 World Journal of Neuroscience, 2011, 1, 9-18 doi:10.4236/wjns.2011.12002 Published Online August 2011 (http://www.SciRP.org/journal/wjns/ WJNS ). Published Online August 2011 in SciRes. http://www.scirp.org/journal/WJNS Neurogenesis-enhancing effect of sodium ferulate and its role in repair following stress-induced neuronal damage Lijian Yu1*, Yongping Zhang1, Mingneng Liao1, Yanping W ang1, Rundi Ma1, Xiaoyu Zhang1,2, Tingxi Yu1,3* 1Key Laboratory of Marine Materia Medica, Guangdong Ocean University, Zhanjiang, China; 2Department of Otorhinolaryngology-Head and Neck Surgery, University of Maryland School of Medicine, Baltimore, USA; 3Cell Biology Group, Department of Surgery, Department of Pathology, University of Maryland School of Medicine and Baltimore Veterans Affairs Medical Center, Baltimore, USA. Email: *yutingxi@yahoo.com, *ywyj9578@sohu.com Received 22 April 2011; revised 11 May 2011; accepted 18 June 2011. ABSTRACT Ferulic acid (FA) is a ubiquitous phenolic acid of low toxicity, and sodium ferulate (SF) is its sodium salt. Our previous studies have revealed that FA shows neuroprotective effect and significant antidepressant- like effect. The aim of this study was to investigate its potential neurogenesis-enhancing effect and its role in repair following stress-induced neuronal damage. MTT assay was performed to measure the effect of SF on the growth of rat pheochromocytoma (PC12) cells; morphological and immunocytochemical meth- ods were used for assessing its differentiation-induc- ing action. Chronic mild stress (CMS) tests were per- formed to establish rat model of depression. The histopathology of animal brains was studied to ana- lyze CMS-induced morphological changes and the effect of SF on the repair of CMS-induced brain in- jury. The expressions of nerve growth factor (NGF) and brain-derived neurotrophic factor (BDNF) and the proliferation of neural stem cell/neural progenitor cells were assessed in the hippocampi of chronic mild stress (CMS)-induced depression-like model rats by immunohistochemistry and bromodeoxyuridine (BrdU)- incorporation assays, respectively. Our in vitro tests showed that SF promoted the proliferation of PC12 cells in the concentration range of 5 - 320 µM, and induced PC12 cells to differentiate to more mature cells with the morphological characteristics and mo- lecular marker of neuronal-like cells. In vivo tests showed that SF up-regulated the expressions of NGF and BDNF, and induced the proliferation of neural stem cell/neural progenitor cells in the hippocampi of CMS-induced depression-like model rats. This study provides evidences that SF shows neurogenesis-en- hancing effect, and its antidepressant-like effect of SF may be related directly and closely to its above-men- tioned effect. Keywords: Sodium Ferulate; Neurogenesis-Enhancing Effect; Rat Pheochromocytoma (PC12) Cells; Stress- induced Neuronal Damage 1. INTRODUCTION Today, preclinical and clinical investigations have shown the involvement of dysregulation of hypothalamic-pi- tuitary-adrenal (HPA) axis in the pathogenesis of de- pression. Excessive release of glucocorticoid is closely related to the hippocampus atrophy. Hypercortisolemia and the associated hippocampal atrophy were observed in patients with depression, which could be ameliorated by the treatment with antidepressants [1]. Moreover, the volumes of the double-side hippocampus were reduced in the major depressive patients compared to the healthy control and there was a positive correlation between the hippocampus atrophy and the time course of the depress- sion [2,3]. Consequently, it is reasonable that, in some cases, psychopathology arises as a consequence of al- tered morphological structure of particular brain. There is growing evidence that adult animals continue to pro- duce new neurons in the dentate gyrus of hippocampus; neurogenesis in the adult dentate gyrus has been ob- served in all mammalian species examined to date, in- cluding humans [4]; stress causes a decrease of neuro- genesis in the dentate gyrus; and antidepressant treat- ment in turn stimulates the cell proliferation in the den- tate gyrus. Furthermore, the waning and waxing of neu- rogenesis in brain areas such as the dentate gyrus is proposed as a key factor in the descent into and recovery from clinical depression, respectively, and a decrease in neurogenesis could occur due to genetic factors and stress (especially because of the involvement of adrenal corticoids) [5]. Ferulic acid (FA), 3-(4-hydroxy-3-methoxyphenyl)-2-  L. J. Yu et al. / World Journal of Neuroscience 1 (2011) 9-18 10 propenoic acid (Figure 1(a)), is one of the most com- mon phenolic acids with low toxicity [6]. FA is waterin- soluble, but its sodium salt, sodium ferulate (SF) (Figure 1(b)), is water-soluble, stable, and can be prepared by chemical synthesis [7]. FA shows a lot of biological ac- tivities, including antioxidant [8,9]), anti-inflammatory [10,11], and hypotensive effect [12,13]. Our previous work demonstrates that SF shows obvi- ous neuro-protective effect [14]. The simultaneous ad- ministration of SF with monosodium glutamate (MSG) reverses the effects of MSG on behavior and histopa- thology in mice [15-17]; intracerebroventricularly ad- ministered SF mediates brain repair following MSG- induced exitotoxic neuronal damage in adult mice [18, 19]; and administration of SF also shows antidepressant- like effects in animal models of depression [14]. There- fore, it is interesting and reasonable to further investigate the neurogenesis-enhancing effect of SF in vitro and in vivo and to discuss their relationship with its antide- pressant-like effects. 2. MATERIALS AND METHODS 2.1. Drugs and Chemicals SF was purchased from Yaoyou Pharmaceutic Co. Ltd (Chongqing, China); fluoxetine (FLU) hydrochloride purchased from Eli Lilly and Company Limited (USA). Mouse anti-neuronfilament-200 (NF-200) monoclonal body was purchased from Wuhan Boster Biological Technology LTD (China), and TRICH-anti-mouse IgG purchased from Beijing Zhong Shan—Golden Bridge Biological Technology Co. LTD (China). Rabbit anti-rat polyclonal antibodies (anti-NGF and anti-BDNF) were purchased from Beijing Boisynthesis Biotechnology LTD (Beijing, China). 5-Bromo-2’-deoxyuridine(BrdU) was purchased from Roche Diagnostics GmbH (Boeh- ringer-Mannheim, Indianapolis, IN), and rat anti-BrdU monoclonal antibody purchased from Abcam (Hong Kong) Ltd. Biotinyllated horse anti-mouse antibody, avidin-biotin complex were purchased from Beijing Figure 1. Chemical structure of ferulic acid (a) and sodium ferulate (b). Biosynthesis Biotechnology Co. (China). DAB substrat kit was purchased from Invitrogen corporation. Dulbec- co’s Modified Eagle Medium (DMEM) and 3-(4,5-dime- thylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) were purchased from Sigma Chemical Co.. NCS was obtained from Sijiqing Biological Material Co. (Hang- zhou, China). Horse serum was purchased from Hyclone. All other chemicals used were of reagent grade. 2.2. Cell Line and Cell Culture A clonal cell line derived from a pheochromocytoma (PC12) of the rat adrenal medulla [20] was purchased from Shanghai Institute of Biochemistry and Cell Bio- logy, Chinese Academy of Sciences. Cells were grown in DMEM supplemented with 10% newborn calf serum (NCS), 5% horse serum and antibiotics (100 units/ml penicillin, 100 μg/ml streptomycin) in flasks precoated with collagen [21]. The culture was replaced after 2 months of passage by thawing a fresh aliquot of frozen cells. The culture were maintained in a humidified atmos- phere containing 5% CO2 at 37˚C. Cells in log phase growth were used in the experiments. 2.3. Animals Adult male Sprague-Dawley (SD) strain rats weighing 220 - 280 g (specific pathogen free) purchased from the experimental animal center of Guangdong Medical Col- lege (experimental animal license SCXKyue 2007-2008 A034, No.0001909; Zhanjiang, China), were used across all the experiments. They had free access to tap water and standard laboratory food unless otherwise stated. Housing conditions were controlled, temperature was maintained at 22˚C ± 1˚C with approximately 60% rela- tive humidity. They were kept on a reversed 12/12 h light/dark cycle (light 07:00 - 19:00 h). Animals were acclimated to the animal quarters for 1 week before any experimental procedure. All the animals were treated in compliance with “Guidance Suggestions (Instructions) for the Care and Use of Laboratory Animals” Issued on September 30, 2006 by The Ministry of Science and Technology of the People’s Republic of China. 2.4. MTT Assay The mitochondrial metabolism of MTT to its insoluble blue formazan was used for enumerating cells to assess the effects of SF, H2O2, and glucocorticoid on the growth of PC12 cells and the protective effects of SF against oxidative damage and glucocorticoid-induced neurotox- icity according to the methods of Hansen et al. [22]. Briefly, Single-cell suspensions were prepared and seeded onto 96 well microculture plates at 1.0 × 105 cells/ml (90 µl/well). Cells were cultured for 12 h before addition of drugs. Drugs were diluted into DMEM and added to C opyright © 2011 SciRes. WJNS  L. J. Yu et al. / World Journal of Neuroscience 1 (2011) 9-18 11 each well in a volume of 10 µl. Cells were incubated at 37˚C for the time indicated. MTT solution (5 mg/ml) was aliquoted to each well in a volume of 20 µl, and 5 h later 100 µl of the solubilization solution (10% SDS-5% isobutyl alcohol-0.012 M HCl (w/v/v)) was added into each well. The plates were allowed to stand overnight in the incubator in a humidified atmosphere. Absorbance at 570 nm was determined for each well using an ELISA reader. Control wells contained all of the agents pre- sented in the treated wells except the drug(s). Each ex- perimental point was performed in three replicates. Data were expressed as a percentage of untreated control cul- tures. 2.5. Effect of SF on the Growth of PC12 Cells PC12 cells were cultured with different concentrations of SF for 24 h. Cell viability was examined by MTT assay. 2.6. Differentiation Induction of PC12 Cells PC12 cells were cultured in DMEM with SF for the time indicated, and the effects of different concentrations (10 µM, 20 µM, 40 µM, 80 µM, 160 µM) of SF on morpho- logical and molecular changes were investigated at dif- ferent time points in PC12 cells. Cells were analyzed throughout the differentiation process (6 h and 1, 2, 3, 4, 5, 6, 7, 8, and 9 days), for neurite outgrowth, and for molecular marker NF-200. For neurite outgrowth, visual fields of 1 cm2 were randomly selected in each dish of SF-induced differenti- ated PC12 cells. Photographed images were generated for all fields at each time point (6 h and 1, 2, 3, 4, 5, 6, 7, 8, and 9 days) using Leica DM IRB photomicroscope [23]. For molecular marker NF-200 [24], the cells were submitted to the immunocytochemistry of the NF-200 [25,26] on the 7th day. Briefly, cells in the wells of the 24-wells plates were washed with PBS. Cells then were fixed at room temperature for 30 min. Nonspecific bind- ing sites were blocked with 5% NCS, 5% goat serum and 0.5% triton X-100 all in PBS for 1 hour. The pri- mary antibody was added to the cells after being appro- priately diluted in PBS and incubated overnight at +4˚C. 0.01 M PBS was added to the cells in control group in- stead of the primary antibody. Cells were washed with PBS 3 times for 10 minutes and the secondary antibodies were added after being appropriately diluted in PBS and left for one hour at room temperature. After adding the second secondary antibodiy for 1 h, cells were washed again in PBS 3 times for 10 minutes. Slides were stored in dark at 4˚C. Slides were later visualized using Leica inverted confocal microscope and Zeiss laser confocal microscope. 2.7. CMS Procedures The animals were assigned randomly into six matched groups (n = 12 animals in each group) based on sucrose consumption (1% sucrose solution) before onset of CMS: control, CMS, fluoxetine control (CMS + FLU), CMS + SF (10, 20, 40 mg/kg/d) groups. The stressed rats were ex- posed to CMS for 28 days; The rats in CMS + FLU group were exposed to CMS and received administration of FLU (2.0 mg/kg/d, ig, once-daily) for 28 days; the rats in CMS + SF groups were exposed to CMS and re- ceived administration of SF (10, 20, 40 mg/kg/d, ip, once-daily) respectively for 28 days. The control rats were given ordinary daily care and received ip admini- stration of normal saline instead of SF and FLU, for 28 days. The stressed and control rats were kept in different rooms to allow independent manipulation of their envi- ronments during the duration of the stress procedure. Control rats were housed together, while the stressed rats were housed singly. Most of the stressors were adapted from the procedure described by Willner and collaborators [27] and some stressors were included from Moreau and collaborators (e.g. empty water bottle, restricted food) [28]. Each week included 2 h of paired caging, 3 h of tilted cage (45 degrees), 18 h of food deprivation immediately followed by 1 h of restricted access to food (5 micropellets), 2 × 18 h of water deprivation immediately followed by 1 h exposure to an empty bottle, 21 h of wet cage (200 ml water in 100 g sawdust bedding), and 36 h of continuous light. Stressors were presented both during the rats’ ac- tive (dark) period and during the inactive (light) period. The same stressors were used in all experiments. 2.8. Immunohistochemistry NGF and BDNF levels. For in vivo analysis of NGF and BDNF levels, the whole brains of 4 unselected rat were dissected and removed 2 days after the last antidepres- sant treatment and inmediately fixed in 10% formalin in 0.1 M PBS (pH 7.4) at 4˚C for 2 days, and processed by paraffin embedding methods. Sections 4 μm thick were cut and processed for inmunohistochemistry. The im- munohistochemistry of NGF and BDNF on sections was performed using a streptavidin-peroxidase conjugate (SP) method according to the manufacturer’s instructions [29, 30]. Primary antibodies (see Chemicals) were diluted 1: 100. After incubation of primary antibodies at room temperature overnight, the sections were incubated with second antibodies 15 min at room temperature, and the reaction was visualized with DAB. Controls were made according to the same procedure, but omitting the pri- mary antibody. Sections obtained were matched for comparable hip- C opyright © 2011 SciRes. WJNS 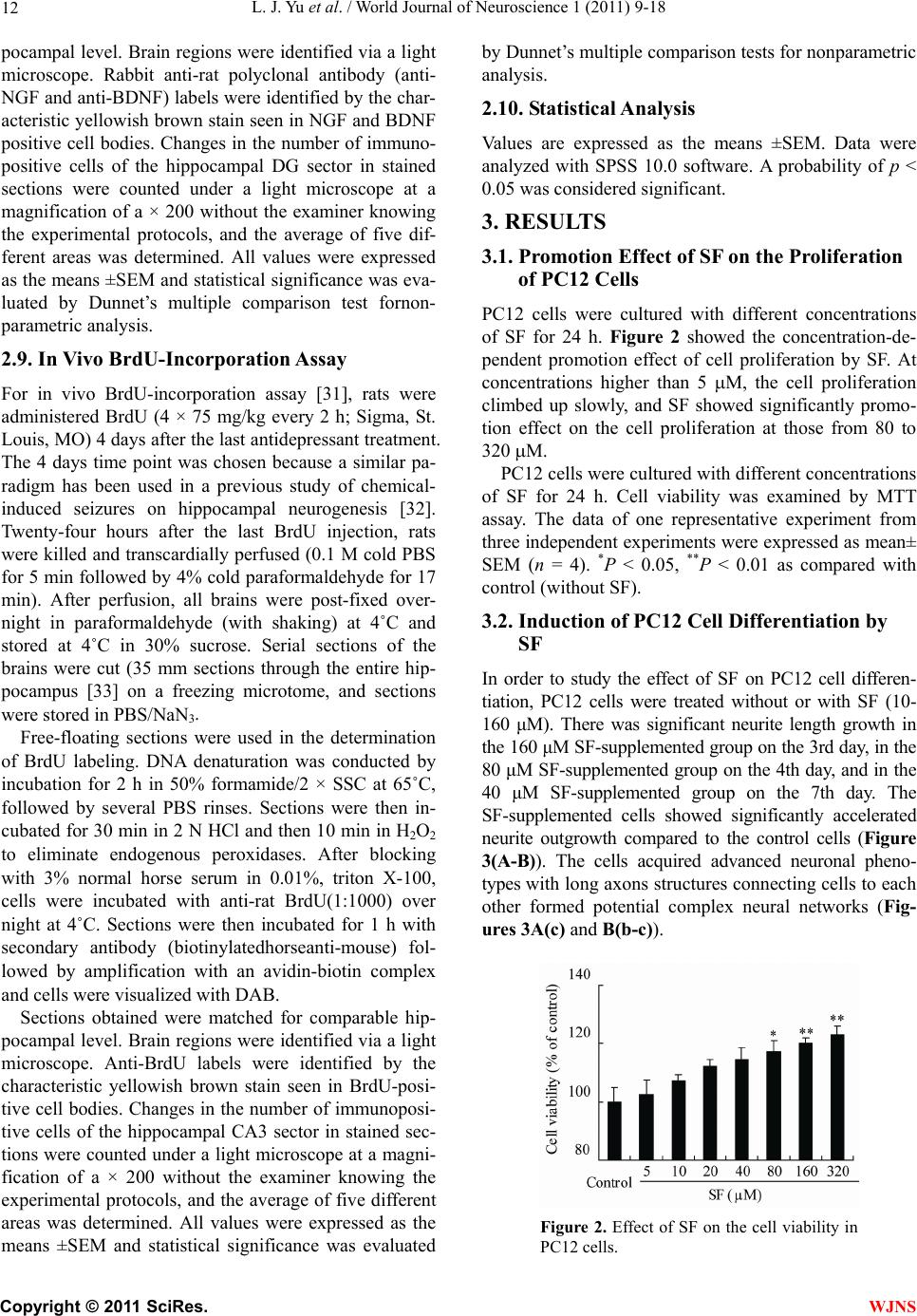 L. J. Yu et al. / World Journal of Neuroscience 1 (2011) 9-18 12 pocampal level. Brain regions were identified via a light microscope. Rabbit anti-rat polyclonal antibody (anti- NGF and anti-BDNF) labels were identified by the char- acteristic yellowish brown stain seen in NGF and BDNF positive cell bodies. Changes in the number of immuno- positive cells of the hippocampal DG sector in stained sections were counted under a light microscope at a magnification of a × 200 without the examiner knowing the experimental protocols, and the average of five dif- ferent areas was determined. All values were expressed as the means ±SEM and statistical significance was eva- luated by Dunnet’s multiple comparison test fornon- parametric analysis. 2.9. In Vivo BrdU-Incorporation Assay For in vivo BrdU-incorporation assay [31], rats were administered BrdU (4 × 75 mg/kg every 2 h; Sigma, St. Louis, MO) 4 days after the last antidepressant treatment. The 4 days time point was chosen because a similar pa- radigm has been used in a previous study of chemical- induced seizures on hippocampal neurogenesis [32]. Twenty-four hours after the last BrdU injection, rats were killed and transcardially perfused (0.1 M cold PBS for 5 min followed by 4% cold paraformaldehyde for 17 min). After perfusion, all brains were post-fixed over- night in paraformaldehyde (with shaking) at 4˚C and stored at 4˚C in 30% sucrose. Serial sections of the brains were cut (35 mm sections through the entire hip- pocampus [33] on a freezing microtome, and sections were stored in PBS/NaN3. Free-floating sections were used in the determination of BrdU labeling. DNA denaturation was conducted by incubation for 2 h in 50% formamide/2 × SSC at 65˚C, followed by several PBS rinses. Sections were then in- cubated for 30 min in 2 N HCl and then 10 min in H2O2 to eliminate endogenous peroxidases. After blocking with 3% normal horse serum in 0.01%, triton X-100, cells were incubated with anti-rat BrdU(1:1000) over night at 4˚C. Sections were then incubated for 1 h with secondary antibody (biotinylatedhorseanti-mouse) fol- lowed by amplification with an avidin-biotin complex and cells were visualized with DAB. Sections obtained were matched for comparable hip- pocampal level. Brain regions were identified via a light microscope. Anti-BrdU labels were identified by the characteristic yellowish brown stain seen in BrdU-posi- tive cell bodies. Changes in the number of immunoposi- tive cells of the hippocampal CA3 sector in stained sec- tions were counted under a light microscope at a magni- fication of a × 200 without the examiner knowing the experimental protocols, and the average of five different areas was determined. All values were expressed as the means ±SEM and statistical significance was evaluated by Dunnet’s multiple comparison tests for nonparametric analysis. 2.10. Statistical Analysis Values are expressed as the means ±SEM. Data were analyzed with SPSS 10.0 software. A probability of p < 0.05 was considered significant. 3. RESULTS 3.1. Promotion Effect of SF on the Proliferation of PC12 Cells PC12 cells were cultured with different concentrations of SF for 24 h. Figure 2 showed the concentration-de- pendent promotion effect of cell proliferation by SF. At concentrations higher than 5 M, the cell proliferation climbed up slowly, and SF showed significantly promo- tion effect on the cell proliferation at those from 80 to 320 M. PC12 cells were cultured with different concentrations of SF for 24 h. Cell viability was examined by MTT assay. The data of one representative experiment from three independent experiments were expressed as mean± SEM (n = 4). *P < 0.05, **P < 0.01 as compared with control (without SF). 3.2. Induction of PC12 Cell Differentiation by SF In order to study the effect of SF on PC12 cell differen- tiation, PC12 cells were treated without or with SF (10- 160 μM). There was significant neurite length growth in the 160 μM SF-supplemented group on the 3rd day, in the 80 μM SF-supplemented group on the 4th day, and in the 40 μM SF-supplemented group on the 7th day. The SF-supplemented cells showed significantly accelerated neurite outgrowth compared to the control cells (Figure 3(A-B)). The cells acquired advanced neuronal pheno- types with long axons structures connecting cells to each other formed potential complex neural networks (Fig- ures 3A(c) and B(b-c)). Figure 2. Effect of SF on the cell viability in PC12 cells. C opyright © 2011 SciRes. WJNS  L. J. Yu et al. / World Journal of Neuroscience 1 (2011) 9-18 13 Figure 3. Neurite outgrowth in the presence of SF. PC12-cells were grown in the absence (control) or presence of SF (80, 160 µM). Neurite outgrowth was monitored for 9 days, starting 6 h post-sup- plementation of SF. Representative micrographs of PC12-cell culture are shown (×200). A. 4rd day: a. control, b. 80 M, c. 160 M; B. 7th day: a. control, b. 80 M, c. 160 M. To determine whether the PC12 cells can synaptically integrate and mature in the culture with SF, we examined the expression of NF-200, marker of neuronal differen- tiation. After 3 - 4 days of incubation cells lost their per- fect round shape and small projections start to grow from these cells. After a week of differentiation in the presence of SF, cells showed polar morphology, axons and dendrites structures were clearly noticed and poten- tial simple neural network was noticed (Figures 3A(c) and B(b-c)). The differentiation process was accom- panied with apoptosis of some differentiation cells. The differentiating cells formed a much complex potential neural networks (Figure 4(c-d)). NF-200 (Red) highly expressed and was highly concentrated along the axon structures of the neuronal cells (Figure 4(d)). Staining with anti-NF-200 antibody showed that NF-200 expre- ssion was evident and PC12 cells underwent a process of maturation. The results revealed that under the action of SF, PC12 cells could be induced to differentiate to more mature cells with the morphological characteristics and molecu- lar marker of neuronal-like cells. 3.3. Protective Effect of SF against CMS-Induced Reduction of Hippocampal NGF and BDNF Levels Hippocampal NGF and BDNF levels were obviously reduced in the CMS mice (Figures 5A(a-b) and B(a-b)). In contrast, SF treatment reversed the effects of CMS on the hippocampal NGF and BDNF levels in a dose-de- pendent manner (Figures 5A(a, d, e, f) and B(a, d, e, f)), and the effect of SF(40 mg/kg/d) was comparable with that of FLU (2.0 mg/kg/d) control (Figures 5A(c, e, f) and B(c, e, f)). Consequently, the results suggest that SF Figure 4. Neuronal-like differentiated and maturated cells (×200). PC12-cells were grown in the absence (control) or presence of SF (80, 160 µM) for 9 days. For molecu- lar marker NF-200, the cells were submitted to the immunocytochemistry of the NF-200 on the 7th day as described in “Materials and methods”. (a) control; (b) control, using PBS instead of primary antibody (NF-200 monoclonal body); (c) PC12 cells treated with 160 μM SF for 7 days. The cells acquired ad- vanced neuronal-like phenotypes with long axons structures connecting cells to each other forming po- tential complex neural networks; (d) PC12 cells treated with 160 μM SF for 7 days were submitted to the immunocytochemistry of the NF-200. The cells with neuronal-like morphology after differentiation and maturation highly expressed NF-200 (red). treatment partially reverses the effects of CMS on hip- pocampal NGF and BDNF. The whole brains of unselected rat (n = 4) were dis- sected and removed 2 days after the last antidepressant treatment and inmediately fixed in 10% formalin, and processed by paraffin embedding methods. Sections 4 μm thick were cut and processed for inmunohistochem- istry. The immunohistochemistry of NGF and BDNF on sections was performed using a streptavidin-peroxidase conjugate (SP) method according to the manufacturer’s instructions. Rabbit anti-rat polyclonal antibody (anti- NGF and anti-BDNF) labels were identified by the char- acteristic yellowish brown stain seen in NGF and BDNF positive cell bodies. Changes in the number of im- munopositive cells of the hippocampal DG sector in stained sections were counted under a light microscope at a magnification of a × 200, and the average of five different areas was determined. All values were ex- pressed as the means ±SEM, and statistical significance was evaluated by Dunnet’s multiple comparison tests for nonparametric analysis. 3.4. Protective Effect of SF against the Reduction of Neurogenesis in the Hippocampal CA3 Sector of CMS-Induced Depression-Like Model Rats Analysis of the number of BrdU-labeled cells demon- strated that the number of BrdU-labeled cells was obvi- ously reduced in the CA3 of CMS mice (Figure 6(a-b)). n contrast, long-term administration of SF (10, 20, 40 I C opyright © 2011 SciRes. WJNS  L. J. Yu et al. / World Journal of Neuroscience 1 (2011) 9-18 Copyright © 2011 SciRes. 14 (A) (B) Figure 5. Representative microphotographs and numbers of NGF(A)- and BDNF(B)-immunopositive neurons in the hippocampal DG sector of CMS-induced depression-like model rats. (A) Representative microphotographs and numbers of NGF-immunopositive neurons (×200). a. Control; b. CMS; c. CMS + FLU (2.0 mg/kg); d. CMS + SF(10 mg/kg); e. CMS + SF(20 mg/kg); f. CMS+; (B) Representative microphotographs and numbers of BDNF-immunopositive neurons (×200). a. Control; b. CMS; c. CMS + FLU (2.0 mg/kg); d. CMS + SF(10 mg/kg); e. CMS + SF(20 mg/kg); f. CMS + SF(40 mg/kg). mg/kg/d) significantly increased the number of BrdU- labeled cells by 8.6%, 27.8%, 47.7% in the CA3 of CMS mice in a dose-dependent manner (Figure 6(b, d, e, f)), respectively, whereas FLU (2.0 mg/kg/d) increased the BrdU labeling by 23.2% (Figure 6(b-c)). Consequently, the results suggest that SF treatment reverses partially the effects of CMS on the neurogenesis in the hippo- campi of the rats, and the effects of SF(20, 40 mg/kg/d) were comparable with that of FLU (2.0 mg/kg/d) con- trol. For in vivo BrdU-incorporation assay, rats were ad- ministered BrdU(4 × 75mg/kg every 2 hr) 4 days after the last antidepressant treatment. Twenty-four h after the last BrdU injection, rats were killed and transcardially perfused. After perfusion, all brains were post-fixed overnight in paraformaldehyde (with shaking) at 4˚C and stored at 4˚C in 30% sucrose. Immunohistochemistry as- say was performed as “Materials and Methods”. Anti- BrdU labels were identified by the characteristic yellow- ish brown stain seen in BrdU-positive cell bodies. Chan- ges in the number of immunopositive cells of the hippo- campal CA3 sector in stained sections were counted under a light microscope at a magnification of a × 400 without the examiner knowing the experimental protocols, and the average of five different areas was determined. All values were expressed as the means ±SEM and statistical sig- nificance was evaluated by Dunnet’s multiple comparison tests for nonparametric analysis. 4. DISCUSSION Our previous investigations reveal that acute administra- tion of SF significantly decrease the duration of immo- WJNS  L. J. Yu et al. / World Journal of Neuroscience 1 (2011) 9-18 15 Figure 6. SF induces the proliferation of neural stem cell/ neural progenitor cells in the hippocampal CA3 sector of CMS-induced depression-like model rats (×200). a. Control; b. CMS; c. CMS + FLU(2.0 mg/kg/d); d. CMS + SF(10 mg/kg/d); e. CMS + SF (20 mg/kg/d); f. CMS + SF(40 mg/kg/d). bility during forced-swimming test and tail-supension test in mice and rats, suggesting that SF has an acute antidepressant-like effect. However, SF has no any ef- fects on reserpine-induced hypothermia, 5-HTP-induced head-twitch response, and potentiation of noradrenaline toxicity in mice, suggesting that the mechanisms of SF antidepressant-like effects are different from those of main kinds of antidepressants that have been employed in the treatment of depression for several decades. Long- term ip administration of SF reverses the effects of CMS on consumption of food and sucrose solution, weight gain, locomotion and exploration behavior, and signifi- canly shortens the immobility time during forced-swim- ming test following CMS in rats, suggesting that SF has a chronic antidepressant-like effect [14]. The present results obtained from in vitro tests show that SF treatment promotes the proliferation of PC12 cells in the concentration range of 5 - 320 µmol/L, and induce PC12 cells to differentiate to more mature cells with the morphological characteristics and molecular marker of neuronal-like cells. The present results ob- tained from in vivo tests show that SF treatment may up- regulate the expressions of NGF and BDNF, and induce the proliferation of neural stem cell/neural progenitor cells in the hippocampi of CMS-induced depression-like model rats. NGF is a well known characterized member of the neurotrophin family that stimulates differentiation, main- tains phenotype, and ensures the survival of various populations of neurons in the central nervous system (CNS) [34,35]. The loss of NGF has been implicated in the loss of cholinergic tone and function [36-38]. An- other neurotrophic factor, the BDNF, does not only pro- mote neuronal survival anddifferentiation, but also regu- lates synaptic neurotransmission and plasticity in the CNS [39,40]. Furthermore, BDNF levels are increased in the hippocampal regions of mice and rats with access to exercise wheels and trophic factor gene therapy can ameliorate hippocampal deficits in monkey [41,42]. NGF and BDNF are responsible for the development, differentiation, maintenance and repair of neurons [43- 45]. Exogenous NGF rescues cholinergic neurons in the basal forebrain and improves cognitive function in im- paired, aged or cholinergically lesioned animals [46-48]. A recent report shows that transplantation of fibroblasts genetically modified tosecrete NGF led to a slowing of cognitive decline in a Phase I clinical trial performed on patients suffering from mild Alzheimer’s disease [49]. BDNF is highly present in the dentate gyrus and hippo- campal CA3 sector, and to a lesser extent in the hippo- campal CA1 sector [50]. It has been reported that the extent of dendritic arborization in the hippocampus was profoundly increased in BDNF transgenic mice com- pared to their control litter mates [51]. In BDNF gene knockout mice, furthermore, long-term potential (LTP) in the hippocampal CA1 region is severely reduced in hippocampal neurons and spatial learning is significantly impaired [52,53]. The results obtained from our experi- ments show that the hippocampal NGF and BDNF levels were obviously reduced in the CMS mice. Of interest is that SF treatment reversed the effects of CMS on the hippocampal NGF and BDNF levels in a dose-dependent manner. This finding suggests that SF-induced increases of the hippocampal NGF and BDNF levels may play a role in the antidepressant-like effect of SF. The hippocampus is one of only a few brain regions where production of neurons occurs throughout the life time of animals, including humans [4]. Hippocampal C opyright © 2011 SciRes. WJNS  L. J. Yu et al. / World Journal of Neuroscience 1 (2011) 9-18 16 neurogenesis can be influenced by several environmental factors and stimuli [54-57]. It has been shown that stressful experiences, including both physical and psy- chosocial stress, suppress the formation of hippocampal granule cells in a number of mammalian species [58-60]. Decreased cell proliferation has also been reported in response to both acute and chronic stress paradigms [61]. Importantly, it has been shown that chronic antidepres- sant treatment signicantly increases the number of BrdU- labeled cells in the dentate gyrus and hilus of the hippo- campus. BrdU is incorporated into the newly synthesized DNA of S-phase cells and may provide an estimate for the fraction of cells in S-phase. A specific marker like BrdU is not only helpful for gaining further insights into the genesis of new neurons in the hippocampus, but also might be applicable to the development of strategies for therapeutic interventions [62]. Administration of sev- eral different classes of antidepressant, but not non-an- tidepressant, agents was found to increase BrdU-labeled cell number, indicating that this is a common and selec- tive action of antidepressants [31]. Of interest is that the results obtained from our experiments show that admini- stration of SF increase BrdU-labeled cell number in the hippocampi of CMS-induced depression-like model rats. This finding raise the possibility that increased cell pro- liferation and increased neuronal number induced by SF in CMS-induced depression-like model rats may be a mechanism by which SF treatment overwhelms the stress- induced atrophy and loss of hippocampal neurons. 5. ACKNOWLEDGEMENTS The authors would like to thank Dr. Depu Yu for his great encourage- ment and continuous promotion. REFERENCES [1] Mao, Q.Q., Ip, S.P., Ko, K.M., Tsai, S.H., Zhao, M. and Che, C.T. (2009) Peony glycosides protect against corti- cos-terone-induced neurotoxicity in PC12 Cells. Cellular and Molecular Neurobiology, 29, 643-647. doi: 10.1007/s10571-009-9357-7 [2] Sapolsky, R.M. (2000) Glucocorticoids and hippocampal atrophy in neuropsychiatric disorders. Archives of Gene- ral Psychiatry, 57, 925-935. doi: 10.1001/archpsyc.57.10.925 [3] Magarinos, A.M., McEwen, B.S., Flugge, G. and Fuchs, E. (1996) Chronic psychosocial stress caused apical den- dritic atrophy of hippocampal CA3 pyramidal neurons in subordinate tree shrews. The Journal of Neuroscience, 16, 3534-3540. [4] Eriksson, P.S., Perfilieva, E., Björk-Eriksson, T., Alborn, A.M., Nordborg, C., Peterson, D.A. and Gage, F.H. (1998) Neurogenesis in the adult human hippocampus. Nature Medicine, 4, 1313-1317. doi:10.1038/3305 [5] Jacobs, B.L. (2002) Adult brain neurogenesis and de- pression. Brain, Behavior, and Immunity, 16, 602-609. doi:10.1016/S0889-1591(02)00015-6 [6] Ji, Y.B. (1999) Pharmacological action and application of blood-activating and stasis-eliminating available compo- sition of traditional Chinese medicine. Heilongjiang Sci- ence and Technique Press, Harbin, 118-121. [7] Jing, H., Yao, L.Y., Li, J.S., Song, Y.Q. and Chao, W. (2002) Research progress of pharmacology of sodium ferulate. Northwest Pharmaceutical Journal, 17, 236- 238. [8] Grafe, E. (1992) Antioxidant potential of ferulic acid. Free Radical Biology and Medicine, 13, 435-448. doi:10.1016/0891-5849(92)90184-I [9] Scott, B.C., Butler, J., Halliwell, B. and Aruoma, O.I. (1993) Evaluation of the antioxidant actions of ferulic acid and catechins. Free Radical Research, 19, 241-253. doi:10.3109/10715769309056512 [10] Fernandez, M.A., Saenz, M.T. and Garcia, M.D. (1998) Anti-inflammatory activity in rats and mice of phenolic acids isolated from Scrophularia frutescens. Journal of Pharmacy and Pharmacology, 50, 1183-1186. doi:10.1111/j.2042-7158.1998.tb03332.x [11] Ozaki, Y. (1992) Antiinflammatory effect of tetramethyl- pyrazine and ferulic acid. Chemical & Pharmaceutical Bulletin, 40, 954-956. [12] Suzuki, A., Kagawa, D., Ochiai, R., Tokimitsu, I. and Saito, I. (2002a) Green coffee bean extract and its me- tabolites have a hypotensive effect in spontaneously hy- pertensive rats. Hypertens i o n R e s e arch, 25, 99-107. doi:10.1291/hypres.25.99 [13] Suzuki, A., Kagawa, D., Fujii, A., Ochiai, R., Tokimitsu, I. and Saito, I. (2002b) Short- and long-term effects of ferulic acid on blood pressure in spontaneously hyper- tensive rats. American Journal of Hypertension, 15, 351- 357. doi:10.1016/S0895-7061(01)02337-8 [14] Zhang, Y.P., Yu, L.J., Wang, Y.P., Liao, M.N., Ma, R.D., Zhang, X.Y. and Yu, T.X. (2011) Neuroprotective` effects of sodium ferulate and its antidepressant-like effect measured by acute and chronic experimental method in animal model of depression. Journal of Behavioral and Brain Science, 1, 37-46. [15] Yu, T.X., Zhao, Y., Shi, W.C., Ma, R.D. and Yu, L.J. (1997) Effects of maternal oral administration of mono- sodium glutamate at a late stage of pregnancy on devel- oping mouse fetal brain. Brain Research, 747, 195-206. doi:10.1016/S0006-8993(96)01181-X [16] Yu, L.J., Zhang, Y.P., Ma, R.D., Bao, L., Fang, J.Z. and Yu, T.X. (2006) Potent protection of ferulic acid against excitotoxic effects of maternal intragastric administration of monosodium glutamate at a late stage of pregnancy on developing mouse fetal brain. European Neuropsycho- pharmacology, 16, 170-177. doi: 10.1016/j.euroneuro.2005.08.006 [17] Zhang, Y.P., Yu, L.J., Ma, R.D., Bao, L., Zeng, R., Fang, J.Z., Zhang, X.Y. and Yu, T.X. (2008a) Potent protective effect of ferulic acid on glutamate-induced neurotoxicity in adult mice. Chinese Journal of Neuromedicine, 7, 596- 599. [18] Yang, P.P., Yu, L.J., Ma, R.D., Zhang, Y.P., Zhang, X.Y. and Yu, T.X. (2008) Intracerebroventricularly adminis- tered sodium ferulate-mediateed brain repair following C opyright © 2011 SciRes. WJNS  L. J. Yu et al. / World Journal of Neuroscience 1 (2011) 9-18 17 glutamate-induced exitotoxic neuronal damage in adult mice. Chinese Journal of Behavioral Medical Science, 17, 13-15. [19] Zhang, Y.P., Yu, L.J., Ma, R.D., Zhang, X.Y. and Yu, T.X. (2008b) Effects of administered sodium ferulate on the repair of glutamate-induced excitotoxic neuronal impair in mouse fetal brain. Chinise Journal of Behavioral Medical Science, 17, 968-971. [20] Shafer, T.J. and Atchison, W.D. (1991) Transmitter, ion channel and receptor properties of pheochromocytoma (PC12) cells: A model for neurotoxicological studies. Neurotoxicology, 12, 473-492. [21] Yoshizumi, M., Kogame, T., Suzaki, Y., Fujita, Y., Kyaw, M., Kirima, K., Ishizawa, K., Tsuchiya, K., Kagami, S. and Tamaki, T. (2002) Ebselen attenuates oxidative stress-induced apoptosis via the inhibition of the c-Jun N-terminal kinase and activator protein-1 signalling pathway in PC12 cells. British Journal of Pharmacology, 136, 1023-1032. doi:10.1038/sj.bjp.0704808 [22] Hansen, M.B., Nielsen, S.E. and Berq, K. (1989) Re- examination and further development of a precise and rapid dye method for measuring cell growth/cell kill. Journal of Immunological Methods, 11 9, 203-210. doi:10.1016/0022-1759(89)90397-9 [23] Dagai, L., Peri-Naor, R. and Birk, R.Z. (2009) Docosa- hexaenoic acid significantly stimulates immediate early response genes and neurite outgrowth. Neurochemical Research, 34, 867-75. doi:10.1007/s11064-008-9845-z [24] Julien J.P. and Mushynski W.E. (1998) Neurofilaments in health and disease. Progress in Nucleic Acid Research and Molecular Biology, 61, 1-23. doi: 10.1016/S0079-6603(08)60823-5 [25] Ali, H., Jurga, M., Kurgonaite, K., Forraz, N. and McGuckin, C. (2009) Defined serum-free culturing con- ditions for neural tissue engineering of human cord blood stem cells. Acta Neurobiologiae Experimentalis, 69, 11- 23. [26] Guzen, F.P., De Almeida Leme, R.J., De Andrade, M.S., de Luca, B.A. and Chadi, G. (2009) Glial cell line- de- rived neuro-trophic factor added to a sciatic nerve frag- ment grafted in a spinal cord gap ameliorates motor im- pairments in rats and increases local axonal growth. Restorative Neurology and Neuroscience, 27, 1-16. [27] Willner, P., Towell, A., Sampson, D., Sophokleous, S. and Muscat, R. (1987) Reduction of sucrose preference by chronic un- predictable mild stress, and its restoration by a tricyclic antidepressant. Psychopharmacology, 93, 358-364. doi:10.1007/BF00187257 [28] Moreau, J.L., Jenck, F., Martin, J.R., Mortas, P. and Hae- fely, W.E. (1992) Antidepressant treatment prevents chronic unpredictabl mild stress-induced anhedonia as assessed by ventral tegmentum self-stimulation behavior in rats. European Neuropsychopharmacology, 2, 43-49. doi:10.1016/0924-977X(92)90035-7 [29] Shi, Z.R., Itzkowitz, S.H. and Kim, Y.S. (1988) A com- parison of three immunoperoxidase techniques for anti- gen detection in colorectal carcinoma tissues. Journal of Histochemistry & Cytochemistry, 36, 317-322. doi: 10.1177/36.3.3278057 [30] Chen, J.Q., Zhan, W.H., He, Y.L., Peng, J.S., Wang, J.P., Cai, S.R. and Ma, J.P. (2004) Expression of heparanase gene, CD44v6, MMP-7 and nm23 protein and their rela- tionship with the invasion and metastasis of gastric car- cinomas. World Journal of Gastroenterology, 10, 776- 782. [31] Malberg, J.E., Eisch, A.J., Nestler, E.J. and Duman, R.S. (2000) Chronic antidepressant treatment increases neu- rogenesis in adult rat hippocampus. The Journal of Neu- roscience, 20, 9104-9110. [32] Parent, J., Yu, T., Leibowitz, R., Geschwind, D., Sloviter, R. and Lowenstein, D. (1977) Dentate granule cell neu- rogenesis is increased by seizures and contributes to ab- errant net-work reorganization in the adult rat hippo- campus. The Journal of Neuroscience, 17, 3727-3738. [33] Paxinos, G. and Watson, C. (1986) The rat brain in ste- reo-taxic coordinates. Academic, San Diego. [34] Ebendal, T. (1992) Function and evolution in the NG Ffamily and it sreceptors. Journal of Neuroscience Re- search, 32, 461-470. doi:10.1002/jnr.490320402 [35] Hefti, F., Knusel, B. and Lapchak, P.A. (1993) Protective effects of nerve growth factor and brain-derived neuro- trophic factor on basal forebrain cholinergic neurons in adult rats with partial fimbrial transactions. Progress in Brain Research, 98, 257-263. doi: 10.1016/S0079-6123(08)62407-3 [36] Backman, C., Rose, G.M., Hoffer, B.J., Henry, M.A., Bartus, R.T., Friden, P. and Granholm, A.C. (1996) Sys- temic administration of nerve growth factor conjugate reverses age-related cognitive dysfunction and prevents cholinergic neuron atrophy. The Journal of Neuroscience, 16, 5437-5442. [37] Isacson, O., Seo, H., Lin, L., Albeck, D. and Granholm, A.C. (2002) Alzheimer’s disease and Down’s syndrome: Roles of APP, tropic factors and Ach. Trends in Neuro- sciences, 25, 79-84. doi:10.1016/S0166-2236(02)02037-4 [38] Counts, S.E., Nadeem, M., Wuu, J., Ginsberg, S.D., Saragovi, H.U. and Mufson, E.J. (2004) Reduction of cortical Trk A but not p75 (NTR) protein in early-stage Alzheimer’s disease. Annals of Neurology, 56, 520-531. doi:10.1002/ana.20233 [39] Lu, B. (2003) Pro-region of neurotrophins: Role in syn- aptic modulation. Neuron, 39, 735-738. doi:10.1016/S0896-6273(03)00538-5 [40] Suzuki, S., Numakawa, T., Shimazu, K., Koshimizu, H., Hara, T., Hatanaka, H., Mei, L., Liu, B. and Kojima, M. (2004) BDNF-induced recruitment of Trk B receptor into neuronal lipid rafts: Roles in synaptic modulation. The Journal of Cell Biology, 167, 1205-1215. doi: 10.1083/jcb.200404106 [41] Smith, D.E., Robers, J., Gage, F.H. and Tusynski, M.H. (1999) Age-associated neuronal atrophy occurs in the primate brain and is reversible by growth factor gene therapy. Proceedings of the National Academy Sciences of the United States of America, 96, 10893-10898. doi:10.1073/pnas.96.19.10893 [42] Cotman, C.W. and Berchtold, N.C. (2002) Exercise: A behavioral intervention to enhance brain health and plas- ticity. Trends in Neurosciences, 25, 295-301. doi:10.1016/S0166-2236(02)02143-4 [43] Bibel, M. and Barde,Y.A. (2000) Neurotrophins: Key C opyright © 2011 SciRes. WJNS  L. J. Yu et al. / World Journal of Neuroscience 1 (2011) 9-18 Copyright © 2011 SciRes. 18 WJNS regulators of cell fate and cell shape in the vertebrate nervous system. Genes & Development, 14, 2919-2937. doi:10.1101/gad.841400 [44] Huang, E.J. and Reichardt, L.F. (2001) Neurotrophins: Roles in neuronal development and function. Annual Re- views of Neuroscience, 24, 677-736. doi: 10.1146/annurev.neuro.24.1.677 [45] Chao, M.V. (2003) Neurotrophins and their receptors: A convergene point for many signaling pathways. Nature Reviews Neuroscience, 4, 299-309. doi:10.1038/nrn1078 [46] Fischer, W., Wictorin, K., Bjorklund, A., Williams, L.R., Varon, S. and Gage, F.H. (1987) A melioration of cho- linergic neuron atrophy and spatial memory impairment in aged rats by nerve growth factor. Nature, 329, 65-68. doi:10.1038/329065a0 [47] Pizzo, D.P. and Thai, L.J. (2004) Intraparenchymal nerve growth factor improves behavioral deficits while mini- mizing the adverse effects of intracerebroventricular de- livery. Neuroscience, 124, 743-755. doi:10.1016/j.neuroscience.2003.12.041 [48] Jakubowska-Dogru, E. and Gumusbas, U. (2005) Chro- nic in-tracerebroventricular NGF administration improves working memory in young adult memory deficientrats. Neuroscience Letters, 382, 45-50. doi: 10.1016/j.neulet.2005.02.059 [49] Tuszynski, M.H., Thal, L., Pay, M., Salmon, D.P., Bakay, R., Patel, P., Blesch, A., Vahlsing, H.L., Ho, G., Tong, G., Potkin, S.G., Fallon, J., Hansen, L., Mufson, E.J., Kor- dower, J.H., Gall, C. and Conner, J. (2005) A phase I clinical trial of nerve growth factor gene therapy for Al- zheimer disease. Nature Medicine, 11, 551-555. doi:10.1038/nm1239 [50] Conner, J.M., Lauterborn, J.C., Yan, Q., Gall, C.M. and Varon, S. (1997) Distribution of brain-derived neuro- trophic factor (BDNF) protein and mRNA in the normal adult CNS: Evident for anterograde axonal transport. The Journal of Neuroscience, 17, 2295-2313. [51] Tolwani, R.J., Buckmaster, P.S., Varma, S., Cosgaya, J.M., Wu, Y., Suri, C. and Shooter, E.M. (2002) BDNF overexpression increases dendrite complexity in hippo- campal dentate gyrus. Neuroscience, 114, 795-805. doi:10.1016/S0306-4522(02)00301-9 [52] Korte, M., Carroll, P., Wolf, E., Brem, G., Thoenen, H. and Bonhoeffer, T. (1995) Hippocampal long-term poten- tiation is impaired in mice lacking brain-derived neuro- trophic factor. Proceedings of the National Academy Sciences of the United States of America, 92, 8856-8860. doi:10.1073/pnas.92.19.8856 [53] Linnarsson, S., Bjorklund, A. and Ernfors, P. (1997) Learning deficits in BDNF mutant mice. European Jour- nal of Neuroscience, 9, 2518-2587. doi: 10.1111/j.1460-9568.1997.tb01687.x [54] Kuhn, H.G., Dickinson-Anson, H. and Gage, F.H. (1996) Neuro-genesis in thedentate gyrus of the adult ratage-related decrease of neuronal progenitor prolifera- tion. The Journal of Neuroscience, 16, 2027-2033. [55] Kempermann, G., Kuhn, H.G. and Gage, F.H. (1997) More hip-pocampal neurons in adul tmice living in an enriched environment. Nature, 386, 493-495. doi: 10.1038/386493a0 [56] Gould, E., Beylin, A.,Tanapat, P., Reeves, A. and Shors, T.J. (1999) Learning enhances adult neurogenesis in the hippocam-pal formation. Nature Neuroscience, 2, 260- 265. doi.org/10.1038/6365 [57] Van Praag, H., Kempermann, G. and Gage, F. (1999) Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus. Nature Neuroscience, 2, 266-270. doi:10.1038/6368 [58] Gould, E., McEwen, B. S., Tanapat, P., Galea, L.A. and Fuchs, E. (1997) Neurogenesis in the dentate gyrus of the adult tree shrew is regulated by psychosocial stress and NMDA receptor activation. The Journal of Neuroscience, 17, 2492-2498. [59] Gould, E., Tanapat, P., McEwen, B.S., Flugge, G. and Fuchs, E. (1998) Proliferation of granule cell precursors in the dentate gyrus of adult monkeys is diminished by stress. Proceedings of the National Academy Sciences of the United States of the America, 95, 3168-3171. doi:10.1073/pnas.95.6.3168 [60] Tanapat, P., Galea, L.A. and Gould, E. (1998) Stress in- hibits the proliferation of granule cell precursors in the developing dentate gyrus. International Journal of De- velopmental Neuroscience, 16, 235-239. doi:0.1016/S0736-5748(98)00029-X [61] Fuchs, E., Flugge, G., McEwen, B.S., Tanapat, P., Gould, E. (1997) Chronicsubordination stress inhibits neuro- genesis and decreases the volume of the granule cell layer. Society Neurosci Abstr, 23, 317. [62] Kee, N., Sivalingam, S., Boonstra, R. and Wojtowicz, J.M. (2002) The utility of Ki-67 and BrdU as prolifera- tive markers of adult neurogenesis. Journal of Neurosci- ence Methods, 115, 97-105. doi:10.1016/S0165-0270(02)00007-9
|