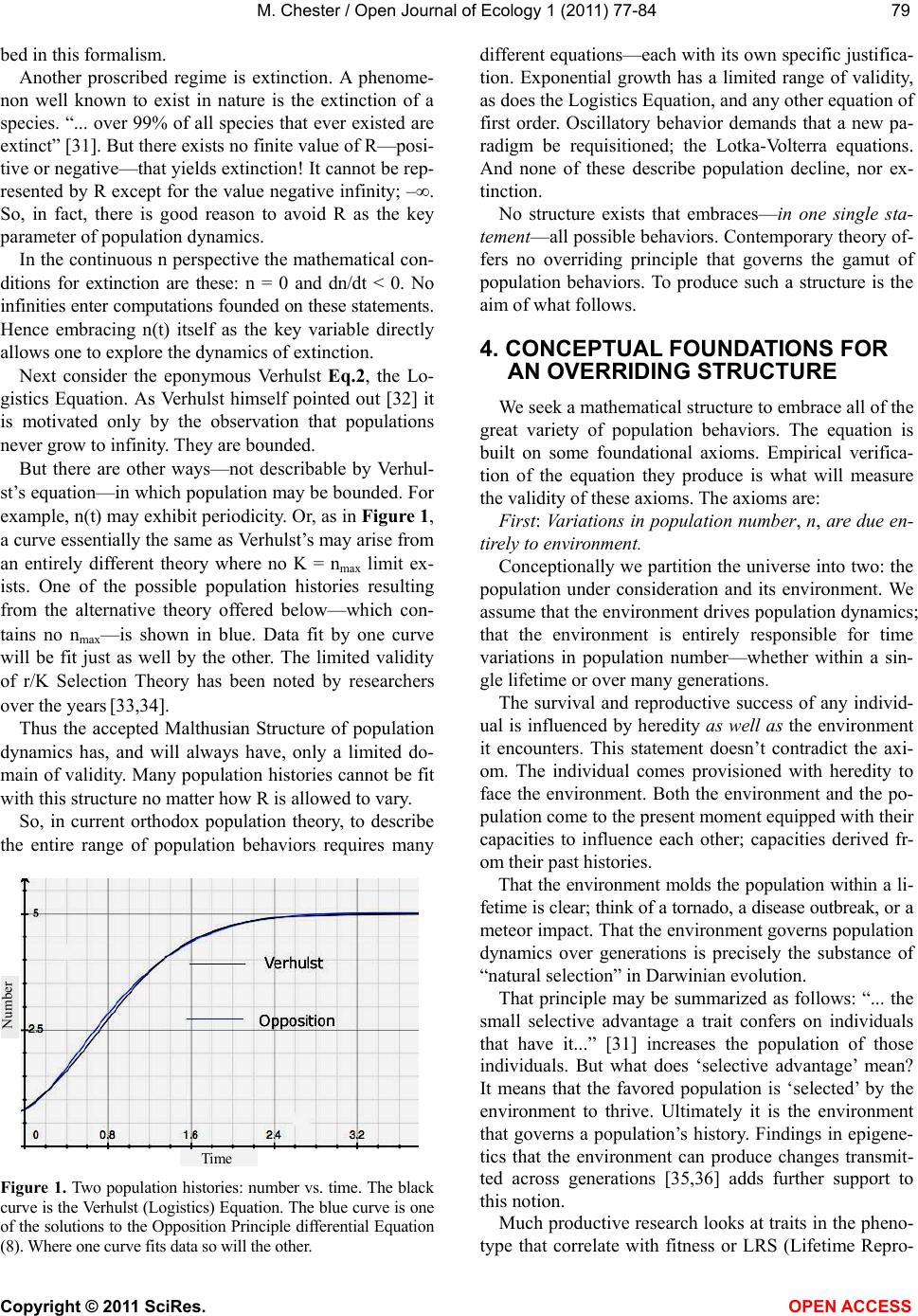
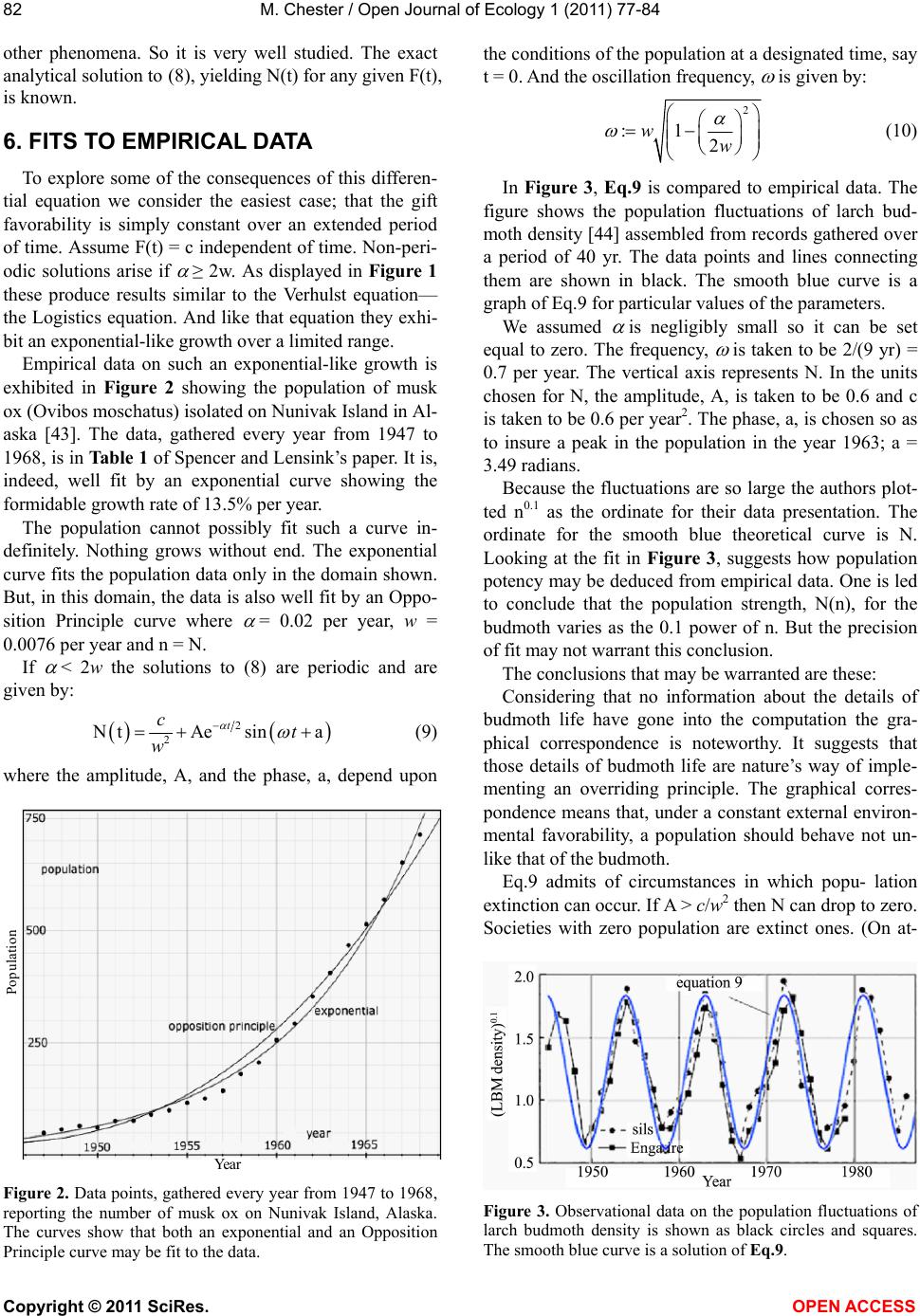
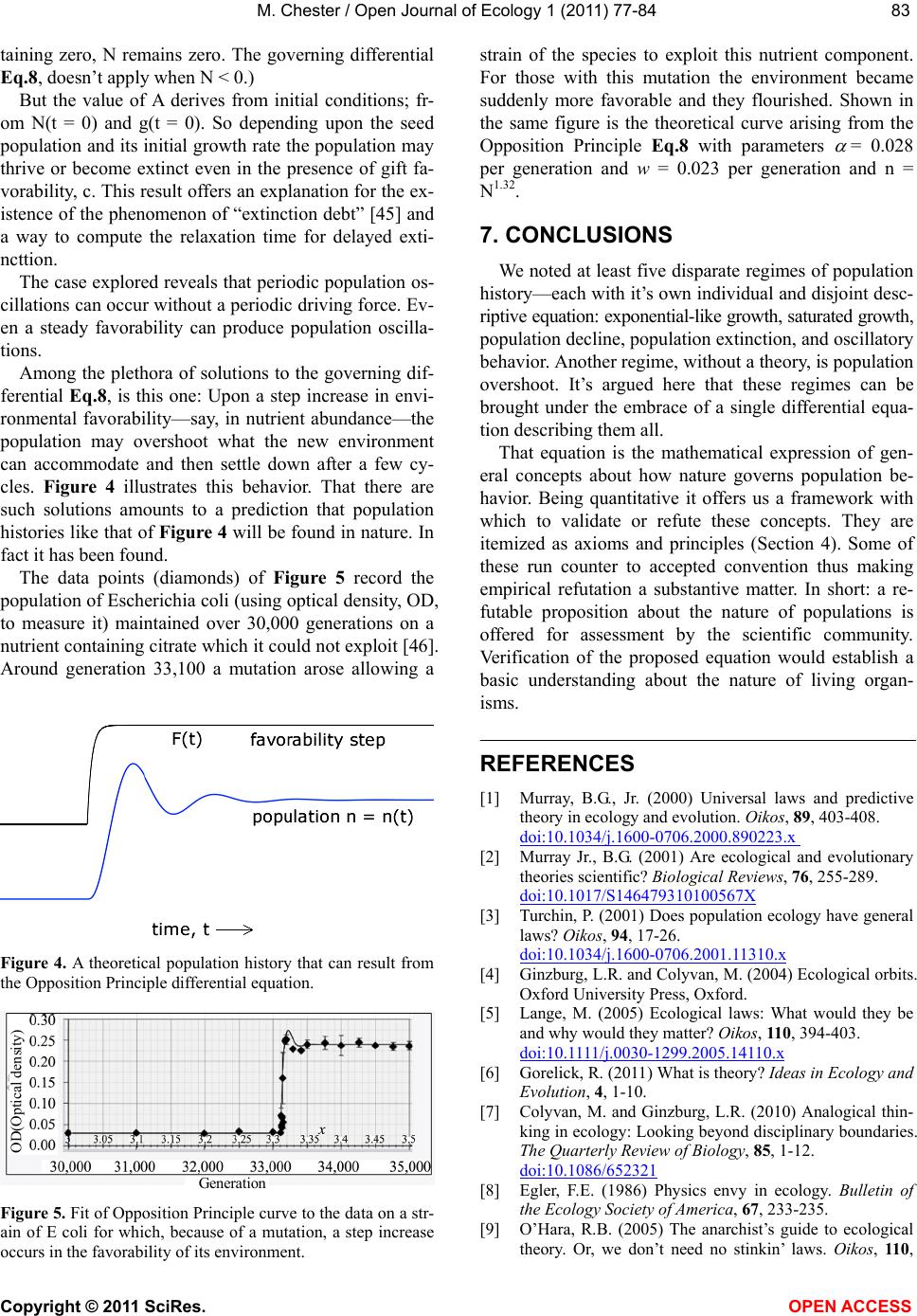
M. Chester / Open Journal of Ecology 1 (2011) 77-84
Copyright © 2011 SciRes. OPEN ACCESS
84
ology. Oikos, 103, 695-701.
390-393.
[10] Berryman, A. (2003) On principles, laws and theory in po-
pulation ec
doi:10.1034/j.1600-0706.2003.12810.x
[11] Lockwood, D.R. (2008) When logic fail
terly Review of Biology, 83, 57-64.
s ecology. Qua-
0.1086/529563doi:1
[12] Holt, R.D., (2009) Darwin, malthus and movement: A
hidden assumption in the demographic foundations of
evolution. Israel Journal of Ecology and Evolution, 55,
189-198. doi:10.1560/IJEE.55.3.189
[13] Lotka, A.J. (1956) Elements of mathematical biology.
Dover, New York.
[14] Volterra, V. (1926) Fluctuations in the abundance of a
species considered mathematically. Nature, 118, 558-560
doi:10.1038/118558a0
[15] Murray, J.D. (1989) Mathematical biology. Springer-Ver-
lag, Berlin.
[16] Vainstein, J.H., Rube, J.M. and Vilar, J.M.G. (2007) Sto-
chastic population dynamics in turbulent fields. Euro-
pean Physical Journal Special Topics, 146, 177-187.
doi:10.1140/epjst/e2007-00178-7
[17] Ginzburg, L.R. (1972) The analogies of the “free motio
and “force” concepts in populatio
n”
n theory (in Russian).
l Bi-
In: Ratner, V.A. Ed., Studies on theoretical genetics, Aca-
demy of Sciences of the USSR, Novosibirsk, 65-85.
[18] Ginzburg, L.R. (1986) The theory of population dynamo-
ics: I. Back to first principles. Journal of Theoretica
ology 122, 385-399. doi:10.1016 /S0022-5193(8 6)80180-1
[19] Ginzburg, L.R. (1992) Evolutionary consequences of ba-
sic growth equations. Trends in Ecology and Evolution, 7,
133. doi:10.1016/0169-5347(92)90149-6
[20] Ginzburg, L.R. and Taneyhill, D. (1995) Higher growth
rate implies shorter cycle, whatever the cause: A reply to
Berryman. Journal of Animal Ecology, 64, 294-295.
doi:10.2307/5764
[21] Ginzburg, L.R. and Inchausti, P. (1997) Asymmetry of p
pulation cycles: Ab
o-
undance-growth representation of hid-
den causes of ecological dynamics. Oikos, 80, 435-447.
doi:10.2307/3546616
[22] Britton, N.F., (2003) Essential mathematical biology. Sprin-
ger, London.
[23] Turchin, P. (2003) Complex population dynamics: A theo-
retical/empirical synthesis. Princeton University Press, Prin-
iley and Sons, New York.
s, Cambridge.
nviron-
ceton.
[24] Vandemeer, J., (1981) Elementary mathematical ecology.
John W
[25] Nowak, M.A. (2006) Evolutionary dynamics: Exploring
the equations of life. Harvard Pres
[26] Torres, J.-L., Pérez-Maqueo, O., Equihua, M. and Torres,
L. (2009) Quantitative assessment of organism-e
ment couplings. Biology and Philosophy, 24, 107-117.
doi:10.1007/s10539-008-9119-9
[27] Jones, J.M. (1976) The r-K-Selection Continuum. The Am
rican Naturalist, 110, 320-323.
e-
i:10.1086/283069do
[28] Ruokolainen, L., Lindéna, A., Kaitalaa, V. and Fowler,
M.S. (2009) Ecological and evolutionary dynamics under
coloured environmental variation. Trends in Ecology and
Evolution, 24, 555-563. doi:10.1016/j.tree.2009.04.009
[29] Okada, H., Harada, H., Tsukiboshi, T. and Araki, M.
(2005) Characteristics of Tylencholaimus parvus (Nema-
toda: Dorylaimida) as a fungivorus nematode. Nematol-
ogy, 7, 843-849. doi:10.1163/156854105776186424
[30] Ma, Z.S. (2010). Did we miss some evidence of chaos in
laboratory insect populations? Population Ecology, 53,
405-412. doi:10.1007/s10144-010-0232-7
[31] Carroll, S.B. (2006) The Making of the fittest. W.W. Nor-
ton, New York.
[32] Verhulst, P.-F., (1838) Notice sur la loi que la population
poursuit dans son accroissement. Correspondance Mathé-
7/BF00347974
matique et Physique, 10, 113-121.
[33] Parry, G.D. (1981) The meanings of r- and K-selection.
Oecologia, 48, 260-264. doi:10.100
r arti-
[34] Kuno, E. (1991) Some strange properties of the logistic
equation defined with r and K—inherent defects o
facts. Researches on Population Ecology, 33, 33-39.
doi:10.1007/BF02514572
[35] Gilbert, S.F. and Epel, D. (2009) Ecological developme
tal biology. Sinauer, Sunde
n-
rland.
isms and implica-
[36] Jablonka, E. and Raz, G. (2009) Transgenerational epige-
netic inheritance: Prevalence, mechan
tions for the study of heredity and evolution. Quarterly
Review of Biology, 84, 131-176. doi:10.1086/598822
[37] Clutton-Brock, T.H. (Ed.) (1990) Reproductive success:
Studies of individual variation in contrasting breeding
ating in-
systems. University of Chicago Press, Chicago.
[38] Coulson, T., Benton, T.G., Lundberg, P., Dall, S.R.X.,
Kendall, B.E. and Gaillard, J.-M. (2006) Estim
dividual contributions to population growth: Evolution-
ary fitness in ecological time. Proceedings of the Royal
Society B, 273, 547-555. doi:10.1098/rspb.2005.3357
[39] Owen-Smith, N. (2005) Incorporating fundamental laws
of biology and physics into population ecology: The me-
taphysiological approach. Oikos, 111, 611-615.
doi:10.1111/j.1600-0706.2005.14603.x
[40] Owen-Smith, R.N. (2002) Adaptive herbivore
Cambridge University Press, Cambridge
ecology.
.
doi:10.1017/CBO9780511525605
[41] Pelletier, F., Garant, D. and Hendry, A.P. (20
lutionary dynamics. Phililosophica 09) Eco-evo-
l Transactions Royal
Society B, 364, 1483-1489. doi:10.1098/rstb.2009.0027
[42] McNamara, J.M. and Houston, A.I. (2009) Integrating fun-
ction and mechanism. Trends in Ecology and Evolution,
24, 670-675. doi:10.1016/j.tree.2009.05.011
[43] Spencer, C.L. and Lensink, C.J. (1970) The muskox of Nu-
nivak Island, Alaska. The Journal of Wildlife Manage-
ment, 34, 1-15. doi:10.2307/3799485
[44] Turchin, P., Wood, S.N., Ellner, S.P., Kendall, B.E., Mur-
doch, W.W., Fischlin, A., Casas, J., McCauley, E. and Bri-
ggs, C.J. (2003) Dynamical effects of plant quality and
parasitism on population cycles of larch budmoth. Ecol-
ogy, 84, 1207-1214.
doi:10.1890/0012-9658(2003)084%5B1207:DEOPQA%
5D2.0.CO;2
[45] Kuussaari, M., Bommarco, R., Heikkinen, R.K., Helm,
A., Krauss, J.
, Lindborg, R., Öckinger, E., Pärtel, M., Pi-
no, J., Rodà, F., Stefanescu, C., Teder, T., Zobel, M. and
Ingolf, S.-D.I. (2009) Extinction debt: A challenge for
biodiversity conservation. Trends in Ecology and Evolu-
tion, 24, 564-571. doi:10.1016/j.tree.2009.04.011
[46] Blount, Z.D., Boreland, C.Z. and Lenski, R.E. (2008) His-
torical contingency and the evolution of a key innovation
in an experimental population of Escherichia coli. Pro-
ceedings of the National Academy of Science, 105, 7899-
7906. doi:10.1073/pnas.0803151105