Open Journal of Ecology
Vol.2 No.4(2012), Article ID:24557,4 pages DOI:10.4236/oje.2012.24020
A chemical ecological investigation of the allelopathic potential of Lamium amplexicaule and Lamium purpureum
![]()
Department of Chemistry, University of Alabama in Huntsville, Huntsville, USA; *Corresponding Author: wsetzer@chemistry.uah.edu
Received 23 August 2012; revised 28 September 2012; accepted 10 October 2012
Keywords: Allelopathy; Essential Oil Composition; Lamium amplexicaule; Lamium purpureum
ABSTRACT
The overall goal of the project was to test the hypothesis that Lamium amplexicaule and Lamium purpureum, weedy invasive species to North America, use phytotoxic allelochemicals in interplant competition. The chemical compositions of the essential oils from the aerial parts of L. amplexicaule and L. purpureum have been obtained by hydrodistillation and analyzed by gas chromatography-mass spectrometry. The essential oils and several essential oil components have been screened for phytotoxic activity on lettuce (Lactuca sativa) and perennial ryegrass (Lolium perenne) as well as nematocidal activity against Caenorhabditis elegans, brine shrimp (Artemia salina) lethality, and insecticidal activity against the red imported fire ant (Solenopsis invicta × richteri). L. amplexicaule essential oil was composed largely of α-pinene, β- pinene, 1-octen-3-ol, (E)-caryophyllene, and germacrene D, while L. purpureum oil was dominated by α-pinene, β-pinene, 1-octen-3-ol, β-elemene, and germacrene D. Neither essential oil exhibited notable phytotoxicity or lethality against nematodes, brine shrimp, or fire ants. It is unlikely, therefore, that the allelopathy observed in these Lamium species is due to volatile phytochemical constituents.
1. INTRODUCTION
Both Lamium amplexicaule L. (henbit deadnettle) and Lamium purpureum L. (purple deadnettle) are non-native annual, weedy, invasive species, originally from Eurasia [1]. Invasion of exotic plant species can present a substantial problem to natural ecosystems as well as croplands and pastures [2,3]. Both L. amplexicaule and L. purpureum can aggressively spread through fields, pastures, and gardens. L. amplexicaule has been shown to reduce the yield of wheat (Triticum aestivum) [4], and L. purpureum has been shown to reduce the growth of soybeans (Glycine max) [5]. In this work, we test the hypothesis that volatile phytochemical components from these members of the mint family (Lamiaceae) are responsible for the reported allelopathic effects; we have determined the chemical compositions and biological activities of the essential oils from the aerial parts of L. amplexicaule and L. purpureum growing in fields in and near Huntsville, Alabama.
2. MATERIALS AND METHODS
2.1. Plant Material
For each species, plant materials were collected from different sites in and near Huntsville, Alabama. The aerial parts were harvested in the early morning, and the fresh plant materials (aerial parts) were chopped and hydrodistilled using a Likens-Nickerson apparatus to give clear, pale-yellow essential oils (Table 1). Plants were identified by Robert O. Lawton, Department of Biological Sciences, University of Alabama in Huntsville.
2.2. Gas Chromatography—Mass Spectrometry
The essential oils of the Lamium spp. were analyzed by GC-MS using an Agilent 6890 GC with Agilent 5973 mass selective detector [MSD, operated in the EI mode (electron energy = 70 eV), scan range = 40 - 400 amu, and scan rate = 3.99 scans/sec], and an Agilent ChemStation data system. The GC column was an HP-5ms fused silica capillary with a (5% phenyl)-polymethylsiloxane stationary phase, film thickness of 0.25 μm, a length of 30 m, and an internal diameter of 0.25 mm. The carrier gas was helium with a column head pressure of 48.7 kPa and a flow rate of 1.0 mL/min. Inlet temperature was
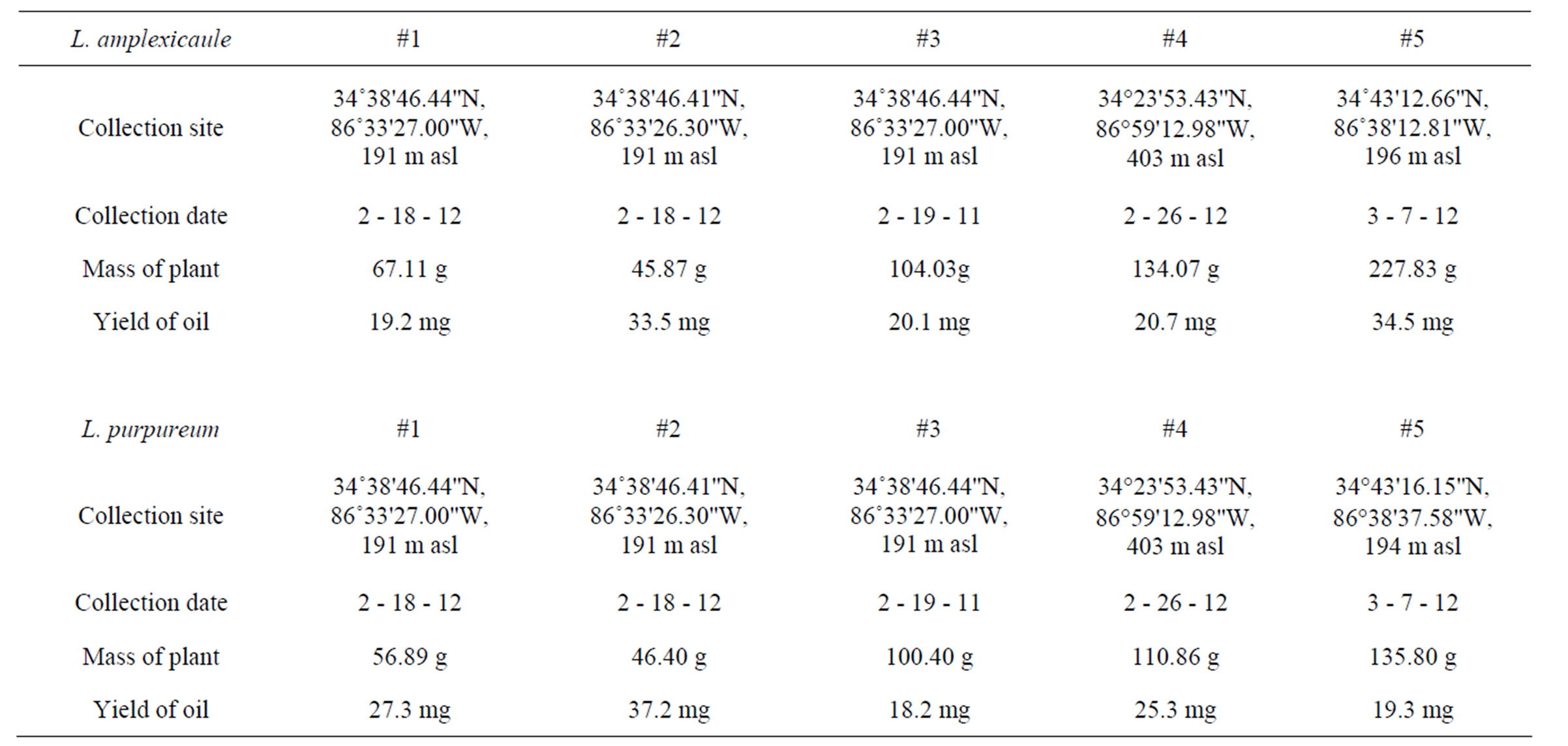
Table 1. Lamium amplexicaule and Lamium purpureum essential oil collection and yields.
200˚C and interface temperature was 280˚C. The GC oven temperature program was used as follows: 40˚C initial temperature, hold for 10 min; increased at 3˚C/min to 200˚C; increased 2˚/min to 220˚C. A 1% w/v solution of each sample in dichloromethane was prepared and 1 μL was injected using a 10:1 split ratio.
Identification of the oil components was based on their retention indices determined by reference to a homologous series of n-alkanes, and by comparison of their mass spectral fragmentation patterns with those reported in the literature [6] and stored on the MS library [NIST database (G1036A, revision D.01.00)/ChemStation data system (G1701CA, version C.00.01.080]. The percentages of each component are reported as raw percentages based on total ion current without standardization. The essential oil compositions of the L. amplexicaule and L. purpureum are summarized in Tables 2 and 3, respectively.
2.3. Allelopathic Activity Assays
An allelopathic bioassay based on lettuce (Lactuca sativa) and perennial rye grass (Lolium perenne) germination and subsequent radicle and hypocotyl growth [7] was measured to study the effects of the L. amplexicaule and L. purpureum essential oils and essential oil components. Stock solutions of each essential oil (4.0 g/L essential oil and 1.0 g/L Tween-80 in water) were prepared and used for the assays. Two-fold serial dilutions of stock test solutions were prepared to give test concentrations of 4000, 2000, 1000, 500, and 250 µg/mL with the control being 1.0 g/L aqueous Tween-80. Seeds were placed in Petri dishes (60 seeds per dish) each dish lined with two layers of Whatman No. 1 filter paper moistened with test solution and the Petri dishes were sealed with Parafilm®. The dishes were incubated at room temperature in the dark for 5 days, after which the number of germinated seeds was determined and the root (radicle) and shoot (hypocotyl) lengths were measured. The allelopathic assay results are summarized in Table 4.
2.4. Brine Shrimp Lethality Assay
The brine shrimp (Artemia salina) lethality test was carried out using a modification of the procedure by McLaughlin [8]. Artemia salina eggs were hatched in a sea salt solution (Instant Ocean®, 38 g/L) with an incandescent light bulb as the heat source. After 48 hours, the newly hatched nauplii were counted using a micropipette and transferred to 20-mL vials. Nine vials each containing 10 A. salina nauplii in 10 mL of sea salt solution (same as the hatching solution) were prepared. Three vials were labeled as controls with first one containing no DMSO, another with 10 µL, and the last one with 100 µL DMSO. Three replicate vials contained 10 µL of 1% essential oil solution in DMSO, and the other three were prepared by adding 100 µL of 1% essential oil solution in DMSO. Surviving A. salina were counted after 24 hours.
2.5. Nematocidal Assay
A nematocidal assay using Caenorhabditis elegans was carried out using a modification of the procedure of Park and co-workers [9]. Stock solutions of each essential oil (2.0 g/L essential oil and 1.0 g/L Tween-80 in water) were prepared and used for the assays. Dilutions
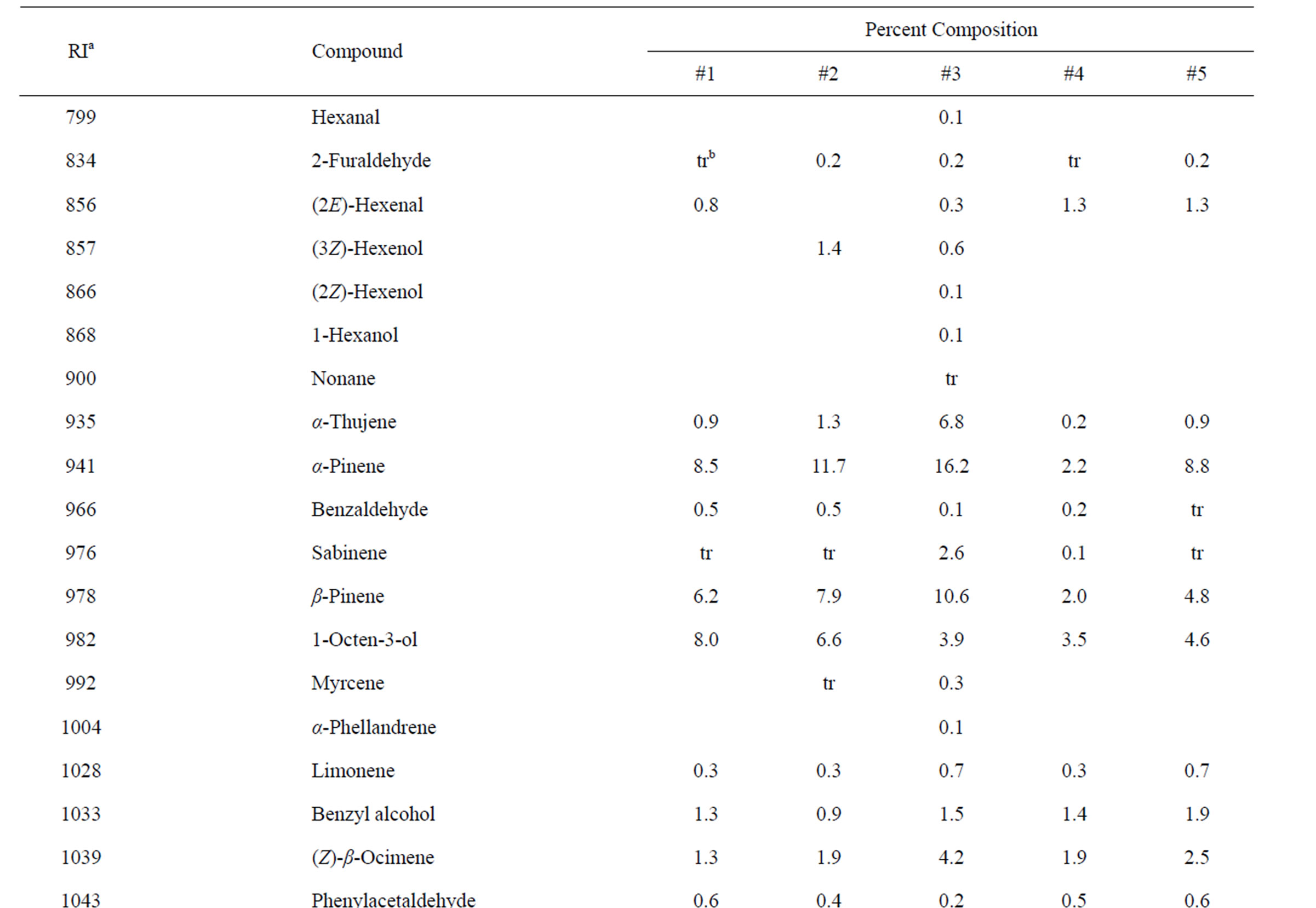
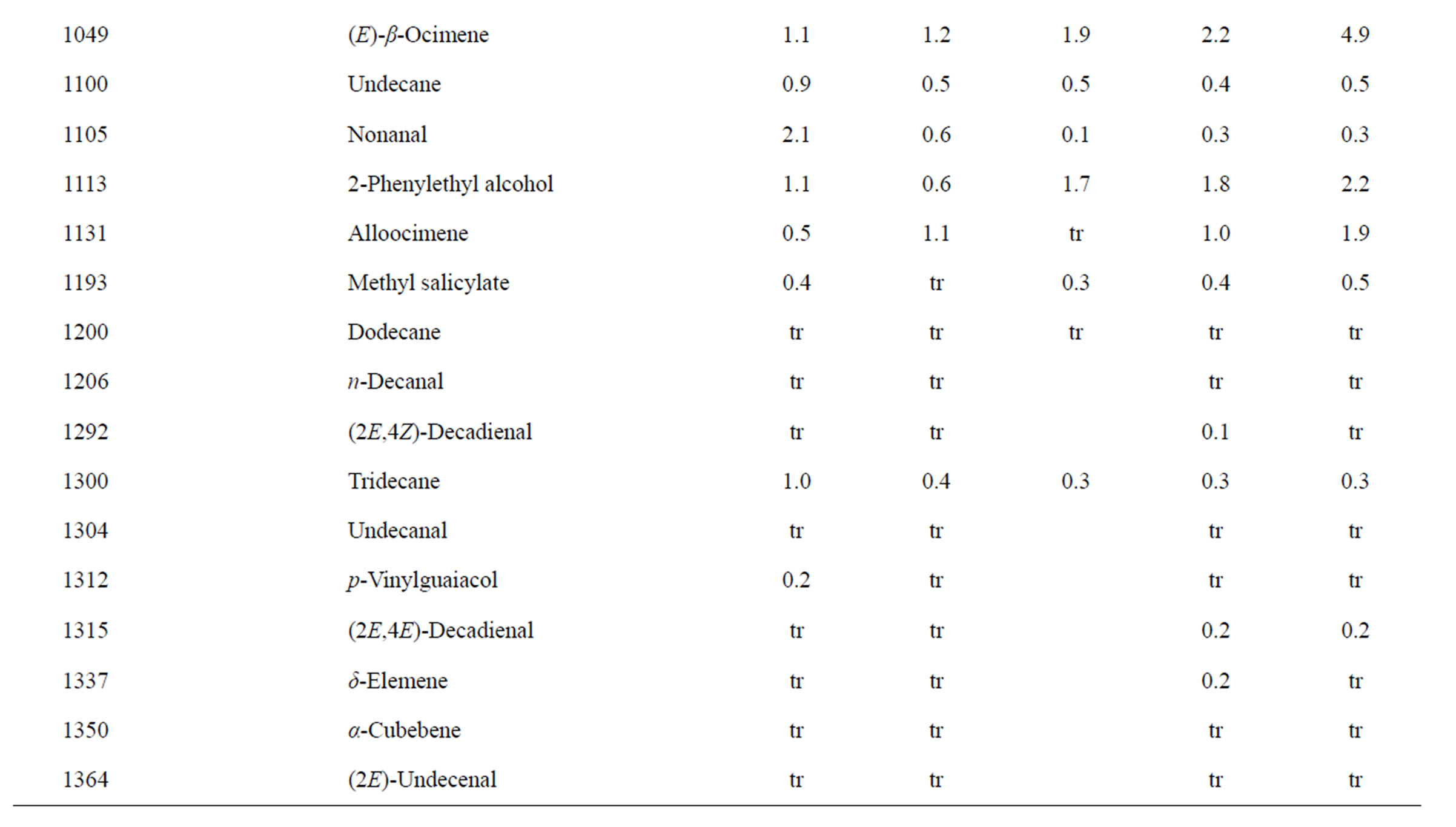
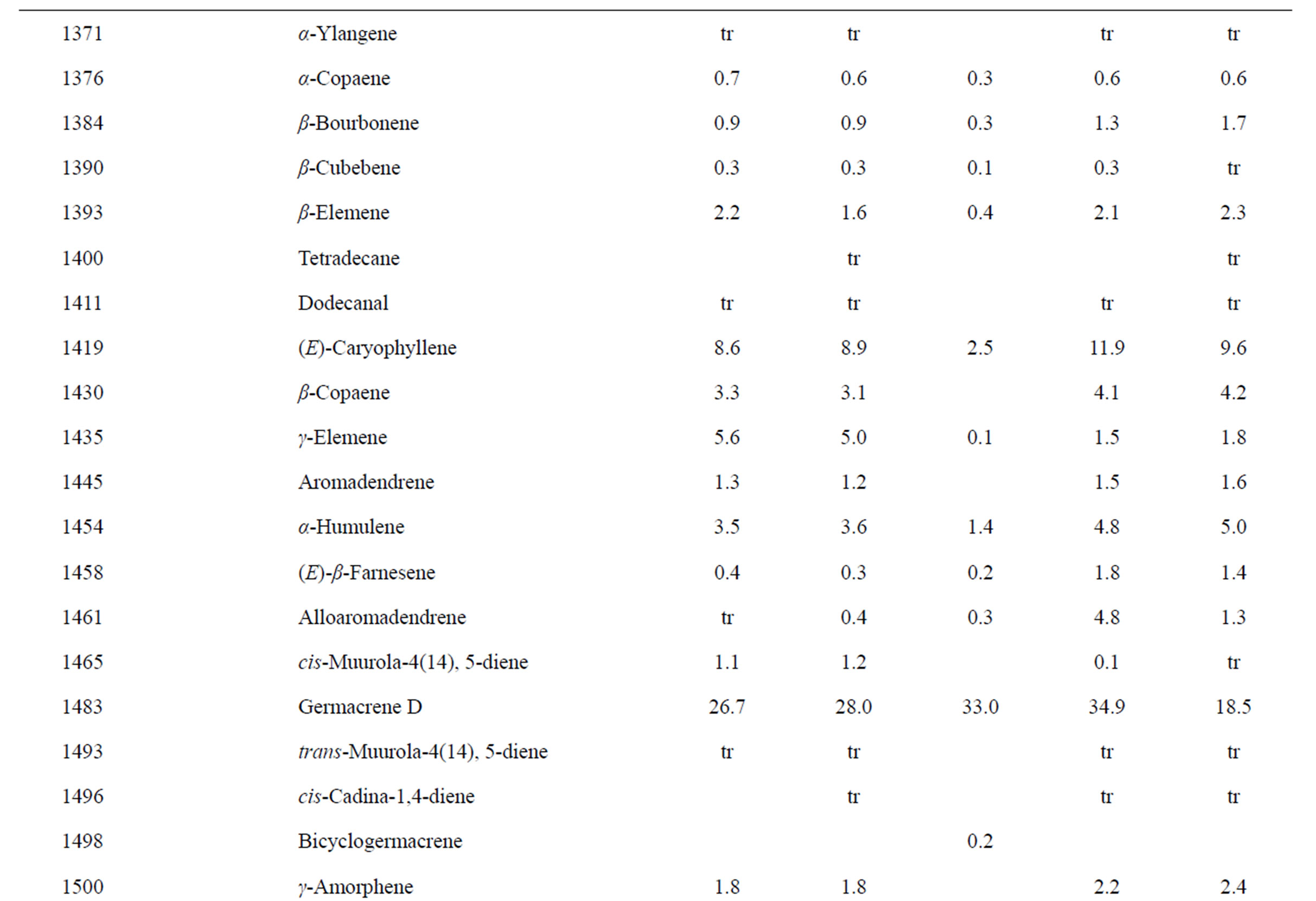
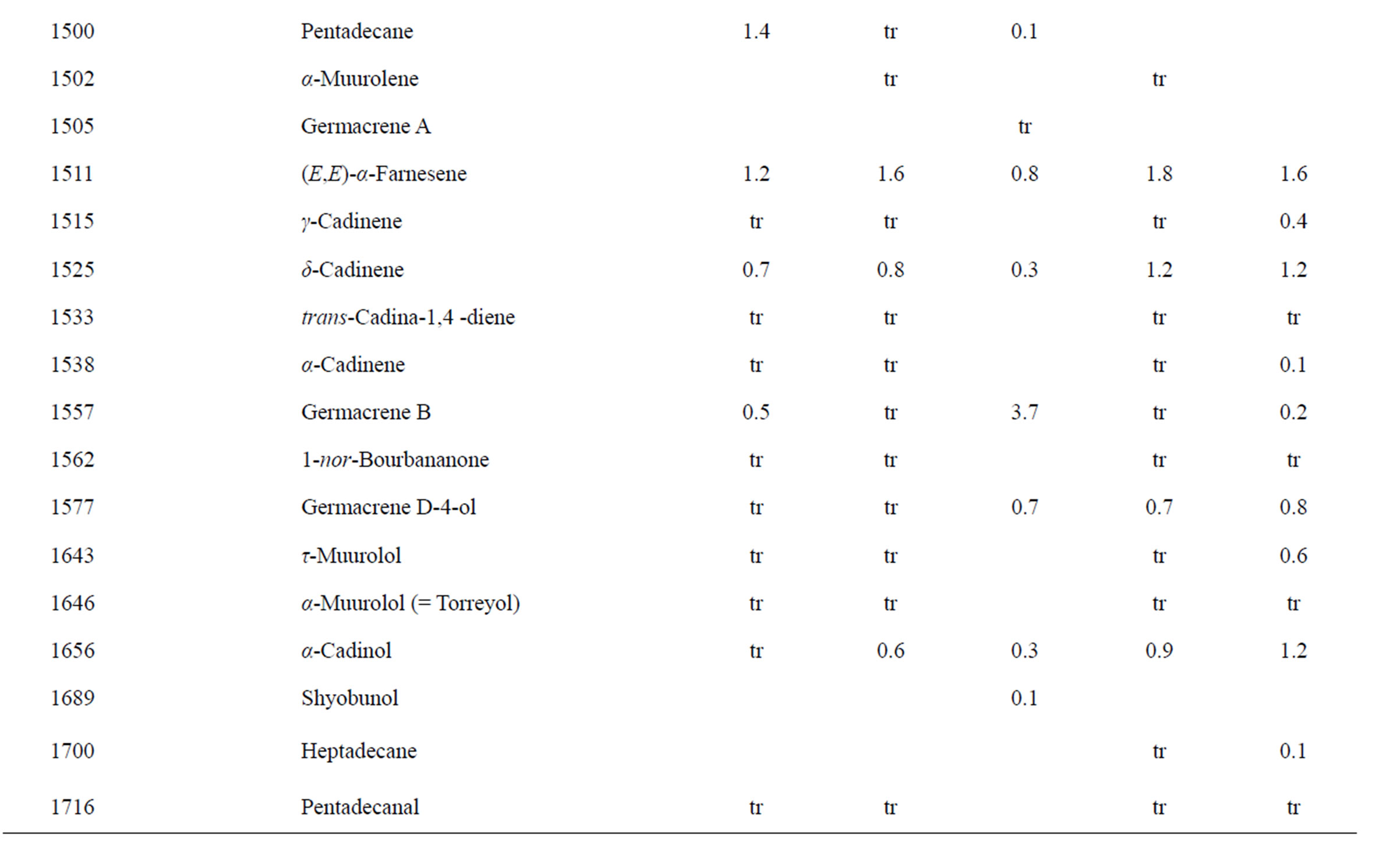

Table 2. Chemical compositions of Lamium amplexicaule essential oils.
of the sample solution were prepared in sterile water solution beginning with 50 μL of the 1% essential oil solution plus 50 μL sterile water. The sample solution was serially diluted (1:1) with sterile water in a 96-well plate. Into each well, 10 - 30 C. elegans (mixtures of juvenile and adult nematodes, male: female: juvenile ~1:1:2) per 50 μL of sample solution. Sterile water and serially diluted DMSO were used as controls. The dead and living nematodes were counted after 24 h under a microscope. Dead nematodes were identified by their immobility, and straight body, even after transfer to clean water.
2.6. Fire Ant Insecticidal Assay
Worker red imported fire ants, probably Solenopsis invicta × richteri hybrid [10], were collected from the University of Alabama in Huntsville. Sample solutions of 2000, 1000, 500, and 250 μg/mL were prepared in 1% aqueous Tween-80® solution. The control was 1% Tween solution. Each assay was carried out in triplicate using a 40-mL vial, fitted with a filter paper disk on the bottom. The filter paper was sprayed with 100 μL of sample solution, the walls of each vial were dusted with talcum powder to keep the ants from climbing out, and then 10 fire ant workers were transferred into each. The mortality of fire ants was recorded after 24 h. The bioassay was carried out in room temperature.
2.7. Statistical Analysis
Calculations were carried out using Excel. Student’s t-test [11] was used to compare radicle and hypocotyl test means with controls. Seed germination IC50 and A. salina LC50 values were determined using the Reed-Muench method [12].
3. RESULTS AND DISCUSSION
The essential oils of L. amplexicaule were dominated by sesquiterpene hydrocarbons, notably germacrene D (18.5% - 34.9%) and (E)-caryophyllene (2.5% - 11.9%),
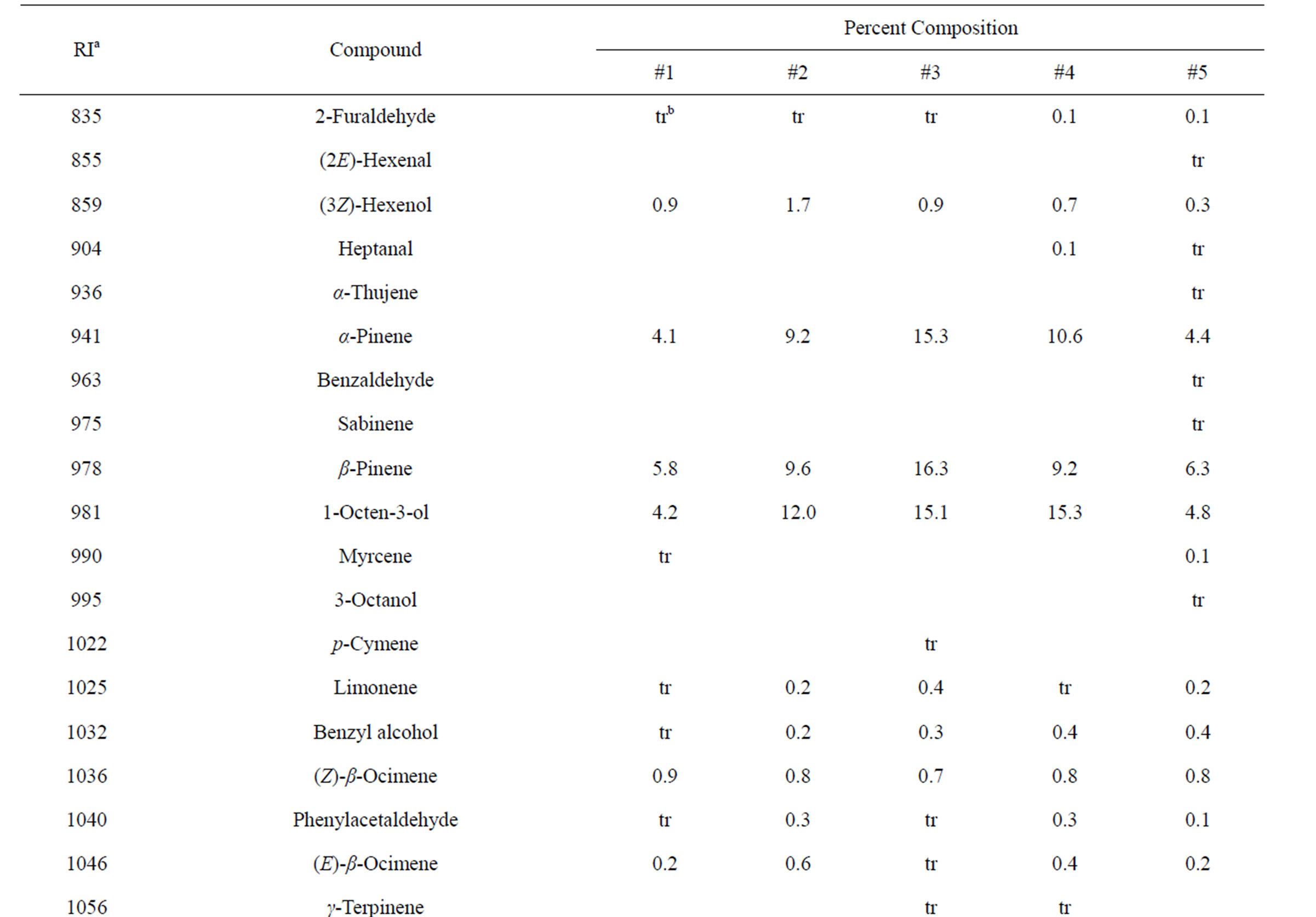
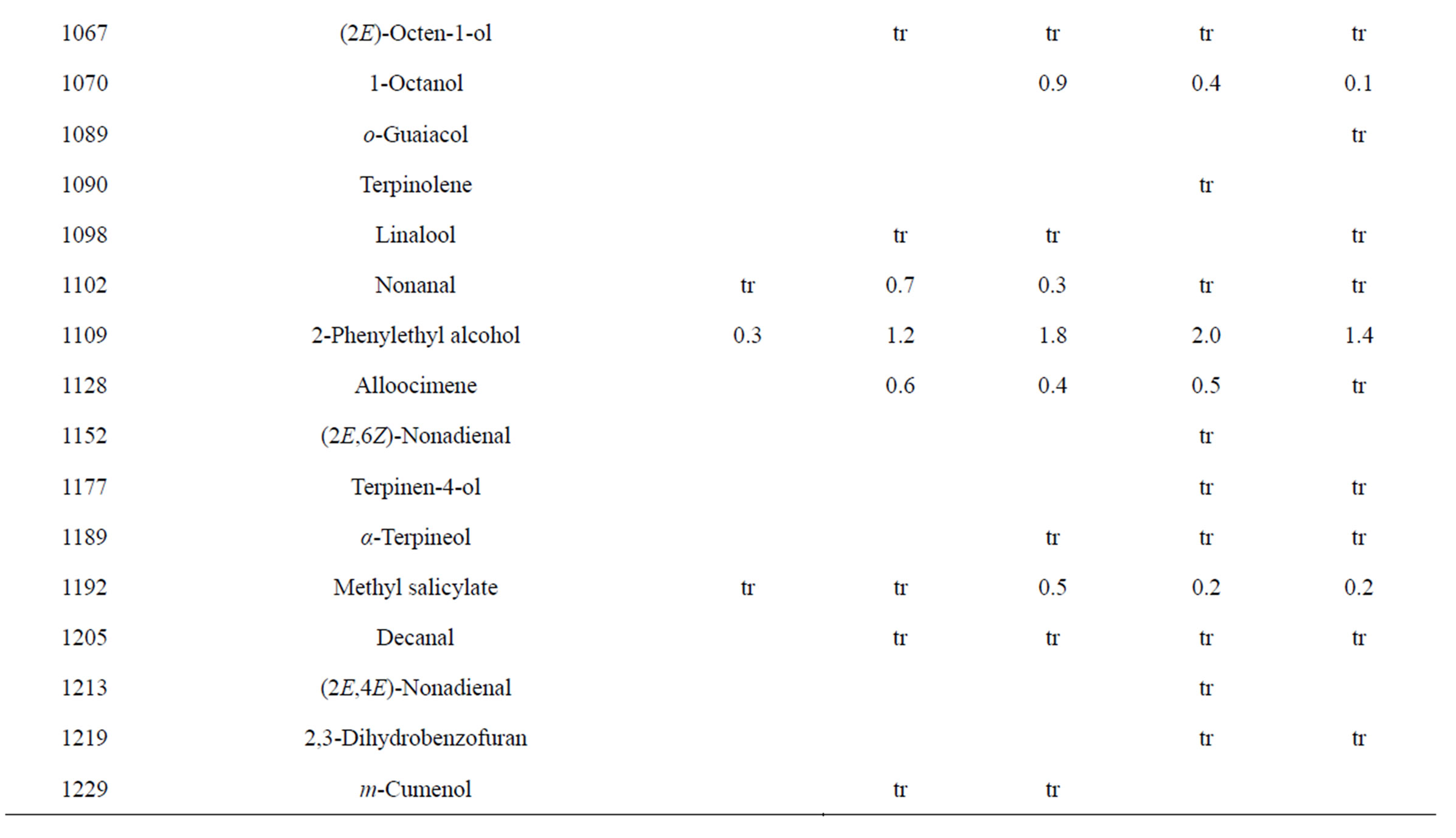
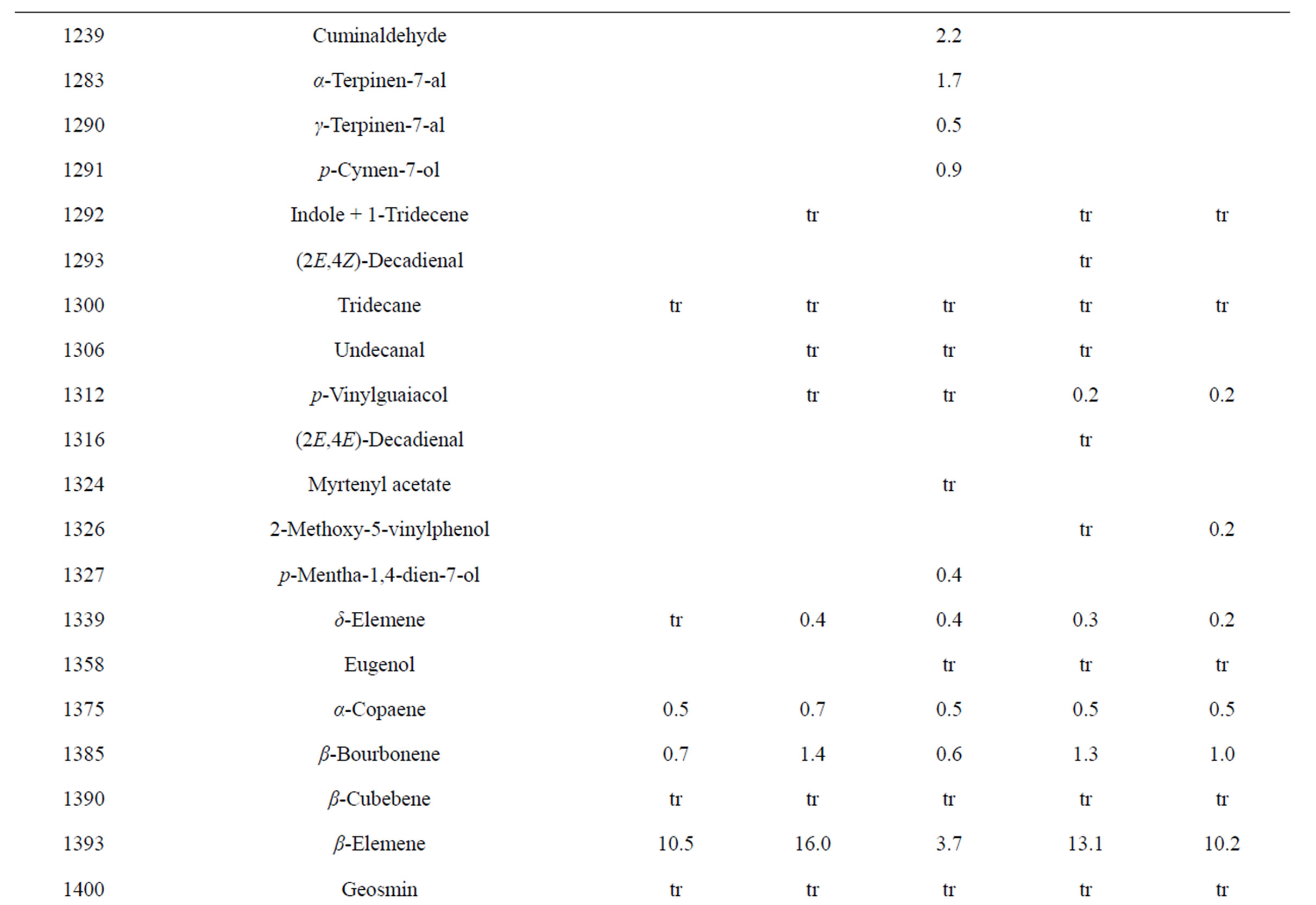
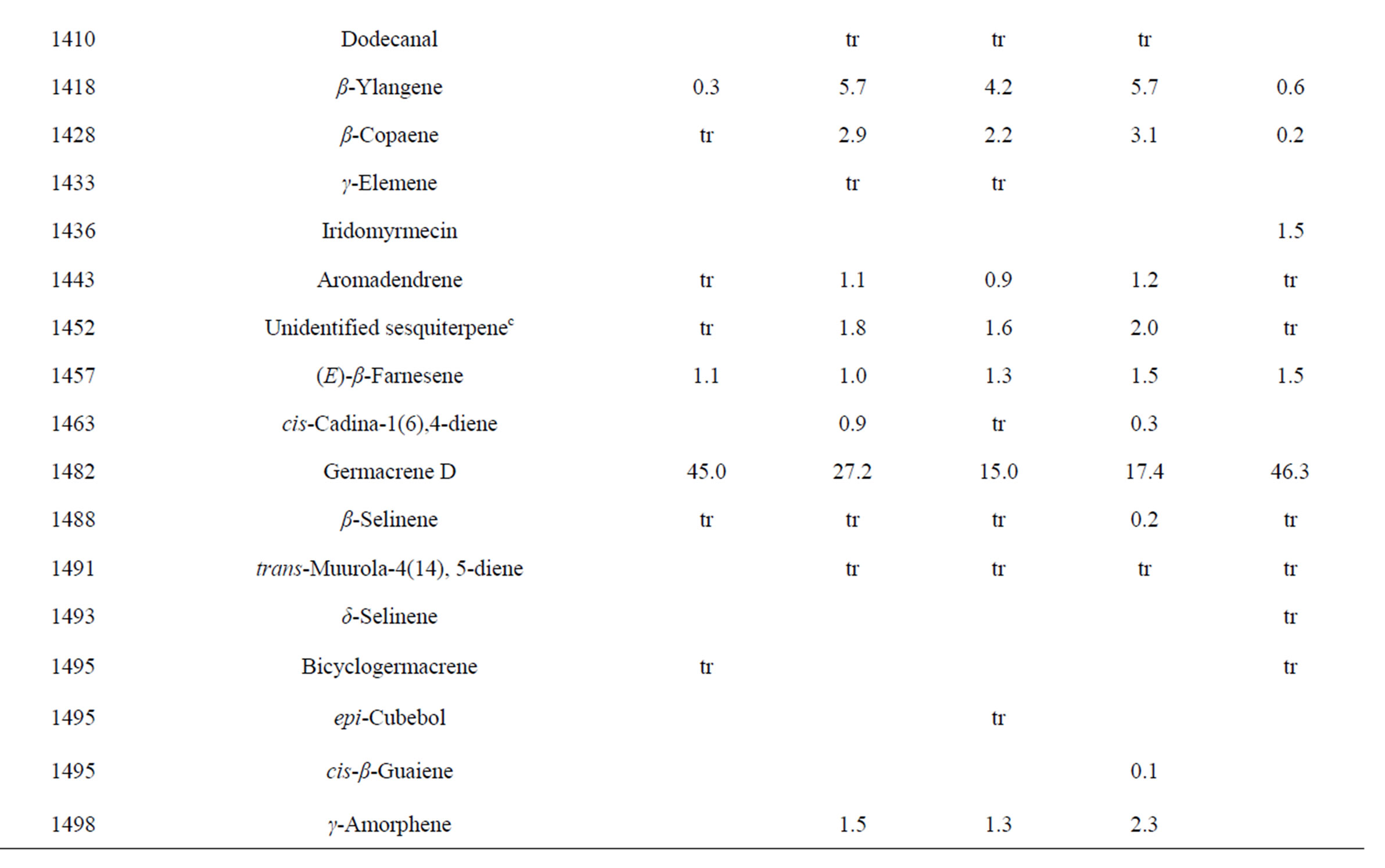
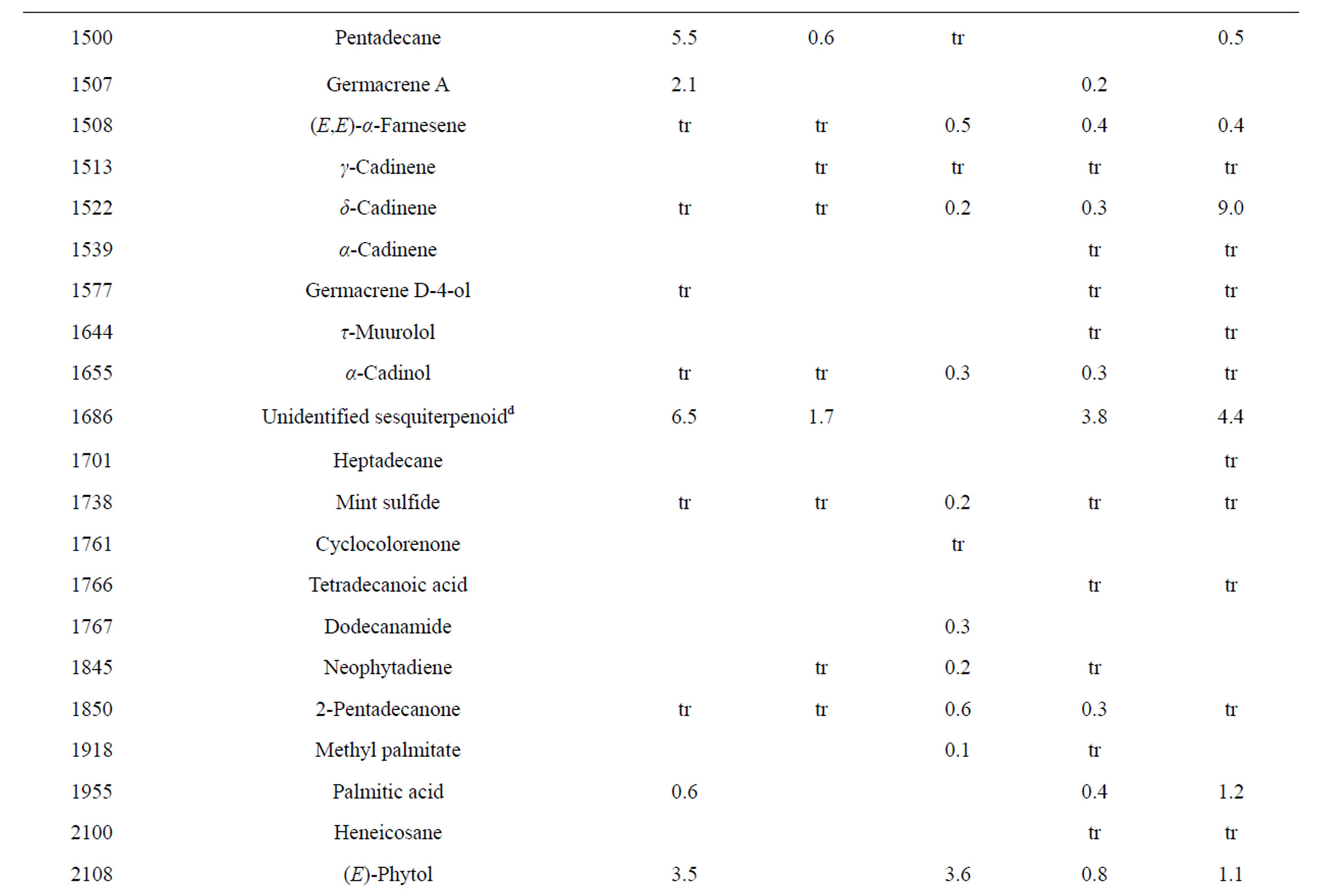
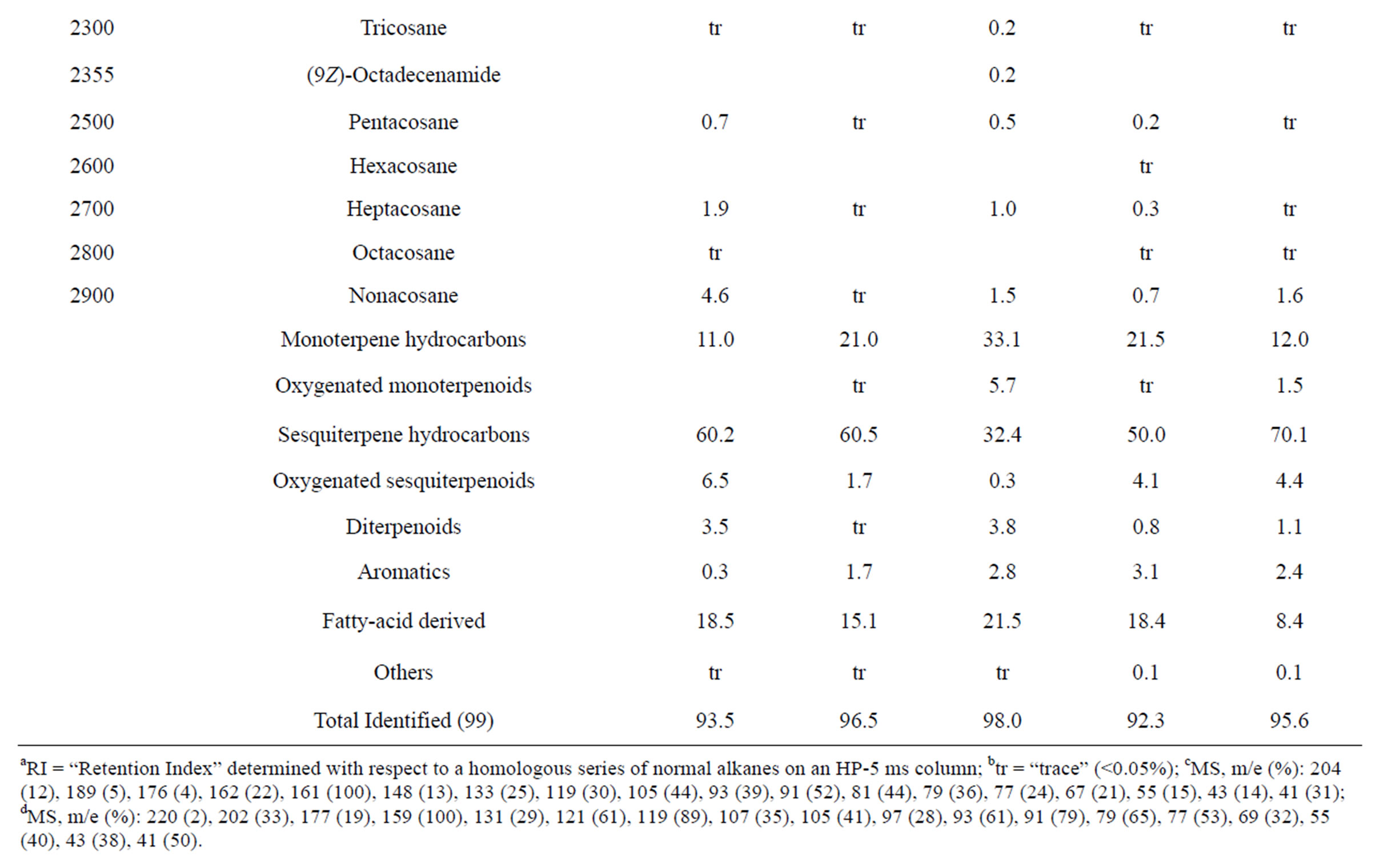
Table 3. Chemical compositions of Lamium purpureum essential oils.
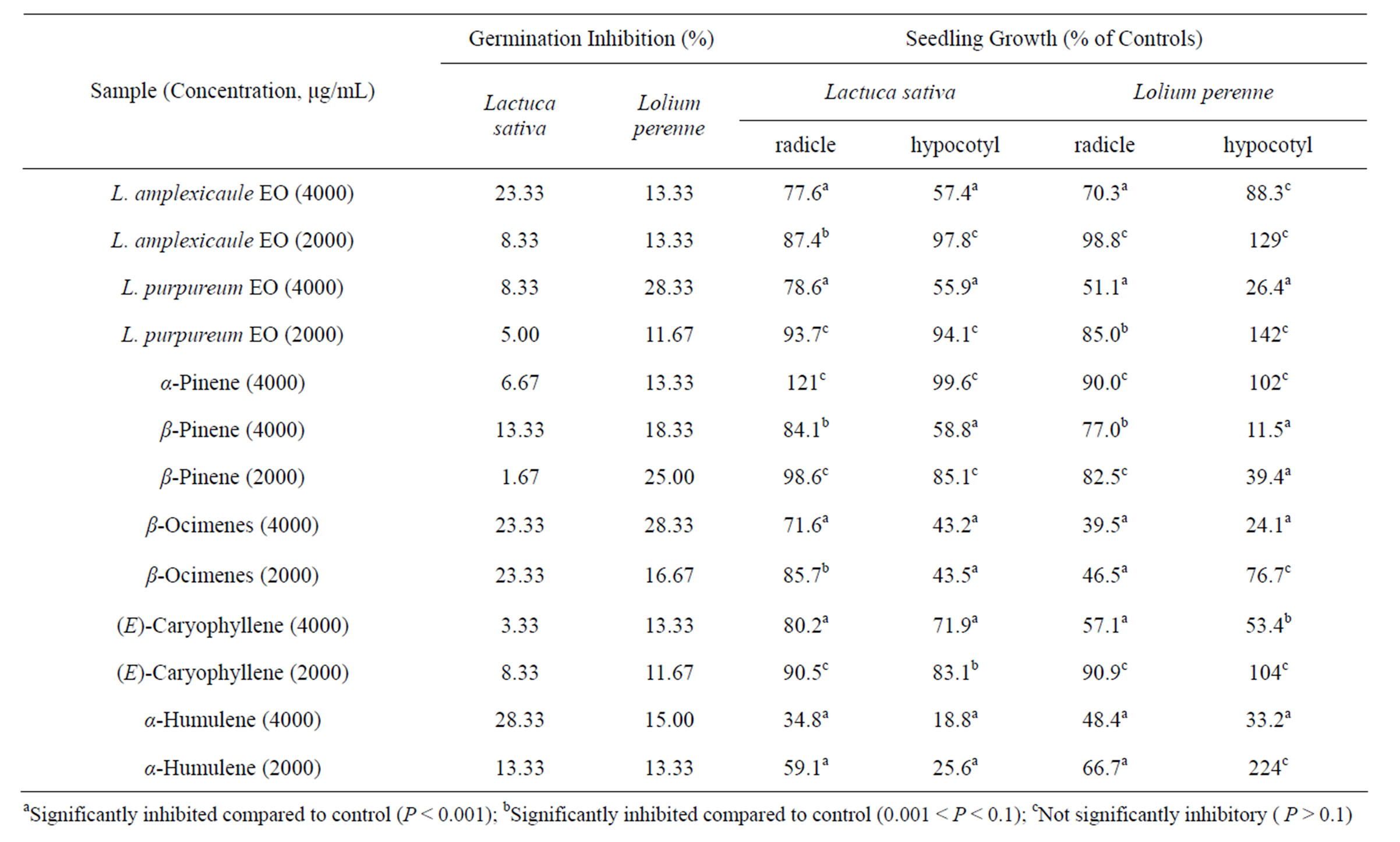
Table 4. Allelopathic activity of Lamium amplexicaule and Lamium purpureum essential oils on lettuce (Lactuca sativa) and perennial ryegrass (Lolium perenne).
and monoterpene hydrocarbons, α-pinene (2.2% - 16.2%), β-pinene (2.0% - 10.6%), as well as 1-octen-3-ol (3.5% - 8.0%). L. purpureum essential oil also had large concentrations of germacrene D (15.0% - 46.3%), α-pinene (4.1% - 15.3%), β-pinene (6.3% - 16.3%), and 1-octen-3-ol (4.2% - 15.3%). L. purpureum oil was also rich in β-elemene (3.7% - 16.0%), but devoid of (E)- caryophyllene. L. amplexicaule and L. purpureum essential oils from Italy have been reported [13], and while there are similarities, there are also some notable differences in compositions. The L. amplexicaule essential oil from Italy had no 1-octen-3-ol and no benzyl alcohol; trans-chrysanthenyl acetate was abundant (41.1%) in the Italian sample, but was not observed in the Alabama samples. Neither β-copaene nor γ-elemene was observed in the Italian L. amplexicaule sample. No 1-octen-3-ol was found in the Italian L. purpureum oil, but rather 1-octen-3-one, which was not detected in the Alabama samples. The Alabama samples did contain β-ylangene but not (E)-caryophyllene.
The essential oil of L. amplexicaule was only marginally allelopathic to lettuce (Lactuca sativa) or perennial ryegrass (Lolium perenne). At 4000 μg/mL, the essential oil showed only 23.3% and 13.3% germination inhibition of L. sativa and L. perenne, respectively. Radicle and hypocotyl elongation of these test species were only slightly inhibited at 4000 μg/mL, but largely unaffected at 2000 μg/mL.
Some of the essential oil components that were screened for allelopathic activity did demonstrate notable activity in terms of growth inhibition, but not germination inhibition. The monoterpenes α-pinene and β-pinene were inactive against L. sativa at 2000 μg/mL, but β-ocimene (mixture of isomers) inhibited growth of both L. sativa and L. perenne. The sesquiterpene (E)-caryophyllene was not notably active at 2000 μg/mL, but α-humulene did exhibit significant radicle elongation inhibition. Unfortunately, neither β-elemene nor germacrene D are commercially available and were not tested.
L. amplexicaule has been shown to reduce the yield of wheat (Triticum aestivum) [4], and has demonstrated allelopathic activity against lettuce (Lactuca sativa) [14]. Similarly, L. purpureum has been shown to reduce the growth of soybeans (Glycine max) [5], and this plant has also demonstrated allelopathic effects on lettuce [16]. Some plant species have demonstrated allelopathic effects toward L. amplexicaule and L. purpureum. For example, a field plot of rye (Secale cereale) reduced the biomass of weeds including L. amplexicaule [16]. Jerusalem artichoke (Helianthus tuberosus) residues reduced weed density, including L. purpureum [17].
L. amplexicaule and L. purpureum essential oils were screened for biological activity against the nematode Caenorhabditis elegans, the brine shrimp Artemia salinaand the red imported fire ant Solenopsis invicta × richteri. Neither oil showed activity against these organisms: C. elegans (LC50 > 2500 μg/mL), A. salina (LC50 > 100 μg/ mL), nor Solenopsis (LC50 > 4000 μg/mL). The absence of nematocidal activity on C. elegans is perhaps not surprising; both L. amplexicaule and L. purpureum are alternative and excellent hosts for the soybean cyst nematode, Heterodera glycines [18,19]. Likewise, insecticidal activity of L. amplexicaule or L. purpureum essential oils should not be expected as these plants are hosts to several generalist phytophagous insects [20]. Thus, for example, L. amplexicaule is an excellent host plant for the tarnished plant bug, Lygus lineolaris [21], the western flower thrip, Frankliniella occidentalis [22], and the silverleaf whitefly, Bemisia tabaci [23], while L. purpureum has been identified as a host for the black bean aphid, Aphis fabae [24]. These Lamium spp. are utilized by the insects as both feeding and reproductive hosts as well as overwintering hosts.
4. CONCLUSION
The allelopathic effects of Lamium amplexicaule and L. purpureum are not likely due to volatile phytochemical components; the essential oils of these plants do not exhibit notable phytotoxic effects. Previously observed allelopathy may have been due to non-volatile chemical components or due to biotic factors such as nematode infestations, phytophagous insect assaults, or plant viral diseases borne by infected plants and spread by insect vectors. L. amplexicaule is known to be a reservoir of tomato spotted wilt virus, which is spread by viruliferous Frankliniella thrips [25,26] as well as melon yellow spot virus, spread by Thrips palmi [27]. Similarly L. purpureum can be infected with cucumber mosaic virus [28] and potato virus Y [29], which are transmitted by the aphid Myzus persicae [30]. Additionally, the silverleaf whitefly, B. tabaci, is known to transmit several economically and ecologically important plant viruses [31].
5. ACKNOWLEDGEMENTS
CDJ and KEW are grateful for summer undergraduate research fellowships provided by the Provost’s Office of the University of Alabama in Huntsville. WNS is grateful to an anonymous private donor for the generous gift of the GC-MS instrumentation. We thank Prof. Robert O. Lawton for plant identification, Prof. Bernhard Vogler for technical assistance with GC-MS data collection, and Mr. Prabodh Satyal and Ms. Nidhi Goel for assistance with the biological assays.
REFERENCES
- Radford, A.E., Ahles, H.E. and Bell, C.R. (1968) Manual of the vascular flora of the carolinas. University of North Carolina Press, Chapel Hill.
- Pimentel, D., Lach, L., Zuniga, R. and Morrison, D. (2000) Environmental and economic costs of nonindigenous species in the United States. BioScience, 50, 53- 65. doi:10.1641/0006-3568(2000)050[0053:EAECON]2.3.CO;2
- Olliff, T., Renkin, R., McClure, C., Miller, P., Price, D., Reinhard, D. and Whipple, J. (2001) Managing a complex exotic vegetation program in Yellowstone National Park. Western North American Naturalist, 61, 347-358. https://ojs.lib.byu.edu/ojs/index.php/wnan/article/viewArticle/1012
- Conley, S.P. and Bradley, K.W. (2005) Wheat (Triticum aestivum) yield response to henbit (Lamium amplexicaule) interference and simulated winterkill. Weed Technology, 19, 902-906. doi:10.1614/WT-04-252R.1
- Mock, V.A., Creech, E., Davis, V.M. and Johnson, W.G. (2009) Plant growth and soybean cyst nematode response to purple deadnettle (Lamium purpureum), annual ryegrass, and soybean combinations. Weed Science, 57, 489- 493. doi:10.1614/WS-09-005.1
- Adams, R.P. (2007) Identification of essential oil components by gas chromatography/mass spectrometry. 4th Edition, Allured Publishing Corp., Carol Stream.
- Kennedy, J.E., Davé, P.C., Harbin, L.N. and Setzer, W.N. (2011) Allelopathic potential of Sassafras albidum and Pinus taeda essential oils. Allelopathy Journal, 27, 111- 122. http://indianjournals.com/ijor.aspx?target=ijor:aj&volume=27&issue=1&article=011
- McLaughlin, J.L. (1991) Bench-top bioassays for the discovery of bioactive compounds in higher plants. Brenesia, 34, 1-14.
- Park, I.K., Kim, J., Lee, S.G. and Shin, S.C. (2007) Nematicidal activity of plant essential oils and components from ajowan (Trachyspermum ammi), allspice (Pimenta dioica) and litsea (Litsea cubeba) essential oils against pine wood nematode (Bursaphelenchus xylophilus). Journal of Nematology, 39, 257-279. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2586506/
- Chen, L., Lu, Y.Y., Hu, Q.B. and Fadamiro, H.Y. (2012) Similarity in venom alkaloid chemistry of alate queens of imported fire ants: Implication for hybridization between Solenopsis richteri and S. invicta in the southern United States. Chemistry & Biodiversity, 9, 702-713. doi:10.1002/cbdv.201100109
- Zar, J.H. (1996) Biostatistical analysis. 3rd Edition, Prentice Hall, New Jersey.
- Reed, L.J. and Muench, H. (1938) A simple method of estimating fifty percent endpoints. American Journal of Hygiene, 27, 493-497.
- Glamini, G., Cioni, P.L. and Morelli, I. (2005) Composition of the essential oils and in vivo emission of volatiles of four Lamium species from Italy: L. purpureum, L. hybridum, L. bifidum, and L. amplexicaule. Food Chemistry, 91, 63-68. doi:10.1016/j.foodchem.2004.05.047
- Fujii, Y., Shibuya, T. and Yasuda, T. (1992) Allelopathy of velvetbean: Its discrimination and identification of LDOPA as a candidate of allelopathic substances. Japan Agricultural Research Quarterly, 25, 238-247.
- Shiraishi, S., Watanabe, I., Kuno, K. and Fujii, Y. (2002) Allelopathic activity of leaching from dry leaves and exudate from roots of ground cover plants assayed on agar. Weed Biology and Management, 2, 133-142. doi:10.1046/j.1445-6664.2002.00063.x
- Pérez, F.J. and Ormeño-Núñez, J. (1993) Weed growth interference from temperate cereals: The effect of hydroxamic-acids-exuding rye (Secale cereale L.) cultivar. Weed Research, 33, 115-119. doi:10.1111/j.1365-3180.1993.tb01924.x
- Tesio, F., Vidotto, F. and Ferrero, A. (2012) Allelopathic persistence of Helianthus tuberosus L. residues in the soil. Scientia Horticulturae, 125, 98-105. doi:10.1016/j.scienta.2011.12.008
- Creech, J.E., Faghihi, J., Ferris, V.R., Westphal, A. and Johnson, W.G. (2007) Influence of intraspecific henbit (Lamium amplexicaule) and purple deadnettle (Lamium purpureum) competition on soybean cyst nematode reproduction. Weed Science, 55, 665-670. doi:10.1614/WS-07-051.1
- Creech, J.E., Webb, J.S., Young, B.G., Bond, J.P., Harrison, S.K., Ferris, V.R., Faghihi, J., Westphal, A. and Johnson, W.G. (2007) Development of soybean cyst nematode on henbit (Lamium amplexicaule) and purple deadnettle (Lamium purpureum). Weed Technology, 21, 1064-1070. doi:10.1614/WT-07-079.1
- Marshall, E.J.P., Brown, V.K., Boatman, N.D., Lutman, P.J.W., Squire, G.R. and Ward, L.K. (2003) The role of weeds in supporting biological diversity within crop fields. Weed Research, 43, 77-89. doi:10.1046/j.1365-3180.2003.00326.x
- Esquivel, J.F. and Mowery, S.V. (2007) Host plants of the tarnished plant bug (Heteroptera: Miridae) in central Texas. Environmental Entomology, 36, 725-730. doi:10.1603/0046-225X(2007)36[725:HPOTTP]2.0.CO;2
- Atakan, E. (2010) Influence of weedy field margins on abundance patterns of the predatory bugs Orius spp. and their prey, the western flower thrips (Frankliniella occidentalis), on faba bean. Phytoparasitica, 38, 313-325. doi:10.1007/s12600-010-0105-9
- Bayhan, E., Ulusoy, M.R. and Brown, J.K. (2006) Host range, distribution, and natural enemies of Bemisia tabaci “B biotype” (Hemiptera: Aleyrodidae) in Turkey. Journal of Pesticide Science, 79, 233-240. doi:10.1007/s10340-006-0139-4
- Wilkinson, T.L. and Douglas, A.E. (2003) Phloem amino acids and the host plant range of the polyphagous aphid, aphis fabae. Entomologia Experimentalis et Applicata, 106, 103-113. doi:10.1046/j.1570-7458.2003.00014.x
- Groves, R.L., Walgenbach, J.F., Moyer, J.W. and Kennedy, G.G. (2002) The role of weed hosts and tobacco thrips, Frankliniella fusca, in the epidemiology of tomato spotted wilt virus. Plant Disease, 86, 573-582. doi:10.1094/PDIS.2002.86.6.573
- Okazaki, S., Okuda, M., Komi, K., Yoshimatsu, H. and Iwanami, T. (2007) Overwintering viruliferous Frankliniella occidentalis (Thysanoptera: Thripidae) as an infection source of tomato spotted wilt virus in green pepper fields. Plant Disease, 91, 842-846. doi:10.1094/PDIS-91-7-0842
- Yamasaki, S., Okazaki, S. and Okuda, M. (2012) Temporal and spatial dispersal of melon yellow spot virus in cucumber greenhouses and evaluation of weeds as infection sources. European Journal of Plant Pathology, 132, 169-177. doi:10.1007/s10658-011-9860-9
- Tomlinson, J.A., Carter, A.L., Dale, W.T. and Simpson, C.J. (1970) Weed plants as sources of cucumber mosaic virus. Annals of Applied Biology, 66, 11-16. doi:10.1111/j.1744-7348.1970.tb04597.x
- Kaliciak, A. and Syller, J. (2009) Aphid transmissibility of genetically different isolates of potato virus Y and susceptibility of weeds to virus infection. Biuletyn Instytutu Hodowli i Aklimatyzacji Roślin, 253, 285-295. http://www.cabdirect.org/abstracts/20103094750.html
- Tomlinson, J.A. and Carter, A.L. (1970) Studies on the seed transmission of cucumber mosaic virus in chickweed (Stellaria media) in relation to the ecology of the virus. Annals of Applied Biology, 66, 381-386. doi:10.1111/j.1744-7348.1970.tb04617.x
- Oliveira, M.R.V., Henneberry, T.J. and Anderson, P. (2001) History, current status, and collaborative research projects for Bemisia tabaci. Crop Protection, 20, 709-723. doi:10.1016/S0261-2194(01)00108-9