Paper Menu >>
Journal Menu >>
 J. Biomedical Science and Engineering, 2011, 4, 591-598 JBiSE doi:10.4136/jbise.2011.49075 Published Online September 2011 (http://www.SciRP.org/journal/jbise/). Published Online September 2011 in SciRes. http://www.scirp.org/journal/JBiSE Mode of interaction of calcium oxalate crystal with human phosphate cytidylyltransferase 1: a novel inhibitor purified from human renal stone matrix Priyadarshini Pathak1, Pradeep Kumar Naik 1, Dipankar Sengupta1, Shrawan Kumar Singh2, Chanderdeep Tandon1* 1Biotechnology & Bioinformatics, Jaypee University of Information Technology, Waknaghat, H.P., India; 2Department of Urology, Post Graduate Institute of Medical Education and Research (PGIMER), Chandigarh, India. Email: *chanderdeep.tandon@juit.ac.in; tandonchanderdeep@yahoo.com Received 17 June 2011; revised 12 July 2011; accepted 10 August 2011. ABSTRACT Nephrolithiasis is a common clinical disorder, and calcium oxalate (CaOx) is the principal crystalline component in approximately 75% of all renal stones. It is widely believed that proteins act as inhibitors of crystal growth and aggregation. Acidic amino acids present in these proteins play a significant role in the inhibition process. In this study, interaction of cal- cium oxalate with human phosphate cytidylyltrans- ferase 1(CCT), a novel calcium oxalate crystal growth inhibitor purified from human renal stone matrix has been elucidated in silico and involvement of acidic amino acids in the same. As only sequence of CCT is available, henceforth its 3-D structure was modeled via Homology modeling using Prime module of Schrodinger package. Molecular dynamic simulation of modeled protein with solvation was done by mac- romodel (Schrodinger). The quality of modeled pro- tein was validated by JCSG protein structure valida- tion (PROCHECK & ERRAT) server. To analyze the interaction of modeled protein CCT with calcium oxalate along with role played by acidic amino acids, ‘Docking simulation’ was done using MOE–Dock. Interaction between calcium oxalate and CCT was also studied by substituting acidic amino acid in the active sites of the protein with neutral and positively charged amino acids. The in silico analysis showed the bond formation between the acidic amino acids and calcium atom, which was further substantiated when substitution of these acidic amino acids with alanine, glycine, lysine, arginine and histidine com- pletely diminished the interaction with calcium ox- alate. Keywords: Calcium Oxalate; Matrix-Assisted Laser Desorption/ I onization-Time of Flight (MALDI-TOF-MS); Antilithiatic; Human Phosphate Cytidylyltransferase 1; MOE–Dock. 1. INTRODUCTION Calcium oxalate is the most significant component of urin ary stones [1]. The mechanisms for the formation of calcium oxalate urinary stones are still not understood, though it is thought that organic macromolecules, in particular proteins, play a significant role. It is widely believed that proteins act as inhibitors of crystal growth and aggregation. Prothrombin fragment 1 (PFT1) [2], nephrocalcin [3], bikunin [4,5] and osteopontin [6] have all been shown to inhibit growth or aggregation or both in crystallization experiments. Nucleolin-related protein (NRP) is found on the surface of inner medullary collecting d uct (IMCD) cel ls in culture (cI MCD) and sele- ctively adsorbs to calcium oxalate (CaOx). NRP mediates attachment to the renal tubular epithelium of Ca stone crystals through an electro static interaction with a highly acidic region (acidic fragment (AF)) similar to those of other proteins that have been reported to affect urinary crystal formation. In vitro studies have shown that OPN is a potent inhibitor of COM growth, promotes the formation of calcium oxalate dihydrate (COD) rather than monohydrate, and inhibits the aggregation of COM crystals [7]. In vivo, OPN is the major component of the organic matrix of oxalate-containing kidney stones. OPN con- tains; 300 amino acids, including a conserved sequence of contiguous aspartic acid residues [8]. This polypep- tide chain undergoes extensive posttranslational modifi- cation, including glycosylation, phos-phorylation, and sulfation. The exact pattern of modification depends upon the species and tissue in which the protein is syn- thesized. The bovine milk isoform of OPN contains 27  P. Pathak et al. / J. Biomedical Science and Engineering 4 (2011) 591-598 Copyright © 2011 SciRes. JBiSE 592 sites of serine phosphorylation, 1 of threonine phos- phorylation, and 3 of O-linked glycosylation [9]. Human milk OPN is phos-phorylated at 34 serine and 2 threonine residues, and O-glycosylated at five sites [10]. The specific mechanism by which these proteins might interact with calcium oxalate surfaces is still in its infancy. Researchers working in other areas of biomin- eralization [11] have suggested that acidic amino acid residues such as Asp and Glu, that are expected to be deprotonated and negatively charged at urinary pH, are attracted to the po sitively charged calcium ions. It would therefore be expected that proteins rich in γ-carboxyglu- tamic acid, Gla, such as PFT1, with two deprotonated carboxylate groups would be even better at electrostati- cally binding to calcium sites [12,13]. Very recently, we have isolated and characterized a novel protein, human phosphate cytidylyltransferase 1, choline,beta (CCT) which is anionic in nature (MW 42 kDa) from organic matrix of calcium oxalate renal stones, which inhibits the growth of calcium oxalate [14]. It was identified by MALDI-TOF-MS followed by da- tabase search on MASCOT server as human phos-phate cytidylyltransferase 1, beta. Molecular weight of this novel CaOx crystal growth inhibitor from human renal stone matrix is also same as that of human phosphate cytidylyltransferase 1, choline, beta. It is involved in the biosynthesis of phosphatidylcholine which happens to be an important constituent of human renal stones and is also reported to have an antilithiatic effect. Amino acid sequence of the identified protein revealed, presence of acidic amino acid. The aim of the present work is to study the interaction of this novel CaOx crystal growth inhibitor (CCT) from human renal stone matrix with calcium oxalate at molecular level in silico. 2. METHODS 2.1. Sequence Alignments, Secondary Structure Prediction and Protein Fold Recognition Blast search of the human CCT sequence obtained from MALDI-TOF-MS and MASCOT was done using non- redundant Protein Data Bank for sequence alignment and further selection of appropriate template for secon- dary structure prediction and fold recognition . 2.2. Homology Modeling of Human CCT 1 Choline Beta Homology modeling of the identified human phosphate cytidylyltransferase 1, choline, bet a protei n s tructure wa s done by using Prime module of Schrodinger (Prime ver- sion 1.5, Macromodel version 9.1, Schrodinger, LLC, New York, NY, 2005) software. Considering the high degree of similarity of crystal structure of mammalian CTP: phosphocholine cytidylyltransferase (3HL4_A) of Rattus novergicus with CCT, it was taken as a model cytidylyltransferase (template) for studying structure- function relationships [15,16]. The structure of protein was modeled on the basis of its structural similarity with the chain A, crystal structure of a mammalian CTP: phosphocholine cytidylyltrans- ferase (Protein Data Bank ID: 3HL4) of Rattus nover- gicus as a template. The degree of identity between the template and the human CCT sequence was 53%, which enabled a preliminary model to be generated by Schrö- dinger. The sequence alignment was then improved manually and comparative homology method was used to build the structure of CCT. Macromodel program of Schrödinger software was used for molecular dynamic simulation. To check the quality of the modeled protein, JCSG protein structure validation serve r was u sed wh ere PROCHEK & ERRAT vali dation were done. 2.3. Docking of Homology Model of Human CCT1 Choline Beta with Calcium Oxalate Calcium oxalate structure was drawn with the help of molecular builder of Molecular Operating Environment (MOE) package developed by the Chemical Computing Group Inc. Montreal, Canada. Active site of the modeled protein (CCT) was predicted by using active site finder tool of MOE software. Then, the docking of modeled protein with calcium oxalate was done using MOE- Dock. MOE-dock utilizes a Monte Carlo Simulated An- nealing (SA) method in docking calculations to search for favorable binding configurations of a small, flexible ligand and a rigid macromolecule in a pre-set box. The docking energy calculation was carried out within a user- specified three-dimensional docking box (3D docking box) using the simulated annealing method under the OPLS-AA force field. The energy grids for docking were generated as grid-based potential fields by the MOE- Dock program, to reduce the calculation time. Each docking energy value was calculated as the sum value of the electrostatic, Van der Waals, and flexibility energies. The interaction energy was calculated using the electro- static and Van der Waals potential fields sampled on a grid overlaying the 3D docking box. The 3D docking box was interpolated at the atom positions by tri-linear interpolation. The Van der Waals parameters were taken from the currently active force field. The electrostatic field was calculated based on forcefield in the Coulom- bic manner using the constant dielectric of 1.0 for solva- tion. MOE-Dock performed 25 independent docking runs, and wrote the resulting conformations and their energies to a molecular database file. The lowest dock- ing energy conformation for each active site was chosen for LIGPLOT. 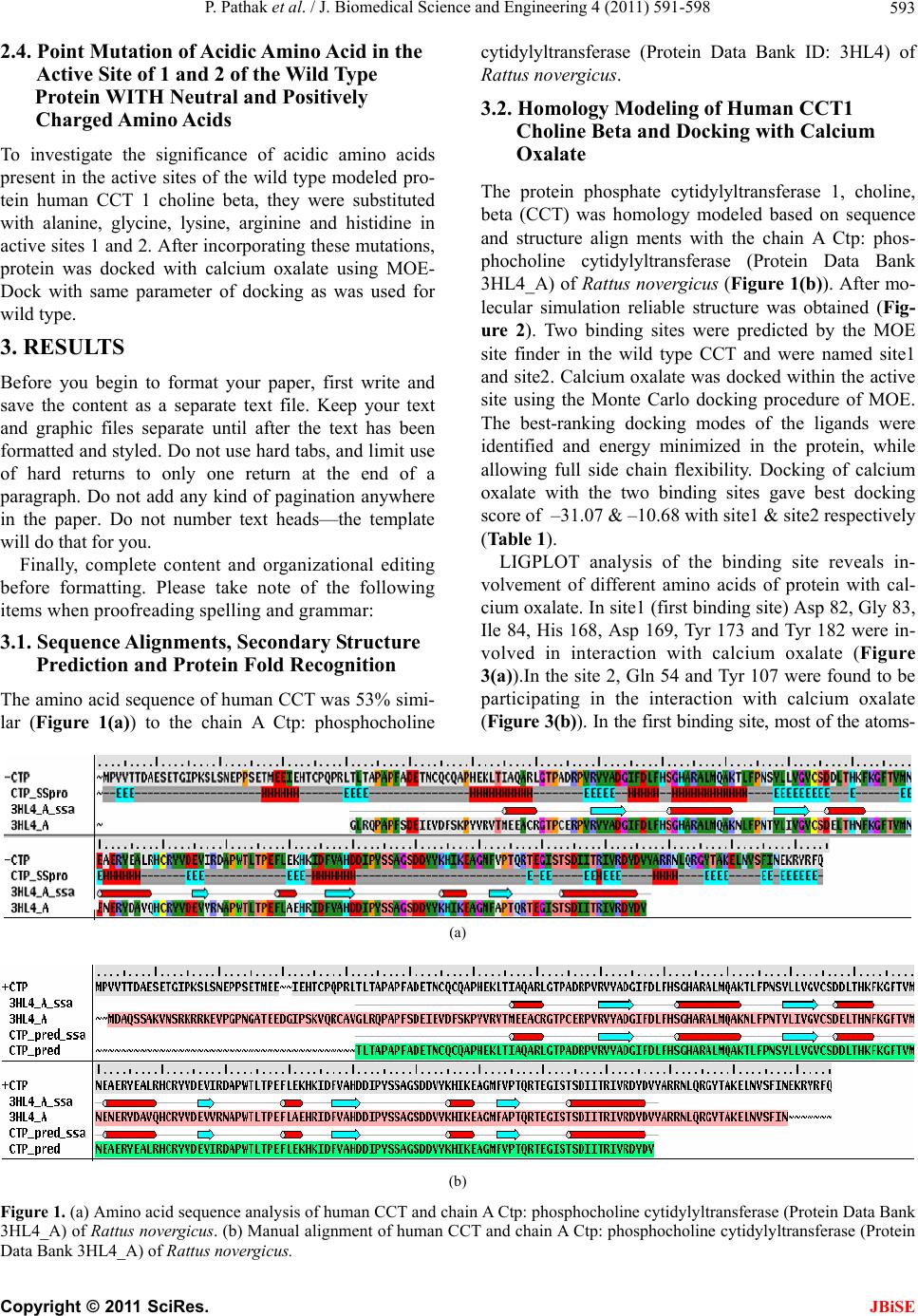 P. Pathak et al. / J. Biomedical Science and Engineering 4 (2011) 591-598 Copyright © 2011 SciRes. JBiSE 593 2.4. Point Mutation of Acidic Amino Acid in the Active Site of 1 and 2 of the Wild Type Protein WITH Neutral and Positively Charged Amino Acids To investigate the significance of acidic amino acids present in the active sites of the wild type modeled pro- tein human CCT 1 choline beta, they were substituted with alanine, glycine, lysine, arginine and histidine in active sites 1 and 2. After incorporating these mutations, protein was docked with calcium oxalate using MOE- Dock with same parameter of docking as was used for wild type. 3. RESULTS Before you begin to format your paper, first write and save the content as a separate text file. Keep your text and graphic files separate until after the text has been formatted and styled. Do not use hard tabs, and limit use of hard returns to only one return at the end of a paragraph. Do not add any kind of pagination anywhere in the paper. Do not number text heads—the template will do that for you. Finally, complete content and organizational editing before formatting. Please take note of the following items when proofreading spelling and gr ammar: 3.1. Sequence Alignments, Secondary Structure Prediction and Protein Fold Recognition The amino acid sequence of human CCT was 53% simi- lar (Figure 1(a)) to the chain A Ctp: phosphocholine cytidylyltransferase (Protein Data Bank ID: 3HL4) of Rattus novergicus. 3.2. Homology Modeling of Human CCT1 Choline Beta and Docking with Calcium Oxalate The protein phosphate cytidylyltransferase 1, choline, beta (CCT) was homology modeled based on sequence and structure align ments with the chain A Ctp: phos- phocholine cytidylyltransferase (Protein Data Bank 3HL4_A) of Rattus novergicus (Figure 1(b)). After mo- lecular simulation reliable structure was obtained (Fig- ure 2). Two binding sites were predicted by the MOE site finder in the wild type CCT and were named site1 and site2. Calcium oxalate was docked within the active site using the Monte Carlo docking procedure of MOE. The best-ranking docking modes of the ligands were identified and energy minimized in the protein, while allowing full side chain flexibility. Docking of calcium oxalate with the two binding sites gave best docking score of –31.07 & –10.68 with site1 & site2 respectively (Table 1). LIGPLOT analysis of the binding site reveals in- volvement of different amino acids of protein with cal- cium oxalate. In site1 (first binding site) Asp 82, Gly 83, Ile 84, His 168, Asp 169, Tyr 173 and Tyr 182 were in- volved in interaction with calcium oxalate (Figure 3(a)).In the site 2, Gln 54 and Tyr 107 were found to be participating in the interaction with calcium oxalate (Figure 3(b)). In the first binding site, most of the atoms- (a) (b) Figure 1. (a) Amino acid sequence analysis of human CCT and chain A Ctp: phosphocholine cytidylyltransferase (Protein Data Bank 3HL4_A) of Rattus novergicus. (b) Manual alignment of human CCT and chain A Ctp: phosphocholine cy tidylyltransferase (Protein Data Bank 3HL4_A) of Rattus novergicus.  P. Pathak et al. / J. Biomedical Science and Engineering 4 (2011) 591-598 Copyright © 2011 SciRes. JBiSE 594 Table1. Docking score of wild type human CCT model with calcium oxalate. W i ld Type Human CCT Site 1 Y80, D82, G83, I84, F85, D86, H89, G91, H92, R94, A95, Q98, C113, A149, P150, W151, T152, L153, H168, D169, Y173, S175, V181, Y182, T194, Q195, R196, T197, E198, G199, I200, S201, T202, S203, D204, R208 Site 2 A48, T51, N52, C53, Q54, A57, P58, H59, E60, K61, L62, Q66, T71, A73, D74, R75, P76, R78, Y107, L109, E144, I146, K160, H161, L162, I163 Docking Score –31.07 –10.68 Figure 2. Homology modeled structure of CCT. of calcium oxalate are highly exposed. Asp 82 and Tyr 182 are covalently bound to the calcium of the ligand (calcium oxalate), while Ile 84 is bound to the oxygen, where it is acting as side chain receptor (Figure 3(a)). In both of these active sites of the wild type modeled pro- tein human CCT 1 choline beta, it was observed that acidic amino acids played a significant role in the inter- action with calcium oxalate. From Ramchandran plot, we found that for the CCT structure predicted by homology modeling 89.7% of residues have their torsion angles in the most favored regions, while remaining 10.3% have in additional al- lowed regions (Figure 4(a)). Further quality of modeled protein was checked using Errat program which revealed an overall quality factor 74.66% (Figure 4(b)) before and 80.14% (Figure 4(c)) after molecular simulation. These results indicate that quality of modeled protein is highly reliable. 3.3. Point Mutation of Acidic Amino Acid of Active Site and Docking with Calcium Oxalate Upon substitution of acidic amino acids present in the active sites (site 1 and site 2) of the wild type modeled protein CCT with alanine, glycine, lysine, arginine and histidine, positive docking scores were obtained indicat- ing a poor interaction with calcium oxalate. No docking score was found when acidic amino acid present in binding site1was mutated to arginine and histidine. In all of these active sites of the wild type modeled protein CCT, it was observed that acidic amino acids played a significant role in the interaction with calcium oxalate. These studies clearly demonstrate that acidic amino ac- ids present in the wild type CCT showed a good docking score, an indicator of interaction with calcium an indi- cator of interaction with calcium oxalate, while substitu- tion of these acidic amino acids with alanine, glycine, lysine, arginine and histidine gave a poor docking score (Table 2). 4. DISCUSSION AND CONCLUSION Of all types of renal stones, calcium oxalate (CaOx) is the most common composition found by chemical analysis [17]. CaOx crystal growth inhibitors (proteins, lipids, glycosami-noglycans, and inorganic compounds) have been proposed to play an important role in renal stone disease [18,19]. During the last few years more and more research has been done at the cellular and mo- lecular levels. In spite of these advances however, the clinical treatment of urolithiasis remain far from satis- factory. Stone recurrence in human beings cannot be predicted an d is beyond the con trol of urologists, mainly because the mechanism of stone formation at molecular level is not yet fully understood [20]. Thus, determining the molecular mechanisms by which urinary con stituents modulate calcium oxalate crystallization is crucial for understanding and controlling urolithiassis in humans. Although a few initial molecular-scale investigations of the con-trolling mechanisms of kidney stone formation by these inhibitory molecules have been recently per- formed [21-24], the majority of previous studies have been concerned with the overall kinetics of crystallization, rather than molecular mechanisms which remain poorly defined. Therefore, in the present work, we have focused our attention on the interaction of the purified (in our lab) human phosphate cytidylyltransferase 1 (CCT) [14] with calcium oxalate using bi oi nf o rmatics tools. Cytidylyltransferases are critical enzymes involved in the biosyn-thetic pathways of lipids and complex carbo- hydrates. These enzymes catalyze a major step of energy input into biosynthesis by forming the activated inter- mediates, CDP-alcohols and CMP-sugars [25]. In fact CCT is involved in the biosynthesis of phosphatid ylcho- line which happens to be an important constituent of human renal stones and is also reported to have an anti-  P. Pathak et al. / J. Biomedical Science and Engineering 4 (2011) 591-598 Copyright © 2011 SciRes. JBiSE 595 lithiatic effect. Several cytidylyltransferases belong to a single family of structures, as defined by sequence similarities and sig- nature sequence that occur in their catalytic domains. Mammalian CCTα contains several functional regions: an N-terminal nuclear localization signal, a central cata- lytic domain, a membrane/lip id activation segment and a C-terminal phosphorylation region. There is a high de- gree of sequence similarity within the catalytic domain of all known forms of CCT, with the yeast catalytic do- main being 56% identical to that of mammalian CCTα, and the catalytic domains of CCTα, and CCTβ being 90% identical [16]. The degree of identity between the template and the human CCT sequence was 53%, which enabled a pre- liminary model to be generated by Schrodinger. Aspartic acid, isoleucine, tyrosine and glycine were found to be interacting with calcium oxalate at site1. More negative the docking score, stronger is the binding between ligand and protein’s active site [26]. The strong interaction be- tween CCT’s active site and calcium oxalate predicts inhibition of the same. Highest docking score (–31.079) (a)  P. Pathak et al. / J. Biomedical Science and Engineering 4 (2011) 591-598 Copyright © 2011 SciRes. JBiSE 596 (b) Figure 3. (a) Asp 82, Gly 83, Ile 84, His 168, Asp 169, Tyr 173 and Tyr 182 were involved in interact i o n with calcium o xalate. ( b ) In the site 2, Gln 54 and T yr 1 07 we re fou nd to be pa rt icipa ting in the interaction with calcium oxalate. with site1 indicates good binding of modeled protein with the ligand calcium oxalate. LIGPLOT of site1 showed involvement of aspartic acid (Asp) at position 82 with calcium while isoleucine (Ile) at position 84, inter- acts with oxygen of oxalate group. The binding site1 was found to be better for interaction with calcium oxalate compared to the other site. Whether a protein or other macromolecule acts as an inhibitor of growth and ag- gregation or a promoter of nucleation and aggregation implies that there must be some mechanism to explain the interaction with the mineral oxalate surfaces. The interaction between calcium and acidic Asp, Glu and Gla is certainly plausible, but it is equally conceivable that basic residues that are normally protonated at urinary pH and positively charged might experience an attraction toward negatively charged oxalate groups [27]. This is corroborated by the presence of basic amino acids too in the inhibitory proteins [28]. In either case steric cons- traints from 3D conformation of the molecule might limit the number of these simple interactions [27]. Posi- tive docking score with mutated binding sites confirms the inhibitory role of acidic amino acids [29-31].  P. Pathak et al. / J. Biomedical Science and Engineering 4 (2011) 591-598 Copyright © 2011 SciRes. JBiSE 597 (a) (b) (c) Figure 4. (a) In Ramchandran plot 89.7% of residues have their torsion angles in the most favored regions, while remain- ing 10.3% have in additional allowed regions. b & c Errat pro- gram which revealed an overall quality factor 74.66% (b) be- fore and 80.14% (c) after molecular simulation. There are reports that members of cytidylyltransferase family have similar conserved sequence [32]. In our study, we found 100% sequence in binding site1 and 29% sequence in site2, in the conserved region of CCT. Interestingly, conserved amino acids are contained within the catalytic core of the CCTs [32]. We found that the acidic amino acid is interacting with the calcium of calcium oxalate. This was further sub- Table 2. Docking score of mutated human CCT modelwith calcium oxalate. Amino acids with which point muta- tion is done at spe- cific positions in Site 1 & 2 Site 1 Point mutation of Aspartic acid at position 82, 86, 169, 204 & Glu- tamic acid at posi- tion 198 Site 2 Point mutation of Aspartic acid at position 74 & Glutamic acid at position 60 & 144 Alanine (A) 63.29 87.81 Glycine (G) 193.39 159.26 Ly s in e ( K) 98.33 138.05 Arginine (R) 105.27 151.53 Histidine (H) 163.29 153.29 stantiated when substitution of these acidic amino acids with alanine, glycine, lysine, arginine and histidine completely diminished the interaction with calcium ox- alate. These findings are in conformity with the presence of acidic amino acids in the various inhibitors of calcifi- cation from human beings [29-31] as well as acidic na- ture of antilithiatic proteins from plants [33,3 4]. 5. FUNDING We thank Jaypee University of Information Technology, Solan, HP, India for providing funds to carry out this research work. 6. ACKNOWLEDGEMENTS We express our gratitude to the Department of Urology, Post Graduate Institute of Medical Education and Research (PGIMER) Chandigarh, India for providing kidney stones. We would also like to thank Mr. Deeptak Ver ma for helping us with MOE simulation studies at Univer- sity of North Carolina, Charlotte. REFERENCES [1] Drach, G.W. (1992) Urinary lithiasis etiology, diagnosis, and medical management. Campbell’s Textbook of Urology, 2085. [2] Grover, P.K. and Ry all, R.L. (1998) Inhibition of calcium oxalate crystal growth and aggregation by prothrombin and its fragments in vitro. European Journal of Biology Chemistry, 263, 50. [3] Nakagawa, Y., Abram, V., Kezdy, F.J., Kaiser, E.T. and Coe, F.L. (1983) Purification and characterization of the principal inhibitor of calcium oxalate monohydrate crys- tal growth in human urine. The Journal of Biological Chemistry, 258, 12594-12600. [4] Atmani, F. and Khan, S.R. (1995) Characterization of uronic acid rich inhibitor of calcium oxalate crystalliza- tion isolated from rat urine. Urological Research, 23, 95. [5] Atmani, F., Mizon, J. and Khan, S. R. (1996) Identi- fication of uronic acid rich protein as urinary bikunin, the light chain of inter-a-inhibitor. European Journal of Bi- ology Chemistry, 236, 984. [6] Wesson, J.A., Johnson, R.J. and Mazzali, M.A, et al. (2003) Osteopontin is a critical inhibitor of calcium ox-  P. Pathak et al. / J. Biomedical Science and Engineering 4 (2011) 591-598 Copyright © 2011 SciRes. JBiSE 598 alate crystal formation and retention in renal tubules. Journal of the American Society Nephrology, 14 , 139- 147. [7] Taller, A., Grohe, B., Rogers, K.A., Goldberg, H.A. and Hunter, G.K. (2007) Specific adsorption of osteopontin and synthetic polypeptides to calcium oxalate monohy- drate crystals. Biophysical Journal, 93, 1768-1777. [8] Sodek, J., Ganss, B. and McKee, M.D. (2000) Osteo- pontin. Critical Reviews in Oral Biology and Medicine, 11, 279-303. [9] Sorensen, E.S., Hojrup, P. and Petersen, T.E. (1995) Post- translational modifications of bovine osteopontin: identi- fication of twenty-eight phosphorylation and three O-gly- cosylation sites. Protein Scienty, 4, 2040-2049. [10] Christensen, B., Nielsen, M.S., Haselmann, K.F., Peter- sen, T.E. and Sorensen, E.S. (2005) Posttranslationally modified residues of native human osteopontin are lo- cated in clusters: identification of 36 phosphorylation and five O-glycosylation sites and their biological implica- tions. Biochemical Journal, 390, 285-292. [11] Addadi, L. and Weiner, S. (1985) Interactions between acidic proteins and crystals: Stereochemical requirements in biomineralization. Proceedingof the Natlional Aca- demy Science of the USA, 82, 4110. [12] Lian, J.B., Prien, E.L., Glimcher, M.J. and Gallop, P.M. (1977) The presence of protein bound ccarboxyglutamic acid in calcium-containing renal calculi. Journal of Clinicine Investigation, 59, 1151. [13] Ry all, R.L., Fleming, D.E. and Grover, P.K. et al. (2000) The hole truth: Intracrystalline proteins and calcium ox- alate kidney stones. Molecular Urology, 4, 391. [14] Priyadarshini, Singh, S.K. and Tandon, C. (2009) Mass spectrometric identification of human phosphate cytidy- lyltransferase 1 as a novel calcium oxalate crystal growth inhibitor purified from human renal stone matrix. Clinica Chimica Acta, 408, 34-38. [15] Lee, J., Johnson, J., Ding, Z., Paetzel, M. and Cornell, R.B. (2009) Crystal structure of a mammalian CTP: phosphocholine cytidylyltransferase catalytic domain re- veals novel active site residues within a highly conserved nucleotidyltransferase fold. The Journal of Biological Chemistry, 284, 33535-33548. [16] Jon, A.F., Heidi, A.C. and Claudia Kent. (1999) Enzy- matic and cellular characterization of a catalytic fragment of CTP: Phosphocholine Cytidylyltransferase α. The Journal of Biological Chemistry, 274, 13384-13389. [17] Coe, F.L., Parks, J.H. and Asplin, J.R. (1992) The patho- genesis and treatment of kidney stones. New England Journal of Medicine, 327, 1141-1152. doi:10.1056/NEJM199210153271607 [18] Zerwekh, J.E., Holt, K. and Pak, C.Y. (1983) Natural urinary macromolecular inhibitors: attenuation of inhibi- tory activity by urate salts. Kidney International, 23, 838-841. [19] Coe, F.L., Parks, J.H. and Nakagawa, Y. (1991) Protein inhibitors of crystallization. Seminars in Nephrology, 11, 98-109. [20] Aggarwal, S., Tandon, C., Forouzandeh, M., Singla, S.K., Kiran, R. and Jethi, R.K. (2000) Role of biomolecules from human renal stone matrix on COM crystal growth. Molecular and Cellular Biochemistry, 210, 109-119. [21] Shirane, Y., Kurokawa, Y., Miyashita, S., Komatsu, H. and Kagawa, S. (1999) Study of inhibition mechanisms of glycosaminoglycans on calcium oxalate monohydrate crystals byatomic force microscopy. Urology Research, 27, 426-431. [22] Guo, S., Ward, M.D. and Wesson, J.A. (2002) Direct visualization of calcium oxalate monohydrate (COM) crystallization and dissolution with atomic force micros- copy (AFM) and the role of polymeric additives. Lang- muir, 18, 4284-4291. [23] Qiu, S.R., Wierzbicki, A. and Orme, C.A. et al. (2004) Molecular modulation of calcium oxalate crystallization by osteopontin and citrate. Proceedingof the Natlional Academy Science of the USA, 101, 1811-1815. [24] Jung, T., Sheng, X., Choi, C.K., Kim, W., Wesson, J.A. and Ward, M.D. (2004) Probing crystallization of cal- cium oxalate monohydrate and the role of macromole- cule additives with in situ atomic force microscopy. Langmuir, 20, 8587-8596. [25] Weber, C.H., Park, Y.S., Sanker, S., Kent, C. and Ludwig, M.L. (1999) A prototypical cytidylyltransferase: CTP: Glycerol-3 phosphate cytidylyltrnsferase from Bacillus subtilis. Structure 7, 9. [26] Bijarnia, R.K., Kaur, T., Naik, P.K., Singla, S.K. and Tandon C. (2008) In silico study on interaction of active binding sites of proteins with calcium oxalate monohy- drate. Office of James Bumett, 2, 92-107. [27] Gul, A. and Rez, P. (2007) Models for protein binding to calcium oxalate surfaces. Urology Research, 35, 63-71. [28] Tandon, C., Aggarwal, S., Forouzandeh, M. and Jethi, R.K. (1998) Inhibitors of in vitro mineralization from rabbit aorta and their role in biomineralization. Journal of Cellular Biochemistry, 68, 287-297. [29] Nakagawa, Y., Ahmed, M., Hall, S.L., Deganello, S. and Coe F.L. (1987) Isolation from human calcium oxalate renal stones of nephrocalcin, a glycoprotein inhibitor of calcium oxalate crystal growth. Evidence that nephr- ocalcin from patients with calcium oxalate nephr- olithi- asis is deficient in γ-carboxyglutamic acid. Journal of Clinical Investigation, 79, 1782-1787. [30] Nakagawa, Y., Abram, V., Parks, J.H., Lau, H.S.H., Ka- wooya, J.K. and Coe, F.L. (1985) Urine glycoprotein crystal growth inhibitors. Evidence for a molecular ab- normality in calcium oxalate nephrolithiasis. Journal of Clinical Investigation, 76, 1455-1462. [31] Shiraga, H., Min, W., VanDusen, W.J. et al. (1992) Inhi- bition of calcium oxalate crystal growth in vitro by uro- pontin: Another member of the aspartic acid rich protein superfamily. Proceedingof the Natlional Academy Scie- nce of the USA, 89, 426-430. [32] Kalmar, G.B., Kay, R.J., Lachance, A., Aebersold, R. and Cornell, R.B. (1990) Cloning and expression of rat liver CTP: Phosphocholine cytidylyltransferase: an amphi- pathic protein that controls phosphatidylcholine synthesis. Proceedingof the Natlional Academy Science of the USA, 87, 6029-6033. [33] Kaur, T., Bijarnia, R.K., Singla, S.K. and Tandon, C. (2009) Purification and characterization of an anticalci- fying protein from the seeds of trachyspermum ammi (L.). Protein and Peptides Letters, 16, 173-81. [34] Bijarnia, R.K., Kaur, T., Singla, S.K. and Tandon, C. (2009) A novel calcium oxalate crystal growth inhibitory protein from the seeds of Dolichos biflorus (L.). The Protein Journal, 28, 161-168. |