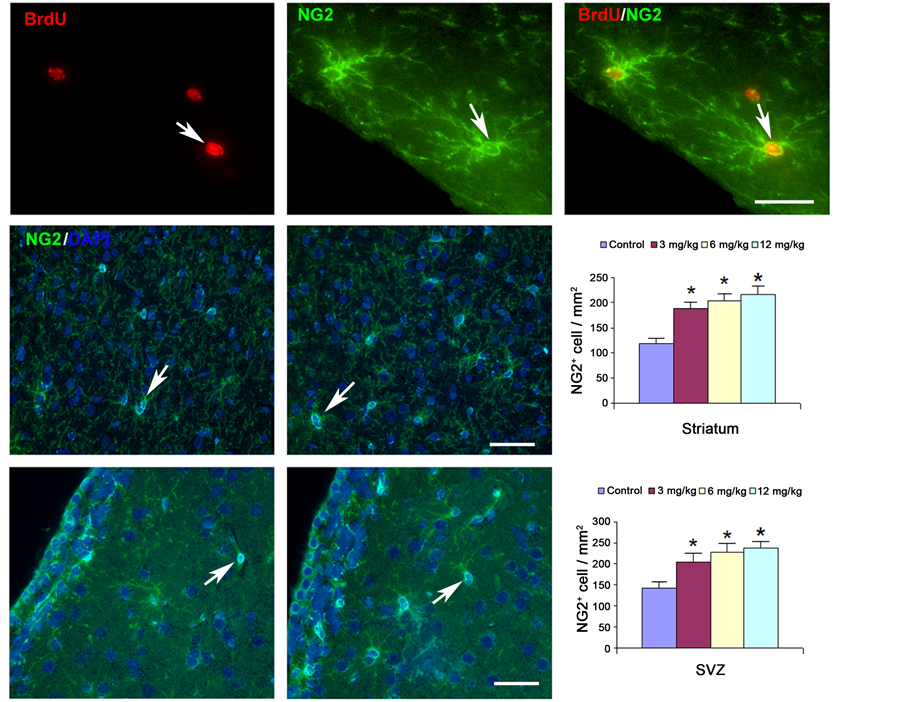
Figure 5. NG-2 staining (marker for proliferating OPCs) is increased in the striatum adjacent to the hematoma (middle center) and the ipsilateral SVZ (lower center) of the Tβ4-treated rats when compared to the saline control (middle left and lower left). Double staining (upper right) shows that BrdU-positive cells (upper left) are co-localized with NG-2 cells (upper center). Quantitative data (middle right and lower right) show significantly increased staining in the 3 mg/kg, 6 mg/kg, and 12 mg/kg of Tβ4-treated rats compared to the saline-treated control group. No significant difference is found between the different dose treatment groups.
[29] , these data suggest that the OPCs labeled with BrdU continue to divide before differentiating into myelinating oligodendrocytes after Tβ4 treatment.
3.5. Tβ4 Treatment Increases Haptoglobin (Hp) Expression in Brain Tissue
To analyze Tβ4-affected ICH rat brain for Hp protein, Hp protein levels in the striatum around the hematomal area were measured. The result showed a significant increase (0.044 ± 0.003 ng/mg, p < 0.05) at least 7 days after the onset of ICH, compared with saline-treated animals (0.031 ± 0.004 ng/mg). Similar to the increase of Hp in brain tissue, Tβ4 triggered a significant increase of Hp (2.23 ± 0.24 mg/dl, p < 0.05) in the peripheral blood as compared with the control group (1.38 ± 0.13 mg/dl). Functional tests showed improved neurological recovery at 7 days after ICH. These results suggest that increasing Hp in response to Tβ4 may help protect the ICH-af- fected brain. Our preliminary data suggest that administration of Tβ4 improves neurological functional outcome after ICH. The improved outcome may be attributed to increasing OPCs and over-expressing Hp against Hb (hemoglobin)-mediated toxicity [30] [31] .
4. Discussion
In the present study, Tβ4 significantly improved neurological outcome after experimental ICH. ICH induced oligodendrocytic progeny and Hp expression, and Tβ4 increased ICH-induced oligodendrocytic progeny and Hp
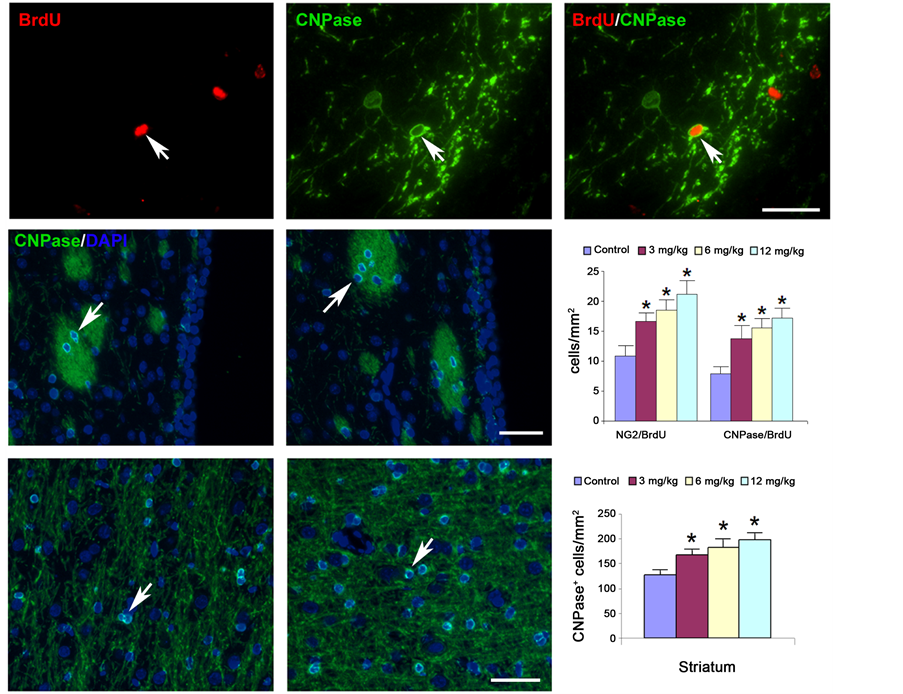
Figure 6. Myelinating oligodendrocytes measured by cyclic nucleotide phosphodiesterase (CNPase) staining (lower right) are increased in the striatum adjacent to the hematoma (lower center) of the Tβ4-treated rats compared to the saline control group (lower left). No CNPase-positive cells are found in SVZ (middle left and middle center). Double staining (upper right) shows that BrdU-positive cells (upper left) are co-localized with NG-2 cells (upper center). Quantitative data (middle right) show significantly increased CNPase staining and significantly increased NG-2/BrdU and CNPase/BrdU double staining in the 3 mg/kg, 6 mg/kg, and 12 mg/kg Tβ4-treated rats compared to the controls. No significant difference is found among the different dose treatment groups.
expression in young adult rats. These data suggest that amplification of OPC proliferation is a mechanism of Tβ4 treatment efficacy post-ICH in rats that coincides with increased Hp expression.
Tβ4 in doses up to 60 mg/kg shows no adverse reactions in rodent, dog and human models [32] . A toxicity study in Beagle dogs, however, observed salivation for a short time post administration at doses of 25 and 60 mg/kg [32] . In a randomized, double-blind, placebo-controlled, dose-response study, volunteers were given Tβ4 intravenously over a dose range of 42 - 1260 mg (18 mg/kg average 70 kg adult) and no dose-limiting toxicities or serious adverse events occurred. The most frequent adverse events are headache and upper respiratory infection. According to these findings and our previous studies in stroke and TBI [20] [26] , the mid-level doses ranged from 3 - 12 mg/kg. A biodistribution study of synthetic Tβ4 in normal mice provides evidence that intraperitoneal injection of 400 µg Tβ4 increases the concentrations of Tβ4 significantly in a variety of organs from the first 40 min to 2 h after injection compared to baseline concentrations. These organs include the brain (72 µg/g vs 42 µg/g), heart (80 µg/g vs 37 µg/g), liver (15 µg/g vs 9 µg/g), kidneys (65 µg/g vs 28 µg/g), and peritoneal fat (47 µg/g vs 13 µg/g) [33] . Our previous magnetic resonance study (MRI) study demonstrated that BBB permeability is increased after ICH [22] and the improved neurological function after Tβ4 treatment suggests that exogenous Tβ4 can cross the BBB.
Tβ4 is a G-actin binding protein that has been implicated in reorganizing actin cytoskeleton, which is needed for cell mobility [34] . Tβ4 has diverse biological functions based on its ability to affect cell behavior or in a paracrine role [34] . Tβ4 has been shown to promote healing in both wounds and corneal injury by increasing cell migration and reducing inflammation [11] [35] [36] . The wound-healing properties of Tβ4 were tested in a clinical trial [37] . In myocardial infarction animal model studies, Tβ4 was shown to promote the survival of cardiomyocytes and improve cardiac function [37] . Tβ4 has been found to be widely distributed in the mammalian central nervous system [15] [16] and it is expressed in most neural cell types of the developing brain and in subsets of neurons and microglia [17] . Exogenous Tβ4 has also been shown to promote the recovery of animals after ischemic stroke and brain trauma [18] . Our current data demonstrate that Tβ4 has the potential for treatment of ICH.
Tβ4 expression is up-regulated in many neurological diseases and its presence in the nervous system likely plays a role in neuroprotection, synaptogenesis, axon growth, cell migration, and changes in plasticity [18] [19] . Previous studies demonstrate that treatments promoting angiogenesis and neurogenesis significantly improve functional recovery after stroke and TBI. Tβ4 may have benefit after ICH by influencing different aspects of the injury/recovery process at different times: early after ICH, Tβ4 may promote endovascular protection, whereas later after 1 day, there may be induction of neurorestoration. ICH induces thrombosis in surrounding brain tissue, as does TBI. This thrombosis can play a role in the spread of cellular injury from the ICH/traumatic core. The influence of the Tβ4 on brain plasticity likely overlaps with reduction of thrombosis at the early time points, but beyond 1-3 days, the influence of the Tβ4 is probably no longer reduced thrombosis, but rather induced neurorestoration. In this study, we chose the therapeutic time at 24 hours post ICH to investigate the effects of Tβ4. Our results show that Tβ4 treatment promotes not only angiogenesis but also neurogenesis in the injured cortex and SVZ.
White matter injury, involving both axons and myelin-producing oligodendrocytes, contributes to mechanical injury, transport defects, ischemia and oxidative damage after ICH. Injury to white matter affects motor, sensory and cognitive function [38] . In our model the mNSS (a comprehensive motor and sensory test) and the cornering turn test (sensory and motor) results consistently indicate decreased neurological function after ICH. Therefore, an important therapeutic approach is to improve the neurological outcome after ICH by targeting repair of white matter damage in the parenchyma via stimulation of OPCs. A previous study in an ischemic stroke model showed that Tβ4 improves remyelination repair by increasing the OPCs and mature oligodendrocytes [26] . Similarly, our data showed that Tβ4 enhances the remyelination process when used to treat ICH.
Hp is an acute-phase a2-acid response plasma protein known for blocking Hb-induced oxidative toxicity. It binds with Hb and forms irreversible Hp-Hb complexes [39] . Hp is detected principally in the circulating blood and some peripheral organs and is present at very low levels in normal brain. After ICH, the expression profile of Hp in the brain is increased [40] [41] , suggesting that Hp may represent an endogenous factor normally involved in neutralizing the toxicity of hemolytic products in ICH-affected brain. Hp in brain is almost exclusively synthesized by oligodendrocytes as demonstrated in in vivo and in vitro models of ICH [42] . Oligodendrocytes themselves are highly vulnerable to oxidative stress [43] and could easily be damaged by hemolytic products. In this study our results show that Hp is increased in the brain tissue after Tβ4 treatment. This result combined with the increase of oligodendrocytes, suggests that Tβ4 treatment may promote oligodendrogenesis after ICH and the subsequent overexpression of Hb-neutralizing Hp.
5. Conclusion
In conclusion, we demonstrate that administration of Tβ4 24 h after ICH improves functional neurological outcome. Our data suggest that a remyelination repair process by the increase in OPCs and mature oligodendrocytes contributes to functional improvement. Our study is a proof-of-concept study, and further preclinical dose response studies are warranted to investigate Tβ4 and its potential role in the treatment of ICH.
References
- Morgenstern, L.B., Hemphill 3rd, J.C., Anderson, C., Becker, K., Broderick, J.P., Connolly Jr., E.S., Greenberg, S.M., Huang, J.N., MacDonald, R.L., Messe, S.R., Mitchell, P.H., Selim, M. and Tamargo, R.J. (2010) Guidelines for the Management of Spontaneous Intracerebral Hemorrhage: A Guideline for Healthcare Professionals from the American Heart Association/American Stroke Association. Stroke, 41, 2108-2129. http://dx.doi.org/10.1161/STR.0b013e3181ec611b
- Lee, S.H., Kim, B.J., Ryu, W.S., Kim, C.K., Kim, N., Park, B.J. and Yoon, B.W. (2010) White Matter Lesions and Poor Outcome after Intracerebral Hemorrhage: A Nationwide Cohort Study. Neurology, 74, 1502-1510. http://dx.doi.org/10.1212/WNL.0b013e3181dd425a
- Lok, J., Leung, W., Murphy, S., Butler, W., Noviski, N. and Lo, E.H. (2012) Intracranial Hemorrhage: Mechanisms of Secondary Brain Injury. Acta Neurochirurgica Supplementum, 111, 63-69. http://dx.doi.org/10.1007/978-3-7091-0693-8_11
- Masumura, M., Hata, R., Nagai, Y. and Sawada, T. (2001) Oligodendroglial Cell Death with DNA Fragmentation in the White Matter under Chronic Cerebral Hypoperfusion: Comparison between Normotensive and Spontaneously Hypertensive Rats. Neuroscience Research, 39, 401-412. http://dx.doi.org/10.1016/S0168-0102(01)00195-X
- Medana, I.M. and Esiri, M.M. (2003) Axonal Damage: A Key Predictor of Outcome in Human CNS Diseases. Brain, 126, 515-530. http://dx.doi.org/10.1093/brain/awg061
- Aguirre, A., Dupree, J.L., Mangin, J.M. and Gallo, V. (2007) A Functional Role for EGFR Signaling in Myelination and Remyelination. Neuroscience Research, 10, 990-1002. http://dx.doi.org/10.1038/nn1938
- Huff, T., Muller, C.S., Otto, A.M., Netzker, R. and Hannappel, E. (2001) Beta-Thymosins, Small Acidic Peptides with Multiple Functions. The International Journal of Biochemistry & Cell Biology, 33, 205-220. http://dx.doi.org/10.1016/S1357-2725(00)00087-X
- Safer, D., Sosnick, T.R. and Elzinga, M. (1997) Thymosin Beta 4 Binds Actin in an Extended Conformation and Contacts Both the Barbed and Pointed ends. Biochemistry, 36, 5806-5816. http://dx.doi.org/10.1021/bi970185v
- Sanders, M.C., Goldstein, A.L. and Wang, Y.L. (1992) Thymosin Beta 4 (Fx peptide) Is a Potent Regulator of Actin Polymerization in Living Cells. Proceedings of the National Academy of Sciences, 89, 4678-4682. http://dx.doi.org/10.1073/pnas.89.10.4678
- Amarenco, P., Bogousslavsky, J., Callahan 3rd, A., Goldstein, L.B., Hennerici, M., Rudolph, A.E., Sillesen, H., Simunovic, L., Szarek, M., Welch, K.M. and Zivin, J.A. (2006) High-Dose Atorvastatin after Stroke or Transient Ischemic Attack. New England Journal of Medicine, 355, 549-559. http://dx.doi.org/10.1056/NEJMoa061894
- Malinda, K.M., Sidhu, G.S., Mani, H., Banaudha, K., Maheshwari,
- Bock-Marquette, I., Saxena, A.,
- Santra, M., Chopp, M., Zhang,
- Smart, N., Risebro, C.A.,
- Carpintero, P., Anadon, R., Diaz-Regueira, S. and Gomez-Marquez, J. (1999) Expression of Thymosin Beta4 Messenger RNA in Normal and Kainate-Treated Rat Forebrain. Neuroscience, 90, 1433-1444. http://dx.doi.org/10.1016/S0306-4522(98)00494-1
- Gomez-Marquez, J. and Anadon, R. (2002) The Beta-Thymosins, Small Actin-Binding Peptides Widely Expressed in the Developing and Adult Cerebellum. The Cerebellum, 1, 95-102. http://dx.doi.org/10.1007/BF02941895
- Yang, H., Cheng, X., Yao, Q., Li, J. and Ju, G. (2008) The Promotive Effects of Thymosin Beta4 on Neuronal Survival and Neurite Outgrowth by Upregulating L1 Expression. Neurochemical Research, 33, 2269-2280. http://dx.doi.org/10.1007/s11064-008-9712-y
- MacLellan,
- Morris, D.C., Zhang,
- Xiong, Y., Zhang, Y., Mahmood, A., Meng, Y., Zhang,
- Zhang, J., Zhang,
- Knight,
- Seyfried, D., Ding, J.,
- Seyfried, D.,
- Xiong, Y., Mahmood, A., Meng, Y., Zhang, Y., Zhang,
- Morris, D.C., Chopp, M., Zhang, L., Lu, M. and Zhang, Z.G. (2010) Thymosin Beta4 Improves Functional Neurological Outcome in a Rat Model of Embolic Stroke. Neuroscience, 169, 674-682. http://dx.doi.org/10.1016/j.neuroscience.2010.05.017
- Zhang, L., Schallert, T., Zhang,
-
- Keirstead,
- Wu,
- Aronowski, J. and Zhao, X. (2011) Molecular Pathophysiology of Cerebral Hemorrhage: Secondary Brain Injury. Stroke, 42, 1781-1786. http://dx.doi.org/10.1161/STROKEAHA.110.596718
- Crockford, D. (2007) Development of Thymosin Beta4 for Treatment of Patients with Ischemic Heart Disease. Annals of the New York Academy of Sciences, 1112, 385-395. http://dx.doi.org/10.1196/annals.1415.051
- Mora, C.A., Baumann, C.A., Paino,
- Low,
- Sosne, G., Hafeez, S.,
- Sosne, G., Szliter,
- Guarnera, G.,
- Yang,
- Zhao, X., Song, S., Sun, G., Strong, R., Zhang, J., Grotta,
- Wagner,
- Wagner,
- Zhao, X., Song, S., Sun, G., Zhang, J., Strong, R., Zhang, L., Grotta,
- McTigue,
NOTES

*Corresponding author.