Advances in Bioscience and Biotechnology
Vol. 3 No. 7 (2012) , Article ID: 24698 , 6 pages DOI:10.4236/abb.2012.37107
Tissue-specific isoenzyme variations in Arabian camel, Camelus dromedaries
![]()
1Department of Biology, Faculty of Science, Taif University, Taif, Saudi Arabia
2Department of Zoology, Faculty of Science, Cairo University, Giza, Egypt
Email: *HUyasser92us@yahoo.comU
Received 13 August 2012; revised 21 September 2012; accepted 25 October 2012
Keywords: Electrophoresis; Esterase; Malate Dehydrogenase; Tissue Specificity; Arabian Camel
ABSTRACT
Malate dehydrogenase, α-esterase and β-esterase isoenzymes have been analyzed by native polyacrylamide gel electrophoresis in different tissues (liver, kidney, muscle and hump) of the Arabian Camel, Camelus dromedaries in order to study the tissue specificity of these isoenzymes. Malate dehydrogenase recorded three fractions while both esterases recorded two fractions in different studied tissues. Malate dehydrogenase was considered as a key isoenzyme in discriminating among the different studied tissues. The percentage amount of both Mdh-3 and Mdh-2 showed significant difference among the different tissues while Mdh-1 recorded significant variation between muscle and hump tissues. The significant variation in the amount of Mdh isoenzyme (on the level of either total or fractions) among the different studied tissues may reflect the role of this enzyme for energy production in this desert animal. With respect to esterases, the percentage amount for both fractions and the total enzyme did not show significant difference among different tissues except β-Est-2 which was significantly higher in hump than in muscle. This enzyme showed higher thickness and intensity in both liver and kidney than in the muscle and hump indicating its greater activity in both liver and kidney.
1. INTRODUCTION
The dromedary (Camelus dromedarius), also called Arabian camel or one humped camel, was domesticated some 5000 years ago in the Arabian Peninsula. This species is able to survive in hot temperatures that is normally lethal to other species. All the functions of this species are seen to be adapted to desert environment which is characterized by little water and poor food [1]. The total population of dromedary is estimated to be around 1.6 million camels within the Arabian Peninsula constituting 51% of the total tropical livestock unit in Saudi Arabia. It is economically very important for milk and meat production which was mainly linked to the population growth [2].
The Arabian camel has paid the attention of many investigators on physiological [3-5], behavioral [6-8], genetical [9-11] and ecological [12,13] levels.
Isoenzymes are multiple forms of a single enzyme, which often have different isoelectric points and therefore can be separated by electrophoresis. Electrophoretic studies were done extensively on the different tissues of various animals from which it reveals that the enzyme exit in multi molecular forms and functions [14]. Several isoenzymes of malate dehydrogenase (EC 1.1.1.37) exist. There are two main isoforms in eukaryotic cells [15]. One is found in the mitochondrial matrix, participating as a key enzyme in the citric acid cycle that catalyzes the oxidation of malate. The other is found in the cytoplasm, assisting the malate-aspartate shuttle with exchanging reducing equivalents [16]. Esterase isoenzymes are one of the lipid-hydrolyzing enzymes, possess high significance in genetics and toxicology [14,17]. Esterases are also used as bioindicators to measure the toxic potency of pesticide residues usually applied in agriculture [18].
To the best of our knowledge, there is no study tackling the isoenzyme system and its tissue specificity in the Arabian camel so far. The present study therefore, aimed to investigate the patterns of variations of malate dehydrogenase and esterases (α and β) isoenzymes in different tissues of the Arabian camel that has been used for milk and meat production.
2. MATERIALS AND METHODS
2.1. Sample Preparation and Isoenzyme Assay
Tissue samples of liver, kidney, muscle and hump were freshly obtained from a slaughterhouse in Taif city, immediately taken to the lab and stored at −80˚C for further laboratory use.
For isoenzyme extraction, approximately 0.5 g of tissue was homogenized in 1 mL saline solution NaCl (0.9%) using a manual Homogenizer. The homogenates were centrifuged at 5000 rpm for 10 minutes and the supernatants were kept at −20˚C until use. For electrophoresis, 30 μL of the extract was mixed with 10 μL of treatment buffer and 35 μL of this mixture was applied to the well. Isoenzymes were electrophorased in 10% native-polyacrylamide gel as described by Stegemann et al. [19]. After electrophoresis, the gels were stained according to their enzyme system with the appropriate substrate and chemical solutions then incubated at room temperature in dark for complete staining. In most cases the incubation for about 1 to 2 hours is enough.
2.2. Malate Dehydrogenase (Mdh)
After electrophoresis, the gel was soaked in 100 mL of 0.05 M Tris HCl pH 8.5 containing 25 mg NBT, 25 mg EDTA, 25 mg NAD, 10 mg malic acid and 3 mg PMS [20]. 0.05 M Tris HCl pH 8.5 was prepared by dissolving 0.605 g Tris in 50 mL distilled water. The pH was adjusted to 8.5 by HCl. Then the solution was completed to 100 ml by distilled water.
2.3. α and β Esterase Enzymes (α-Est and β-Est)
After electrophoresis, the gel was soaked in 0.5 M borate buffer (pH 4.1) for 90 minutes at 4˚C. This procedure lowers the pH of the gel from 8.8 to about 7 at which the reaction proceeds readily. The low temperature minimizes diffusion of the protein within the gel. The gel then was rinsed rapidly in two changes of double distilled water. The gel was stained for esterase activity by incubation at 37˚C in a substrate solution of 100 mg α- naphthyl acetate (α-Est) and b-naphthyl acetate (b-Est) and 100 mg fast blue RR salt in 200 ml of 0.1 M phosphate buffer pH 6.5 [21].
2.4. Gel Fixation
After the appearance of the enzyme bands, the reaction was stopped by washing the gel two or three times with tap water. This was followed by adding the fixative solution, which consists of ethanol and 20% glacial acetic acid (9:11 v/v). The gel was kept in the fixative solution for 24 hours and then was photographed.
2.5. Statistics
All gels were scanned using Gel Doc-2001 Bio-Rad system. For isoenzymes, the bands of enzyme activity were designated using the known system of nomenclature [22]. An abbreviation which corresponds to the name of the enzyme designated each locus. When multiple loci were involved, the fastest anodal protein band was designated as locus one, the next as locus two and so on. Student t-test in the SPSS package v. 13 was used to calculate the significant differences of the percentage amounts for isoforms among the different studied tissues.
3. RESULTS AND DISCUSSION
3.1. Malate Dehydrogenase
Table 1 showed the mean and standard error for the per-
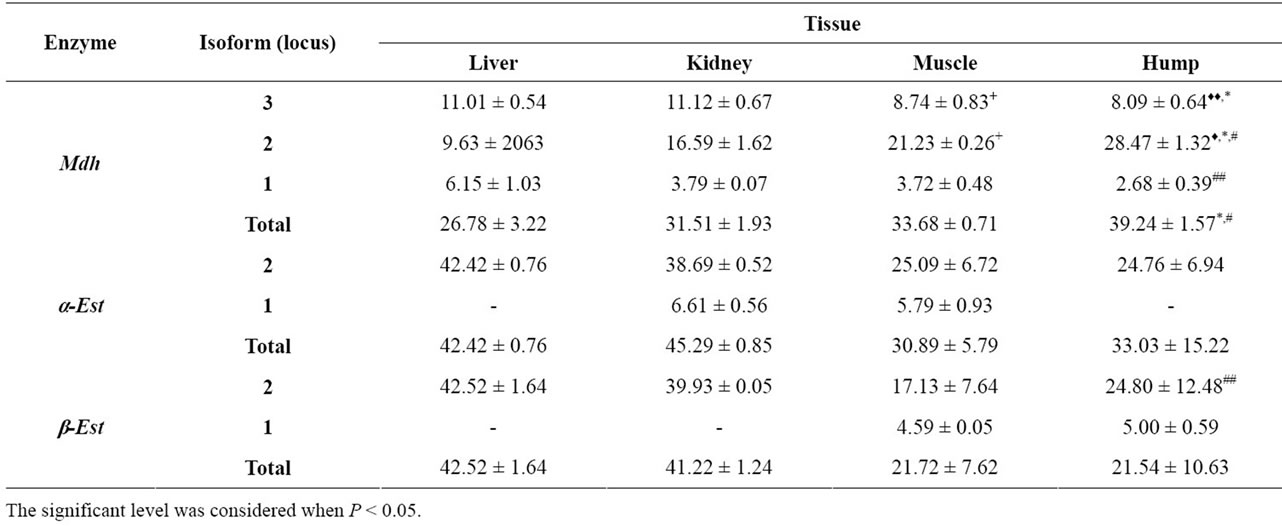
Table 1. Mean ± SE of the percentage amount for the studied isoenzymes in different tissues of Arabian Camel. The significance level was calculated by Student t-test. + = Significance level between liver and muscle, ♦ = Significance level between liver and hump, $ = Significance level between kidney and muscle, * = Significance level between kidney and hump and # = Significance level between muscle and hump.
centage amount of the studied isoenzymes in the different camel tissues. The three Mdh isoforms showed variations in their activities when the different tissues were compared. With respect to Mdh-3, the percentage amount was significantly higher in both liver (P < 0.01) and kidney (P < 0.05) than in the hump. It was 11.01 ± 0.54 in liver and 11.12 ± 0.67 in the kidney compared to 8.09 ± 0.64 in hump tissues. Kidney tissues showed significantly higher activity (P < 0.05, 11.12 ± 0.67) for this fraction compared to muscle tissue (8.74 ± 0.83). Conversely, Mdh-2 was significantly higher (P < 0.05) in hump tissues (28.47 ± 1.32) than in liver (9.63 ± 2063), kidney (16.59 ± 1.62) and muscle tissues (21.23 ± 0.26). Mdh-1 showed significant difference (P < 0.01) only when the muscle and hump tissues were compared being higher in muscle. When the total enzyme was examined, the percentage amount was significantly higher (P < 0.05) in hump tissue (39.24 ± 1.5) than in either kidney (31.51 ± 1.93) or muscle (33.68 ± 0.71) tissues.
Malate dehydrogenases constitute a family of multimeric enzymes that reversibly catalyzes the conversion of oxaloacetate and malate into pyruvate, and that differ in their subcellular localization and specificity for the cofactor. Human cytosolic malate dehydrogenase (socalled Mdh-1) is important in transporting NADH equivalents across the mitochondrial membrane, controlling tricarboxylic acid (TCA) cycle pool size and providing contractile function. Cellular localization studies indicate that Mdh-1 mRNA expression has a strong tissue-specific distribution [23]. The variations of MDH functions are primarily related to aerobic energy production for muscle contraction, absorption/resorption functions and processes related to gas exchange [24]. The significant increase in the amount of Mdh in liver, kidney and muscle tissues, in the present study, could be also an indication of similar functions.
The cytosolic MDHs catalyze the NADP dependent oxidative decarboxylation of malate into pyruvate and carbon dioxide to generate NADPH. Due to its ability to produce NADPH, this enzyme is thought to be a key enzyme in lipid biosynthesis [24]. We therefore can assume that the significant increase of Mdh-2 in hump tissues is for a similar purpose. The total enzyme activity was shown to be significantly higher in hump than in other tissues and this supports the role of the hump tissues in performing its function. Figure 1 illustrates the electrophoretic pattern of Mdh isoenzymes in different studied tissues. Mdh-2 was shown to be the highest expressed isoform and this may indicate the ability of the hump tissues in lipid biosynthesis.
3.2. Esterases
The mean and standard error values for the percentage amounts of both isoenzymes are shown in Table 1. Regarding the total isoenzyme activity, both α and β-esterase isoforms did not show any significant differences among the different studied tissues. The significance in the fractional activity was shown only for β-Est-2 which was higher (P < 0.01) in hump (24.80 ± 12.48) compared to muscle (17.13 ± 7.64). Comparison of α and β naphthyl acetate isoform patterns (Figures 2 and 3) among the different tissues revealed that, the second fraction (α-Est-2 and β-Est-2) was detected in all tissues examined showing high activity in both liver and kidney. Different levels of esterase band activity, denoted by different thickness and degree of staining were in of great enzymatic activity [25]. The present results therefore revealed the higher activity of esterases in liver and kidney than in muscle and hump may be indicative of the role of these organs in hydrolyzing the toxic substances that have been accumulated in the camel body.
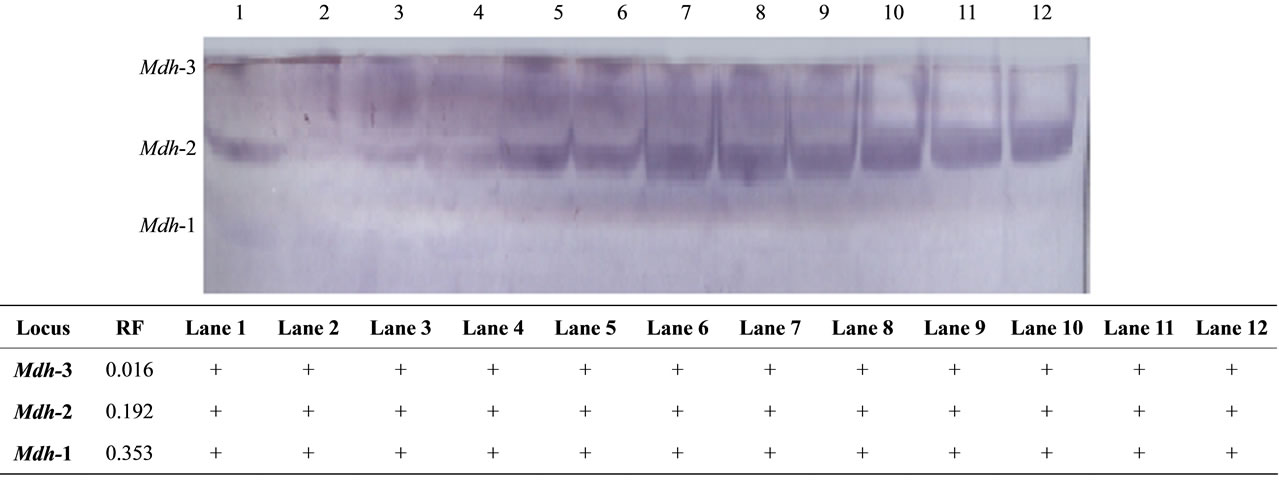
Figure 1. The electrophoretic profile (above) and the recorded isoforms with the relative mobility (RF) (below) of Mdh isoenzymes in the studied tissue samples. Lanes are as follow: 1 - 3 (liver), 4 - 6 (kidney), 7 - 9 (muscle) and 10 - 12 (hump).
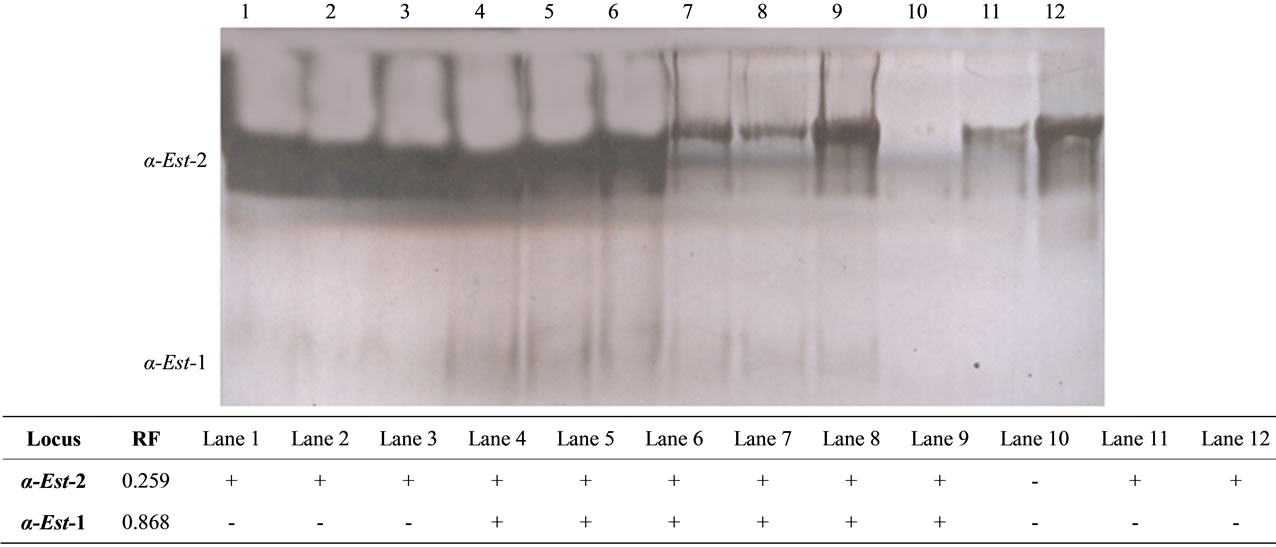
Figure 2. The electrophoretic profile (above) and the recorded isoforms with the relative mobility (RF) (below) of α esterase isoenzymes in the studied tissue samples. Lanes are as follow: 1 - 3 (liver), 4 - 6 (kidney), 7 - 9 (muscle) and 10 - 12 (hump).
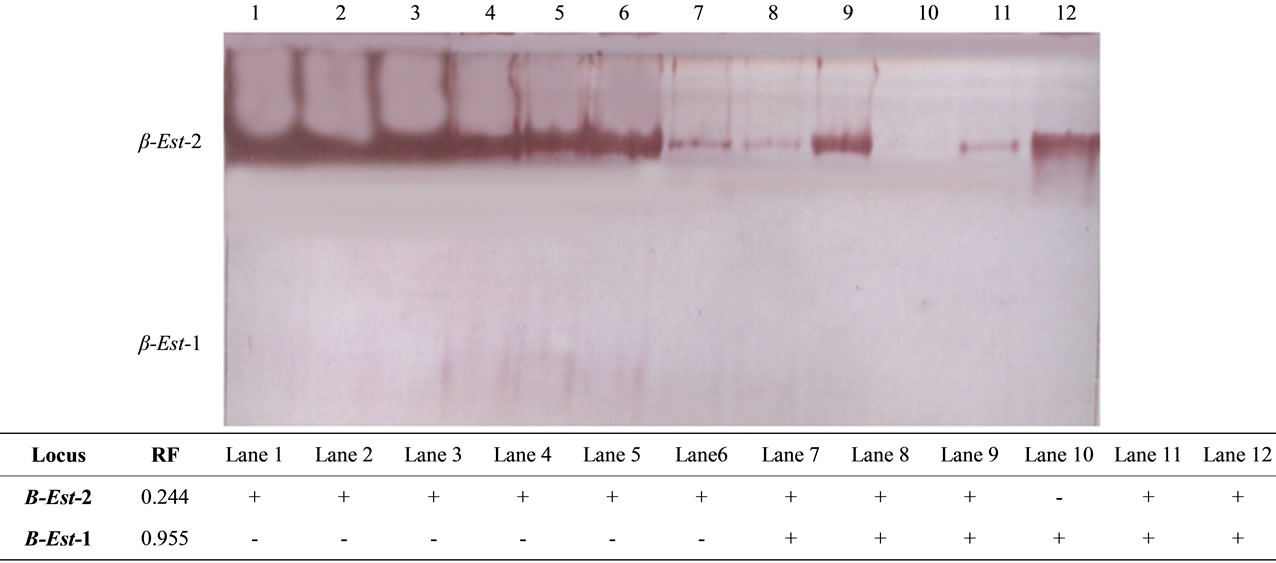
Figure 3. The electrophoretic profile (above) and the recorded isoforms with the relative mobility (RF) (below) of β esterase isoenzymes in the studied tissue samples. Lanes are as follow: 1 - 3 (liver), 4 - 6 (kidney), 7 - 9 (muscle) and 10 - 12 (hump).
On the other hand, the fastest anodal isoform of α esterase (α-Est-1) was detected in kidney and muscle tissues whereas, that of β esterase (β-Est-1) was detected in muscle and hump tissues. This result indicated that esterases were better applicable in revealing the isoenzymes tissue specificity as concluded by Shahjahan et al. [18]. As esterases are used as bioindicators to measure the toxic potency of pesticide residues usually applied in agriculture [18], the null variations in the activity of esterases in the studied tissues may indicate, to some extent, the safety of the diet applied to this animal in Saudi Arabia. We therefore consider that the productivity of Arabian camel for milk and meat in Saudi Arabia is a strate gic standpoint of the economy and is highly recommended.
The results presented herein are for tissues that have been subjected to multiple freeze-thaw conditions and this could affect the banding patterns and the enzyme activity [26]. Because all of our samples were subjected to the same conditions, this factor can be neglected.
4. CONCLUSION
In conclusion, the present study showed that organs and tissues within the animal body showed different behaviors in expressing the enzyme activity. This view can be used for discriminating among populations and species in systematic and physiology.
5. ACKNOWLEDGEMENTS
We are grateful to Dr. Shawkat Ahmed at Ain Shams University of Egypt for his technical support in conducting the practical part of this work.
![]()
![]()
REFERENCES
- Souilem, O. and Barhoumi, K. (2009) Physiological Particularities of Dromedary (Camelus dromedarius) and experimental implications. Scandinavian Journal of Laboratory Animal Science, 36, 19-29.
- Faye, B. and Bonnet, P. (2012) Camel sciences and economy in the world: Current situation and perspectives. In: Johnson, E.H., et al., Eds., 3rd Proceeding ISOCARD Conference, Muscat, Sultanate of Oman, 2-15.
- Azwai, S.M., Saltani, H., Thomas, P.C., Shareha, A.M., El Gammudi, F. and Mohamed, S.O. (1990) Note on cholesterol, glucose, urea and total protein concentration in serum of normal camels. Camel Newsletter, 7, 94.
- Al-Qarawi, A.A. and Ali, B.H. (2003) Variations in the normal activity of esterases in plasma and liver of Camels (Camelus dromedarius), Cattle (Bos indicus), Sheep (Ovis aries) and Goats (Capra hircus). Journal of Veterinary Medicine, 50, 201-203.
- Badryyah, A., Al-Suwaigh, R. and Al-Suhaimi A. (2005) Comparative study on some biochemical constituents of plasma in male camels and goats. Journal of Camel Practice and Research, 12, 141-143.
- Elkhawad, A.O. (1992) Selective brain cooling in desert animals: The Camel (Camelus dromedarius). Comparative Biochemistry and Physiology, 101, 195-202. HUdoi:10.1016/0300-9629(92)90522-RU
- Dereje, M. and Udén, P. (2005) The browsing dromedary camel. Behaviour, plant preference and quality of forage selected. Animal Feed Science and Technology, 121, 297- 308. HUdoi:10.1016/j.anifeedsci.2005.01.017U
- Qari, S.A. and AL-Harbi, M.S. (2011) Effect of seasonal temperatures on physiology and behavior of Arabian Camels (Camelus dromedarieus) in the Mid-Western Region of Saudi Arabia. Glob al Veterinaria, 6, 537-541.
- Ahmed, O.M., Ben Salem, F., Bedhiaf, S., Rekik, B. and Djemali, M. (2010) Genetic diversity in Tunisian dromedary (Camelus dromedarius) populations using microsatellite markers. Livestock Science, 132, 182-185. HUdoi:10.1016/j.livsci.2010.05.016U
- Abdallah, H.R. and Faye, B. (2012) Phenotypic classification of Saudi Arabian camel (Camelus dromedarius) by their body measurements. Emirates Journal Food Agriculture, 24, 272-280.
- Almathen, F., Mwaracharo, J. and Hanotte, O. (2012) Genetic diversity and relationships of indigenous Saudi Arabia camel Camelus dromedarius populations. In: Johnson, E.H. et al., Eds., Third Proceeding ISOCARD Conference, Muscat, Sultanate of Oman, 40-41.
- Abbas, B., Al-Qarawi, A.A. and Al-Hawas, A. (2000) Survey on camel husbandry in Qassim region, Saudi Arabia: Herding strategies, productivity and mortality. Revue d’Elevage et de Médecine Vétérinaire des Pays Tropicaux, 53, 293-298.
- Shorepy, S.A. (2011) Identification of environmental factors affecting the racing performance of race camels in the United Arab Emirates. Emirates Journal of Food Agriculture, 23, 424-430.
- Markert, C.L. and Moller, F. (1959) Multiple forms of enzymes tissue, ontogenetic and species specific pattern. Proceeding of the Natural Academy of Sciences, 45, 753- 763. HUdoi:10.1073/pnas.45.5.753U
- Minarik, P., Tomaskova, N., Kollarova, M. and Antalik, M. (2002) Malate dehydrogenases—Structure and function. General Physiology and Biophysics, 21, 257-265.
- Musrati, R.A., Kollarova, M., Mernik, N. and Mikulasova, D. (1998) Malate dehydrogenase: Distribution, function and properties. General Physiology and Biophysics, 17, 193-210.
- Callaghan, A., Boiroux, V., Raymofld, M. and Pasteur, N. (1994) Prevention of changes in electrophoretic mobility of overproduced esterase from organophosphate-resistant mosquitoes of the Culex pipiens complex. Medical Veterinary and Entomology, 8, 391-394. HUdoi:10.1111/j.1365-2915.1994.tb00106.xU
- Shahjahan, R.M., Karim, A., Begum, R.A., Alam, M.S. and Begum, A. (2008) Tissue specific esterase isozyme banding pattern in Nile Tilapia (Oreochromis niloticus). University Journal of Zoology (Rajshahi University), 27, 1-5.
- Stegemann, H., Afify, A.M.R. and Hussein, K.R.F. (1985) Cultivar identification of dates (Phoenix dactylifera) by protein patterns. Second International Symposium of Biochemical Approaches to Identification of Cultivars, Brounschweig, Germany, 44.
- Jonathan, F.W. and Wendel, N.F. (1990) Visualization and interpretation of plant isozymes. In: Soltis, D.E. and Soltis, P.S., Eds., Isozymes in plant biology, Champan and Hall, London, 5-45.
- Scandaliojs, G. (1964) Tissue-specific isozyme variations in maize. Journal of Heredity, 55, 281-285.
- Allendorff, W. and Utter, F.M. (1978) Population genetics. In: Hoaran, W.S. and Randalal, D.J., Eds., Fish physiology 8, Academic Press, New York, 407-454.
- Lo, A.S.-Y., Liew, C.-T., Ngai, S.-M., Tsui, S.K.-W., Fung, K.-P., Lee, C.-Y. and Waye, M.M.-Y. (2005) Developmental regulation and cellular distribution of human cytosolic malate dehydrogenase (MDH1). Journal of Cell Biochemistry, 94, 763-773. HUdoi:10.1002/jcb.20343U
- Daniš, P. and Farkas, R. (2009) Hormone-dependant and hormone-independant control of metabolic and developmental functionals of malate dehydrogenase. Endocrine Regulations, 43, 39-52.
- Ahasan, M., Rahman, A., Hossain, S., Mazid, S.Z.S. and Hossain, A. (2011) Beta substrate in esterase enzyme of mosquito fish, Gambusia affinis. Bangladesh Pharmaceutical Journal, 14, 53-59.
- Cowan, K.J. and Storey, K.B. (2001) Tyrosine kinases and phosphatases in the estivating spadefoot toad. Cellular Physiology and Biochemistry, 11, 161-172. HUdoi:10.1159/000047803U
NOTES
*Corresponding author.