Open Journal of Ecology
Vol.2 No.3(2012), Article ID:21609,12 pages DOI:10.4236/oje.2012.23019
Spatial pattern of landscape changes and consequence changes in species diversity between 1956-1999 of Pinus halepensis Miller plantations in Montes de Malaga State Park (Andalusia, Spain)
![]()
1Department of Forestry Engineering-, University of Cordoba. Edf. Leonardo da Vinci, Cordoba-Spain, Andalusia, Spain;; *Corresponding Author: irlnacer@uco.es
2Institute Department of Forestry Engineering-University of Cordoba. Edf. Leonardo da Vinci, Cordoba-Spain
3Department of Forestry Engineering-University of Cordoba. Edf. Leonardo da Vinci, Cordoba-Spain
4Department of Forestry Engineering-University of Cordoba. Edf. Leonardo da Vinci, Cordoba-Spain
Received 15 March 2012; revised 8 April 2012; accepted 29 April 2012
Keywords: Diversity; Fragmentation; Landscape Metrics; Vegetation Cover; Mediterranean Ecosystem
ABSTRACT
Changes in forest diversity and the transformation of coniferous plantations into more natural woodlands are of special interest in the Mediterranean. The aim of this study was to investigate changes in land use patterns and vegetation cover from 1956 to 1999 in Pinus halepensis Miller plantations in Montes de Málaga State Park in southern Spain. At the landscape level, a FRAGSTAT analysis was performed using aerial photographs. At the community level, a total of 47 sample plots were analyzed. Field measurements indicated the presence of a total of 70 species in the different vegetation types sampled. Shannon-Wiener (H), and Simpson’s (D) diversity indices showed the greatest species richness in Aleppo pine plantations (H = 1.54 ± 0.2; D = 3.88 ± 0.6), while Holm oak stands in the forested areas (H = 1.37 ± 0.1; D = 3.35 ± 0.6) and agricultural land (H = 1.33 ± 0.01; D = 2.80 ± 0.03) had lower values. FRAGSTATS analyses at landscape and community levels indicated increases in patch richness, patch number, and percent of landscape mainly for pine plantations, Holm oak forests and shrub habitats. Replacement of agriculture by conifer plantations led to the homogenization of landscape structure. This change has modified the landscape by facilitating the recovery of indigenous tree species including Holm oak, which will likely lead to the future establishment of native forests in this area. Our findings suggest that pine plantations could favour woody species diversity by increasing abundance and species richness in contrast to the situation in non-forested agricultural land.
1. INTRODUCTION
As unique ecosystems with high biodiversity, Mediterranean forests have been identified as a biological hotspot [1,2]. The so-called anthropogenic agro-forest landscape in the region [3,4] consists of natural and semi-natural systems such as shrub lands, pastures, and woodlands interspersed with agricultural fields and a large number of pine plantations [5]. Large areas of Spanish landscapes were covered by different agricultural uses in the latter half of the 20th century. This land was converted to forest plantations mainly with the aim of preventing soil erosion, but little attention was paid to the effect on biodiversity. One species, Aleppo pine (Pinus halepensis Miller) dominates 43 percent of reforested areas in Spain [6].
Silvicultural methods can play a vital role in the restoration, protection and conservation of forest ecosystems [7,8]. The role of forest plantations in maintaining biodiversity is complex [9-11]. However, there has been an ongoing effort to manage coniferous plantations to ensure the transformation of the landscape to more diverse broadleaf forests [12-14]. Different conversion methods have been proposed including natural regeneration, which has been shown to be a better alternative than the restoration of forest plantations [7]. Natural regeneration in coniferous plantations depends on many factors, both abiotic (light availability, microclimate and soil conditions) and biotic (sources of diaspores, seed dispersal agents, competition and browsing). The most common problems in extensive plantations are the lack of seed trees of indigenous species, and low light due to a dense canopy [13-15]. However, canopy cover reduces the growth of herbaceous vegetation and, consequently, its competition with tree seedlings [16]. Studies of species diversity in plantations have mainly compared richness to that in natural forests, which are characterized by high diversity [10,13,17,18,10,13]. However, there has been relatively little research comparing diversity in plantations with that in agricultural land existing prior to forest plantations [19,20].
Originally, plantations in Spain were mostly established on agricultural land. An assessment of how the native ecosystem reacts to silvicultural management resulting from pine plantations is a crucial research gap for policy-makers. This study analyzes the changes in landscape patterns and vegetation cover from 1956 to 1999 in the Guadalmedina river basin in Southern Spain. We address the following questions: (i1) what was the role of Pinus halepensis plantations in the dynamic processes associated with the modification of the Mediterranean landscape? (ii2) How has plant diversity changed at the community level with regard to plantation activities?
2. METHODS
2.1. Study Area
The study was conducted in the forested area of Montes of Malaga State Park (Andalusia) and adjacent agricultural areas (Guadalmedina watershed), in southern Spain. The Montes de Málaga State Park (36˚49′´33′′´´N, 4˚32′´00′′´´W) occupies 4,995 ha; its elevation ranges from 90 to 1031 m a.s.l.. . Calcarenite, clay and sandstone (leptisols and luvisols) are the main soil types [21]. The maritime climate is mesic-Mediterranean and sub-humid maritime [22], characterised by mean annual temperatures between 14˚C and 17.5˚C, with extreme values ranging from 3˚C to 30˚C. Mean annual precipitation rangesmaritime climate is mesic-Mediterranean and sub-humid maritime [22], characterised by mean annual temperatures between 14 and 17.5˚C, with extreme values ranging from 3 to 30˚C. Mean annual precipitation ranges between 500 - 700 mm per year. Land use has changed considerably in the agricultural area surrounding the reserve, particularly since the 1930s, due to a reforestation programme conducted by the National Forestry Department and the decrease in traditional agricultural practices (e.g. terracing, grazing, and cultivation of olive and almond groves, and other annual crops).
Habitats with the following associated vegetation currently found in the reserve include: semi-natural grasslands (Brachypodium retusum (Pers.) Beauv, Dactylis glomerata L.); shrublands (Ule munity level analysis (Table 1) based on a review of previous studies [526,27,5]. x parviflorus Pourret, Rosmarinus officinalis L., Crataegus monogyna Jacq.); woodlands including Holm oak forests (Quercus ilex L.) and mixed evergreen woodlands (dominated by Q. ilex, Q. suber L. and Olea europaea L. subsp. sylvestris Brot.), riparian woody vegetation (dominated by Populus alba L., Ulmus minor, Arbutus unedo L. and Salix sp.) and olive and almond orchards. Native conifers (Pinus halepensis Mill.), which reproduce spontaneously, were planted from the 1930s to the 1970s. The current tree cover of Montes de Málaga State Park consists of pure Aleppo pine forest (60%), mixed Aleppo pine and oak species (16.5%), mixed shrub and pasture land (15%), and crops with little or no vegetation (2.3%).
Using aerial photos and forest inventory data, we confirmed that the previous land uses in the forested area of Montes de Málaga State Park had been agricultural. The study area included Montes de Málaga State Park and a surrounding agricultural land adding up to a total area of 13,436 ha (Figure 1).

Table 1. Landscape metrics used in this study and their brief definitions.
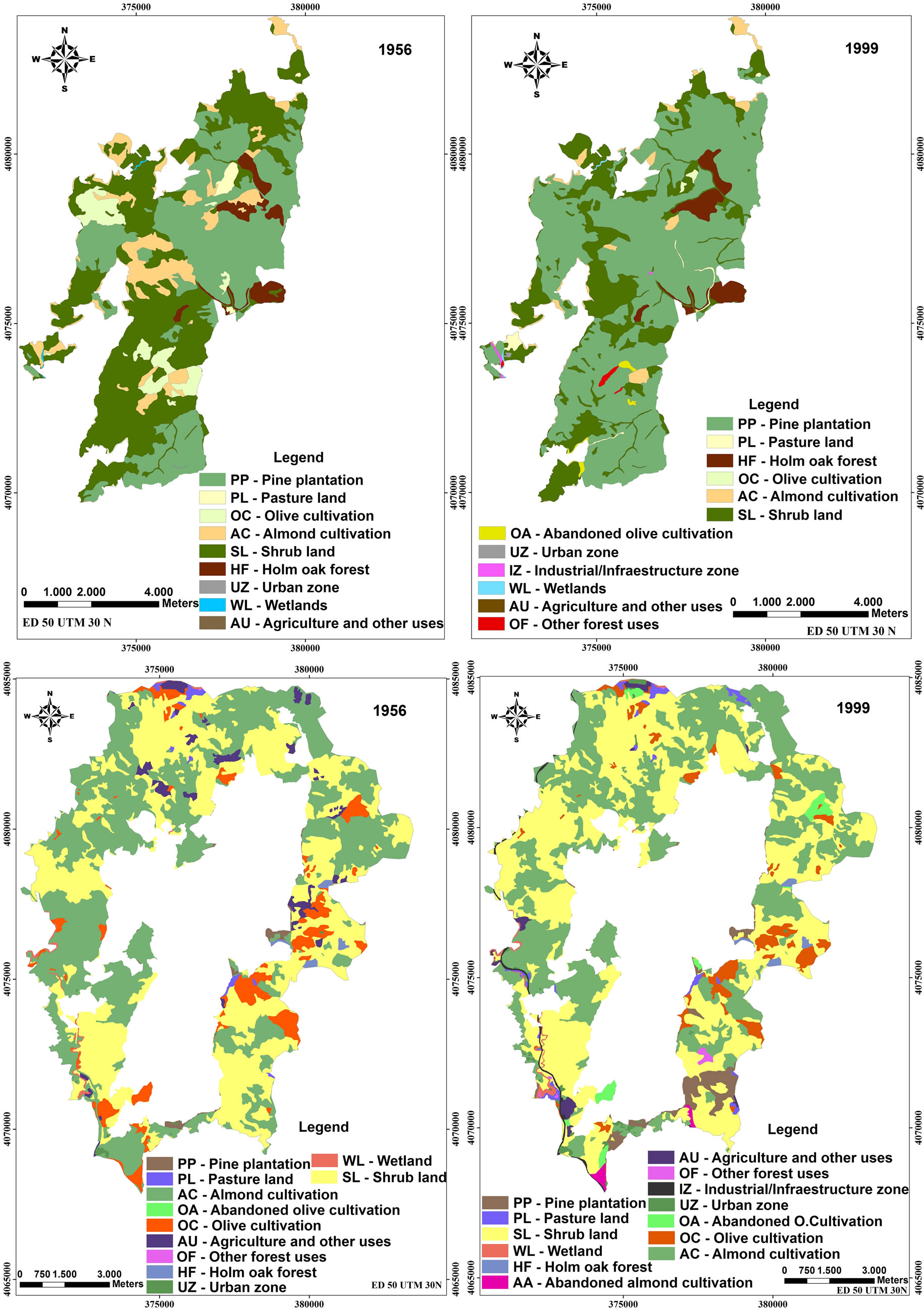
Figure 1. Changes in land use and vegetation cover from 1956 to 1999 of Montes de Málaga State Park and in the area surrounding the reserve (Andalusia, southern Spain).
2.2. Previous Forest Management
Aleppo pine forest was managed for conversion to even-aged stands dating back to 1970. The silvicultural operations in the landscape management scheme for Montes de Málaga State Park were generally designed to induce artificial successional changes in favour of the least frequently existing stand structures, such as Holm oak forest. Silvicultural treatments include regeneration cuts, thinning, pest protection[U1] and fire prevention[U2] treatments, and regimes for diverse and special conditions, for example afforestation, pest control, and others.
Thinning prescriptions include selective removals of 35% - 50% of suppressed (overtopped) trees. Marking was directed to harvest the smallest, slowest growing, damaged, or poorly formed trees with the highest risk of mortality. Thinning interventions carefully avoided opening the upper canopy. Steep, rocky stands and those in xeric sites are treated with selection cuts. Density and composition is controlled with this treatment, while group selection helps provide sites where regeneration can be expected.
Regeneration treatments aim at returning a stand to a more complex and diverse forest. Seed-tree treatments are the most common procedures, sometimes leaving around 30 - 50 seed trees per hectare, with up to 100 trees per hectare where conditions require extra protecprotecttion. Regeneration is also prescribed for stands recently affected by fire, pests or other similar disturbances resulting in severe mortality in which most of the trees that survived the disturbance event are retained as seed trees and harvesting is limited to those areas where the density of surviving trees is excessive.[U3]
2.3. Land Use Maps
Ortho aerial photographs taken in 1956 (grey-scale) and 1999 (colour; scale: 1:33,000 and 1:10.,000 respectively) were used (European datum 1950 Universal Transverse Mercator Huso 30). Each orthophoto was subjected to photo-interpretation [23] by the same interpreter to minimize errors. This procedure was performed for each of the two periods (1956 and 1999), thus, each spatial unit represents a plant community, whose identity may change over time. A total of 50 landscape and vegetation types existing there between 1956 and 1999 were merged into 15 representative landscape types, namely: 1) Pine (P. halepensis Miller) plantation (PP); 2) Pasture (PL); 3) Abandoned Olive tree (Olea europea L.) cultivation (OA); 4) Olive tree cultivation (129 OC); 5) Almond tree (Prunus dulcis [Mill.] D. A. Webb.) cultivation (AC); 6) Abandoned Almond tree cultivation (AA); 7) Shrub (SL); 8) Cereals (GW); 9) Other cultivation (CC); 10) Holm oak (Q. ilex L.) forest (HF); 11) Urban zone (UZ); 12) Industrial/infrastructure zone (IZ); 13) Wetlands (WL); 14) Other Agricultural land uses (AU); and 15) Other Forest land use (OF).
2.4. Field Sampling
To identify and verify changes in vegetation and land use types, we conducted a field inventory in the agricultural area surrounding the reserve and in the Montes de Málaga State Park itself. Data were collected during the months of July and August 2006 from a total of 47 sample plots representative of the following six main vegetation/land use types: pine plantation (PP) (N = 5); Holm oak forest (HF) (N = 6); shrub (SL) (N = 15); abandoned Olive and almond tree cultivation (OA/AA) (N = 10); olive and almond tree cultivation (AC) (N = 5); and pasture (PL) (N = 6) (Figure 1). The central point of each sample plot was referenced with a GPS hand-held unit to ensure accuracy and provide further post-analyses data. We randomly distributed the plots so that the total number of plots per stand varied between 5 and 15, depending on spatial distribution. Each plot was defined as a circular plot with a radius of 15 m (area = 706.96 m2). Variables recorded included abundance of tree, shrub and herbaceous species. The tree parameters measured in each plot included: diameter at breast height (DBH, measured at 1.30 m above the ground), number of trees per hectare (NTH); tree height (TH) (m) obtained with a hypsometer; and crown coverage (CC) (m2·ha), which was measured as the projection of the crown on the ground at the four cardinal points, using a tape measure from the base of the tree. Number of individuals, average height (m) and vegetation percent cover of understory shrubs were also measured. Finally, presence and percent cover of herbaceous species were recorded within a 2 × 15- m belt transect centered on the plot [24]. Richness (S, in number of species), Shannon-Wiener, and Simpson’s diversity indices were calculated based on the number of plant species encountered.
2.5. Landscape Analysis
FRAGSTATS, a program developed by the Forest Science Department, Oregon State University, USA, and widely used for quantifying landscape structure, was used to calculate various landscape metrics [25]. The program produces a number of metrics including area, patch density, size and variability, edge, shape, core area, and diversity. Although it is capable of computing more than 100 spatial statistics for quantifying landscape structure and spatial heterogeneity based on landscape composition and configuration, many of the variables are usually highly correlated and can be reduced using multivariate statistics [25]. In this study, we selected 10 metrics for landscape level analysis and 6 metrics for community level analysis (Table 1) based on a review of previous studies [5,26,27,5].
The metrics chosen were used to characterize spatial heterogeneity, fragmentation, complexity of patch shape, and connectivity for a given landscape. Brief definitions of the selected metrics are provided in Table 1 and detailed definitions are given in the FRAGSTATS User’s guide [25]. Number of patches (NP) and mean patch size (MPS) provided an indication of fragmented landscape condition [28]. The GIS platform used for visualization and spatial analysis was ArcGIS 9.2. From an ecological viewpoint, the combined use of biodiversity indices with FRAGSTATS spatial analyses facilitated the comparison of landscape change in forested areas of Montes de Málaga State Park and surrounding agricultural area.
2.6. Statistical Analyses
Normality of frequency distributions was tested by the Kolmogorov-Smirnov test. Richness (S), Shannon-Wiener (H), and Simpson’s (D) diversity indices of the tree and understory woody species were calculated for each plot [29]. Student’s t-tests were performed to examine differences between the richness and diversity indices between forested areas of Montes de Málaga State Park and surrounding agricultural area (two independent levels) according to vegetation types.
3. Results
3.1. Vegetation and Land Use Change
The maps depicting land use and vegetation types show the structure, land use distribution, and evolution of the landscape from 1956 to 1999 in forested areas of Montes de Málaga State Park and in the agricultural area surrounding the reserve (Figure 1). The most significant change observed within the park was that approximately 67% of the landscape was converted to pine plantations (P. halepensis) compared to 8% in the agricultural area. It is also important to note that Holm oak forest increasedin area by about 21 ha (0.42 % of the total area of the park). These changes were mainly due to the conversion of olive tree (O. europea) and almond tree (P. dulcis) orchards into pine plantations, Holm oak forests, and other forest uses (Table 2).
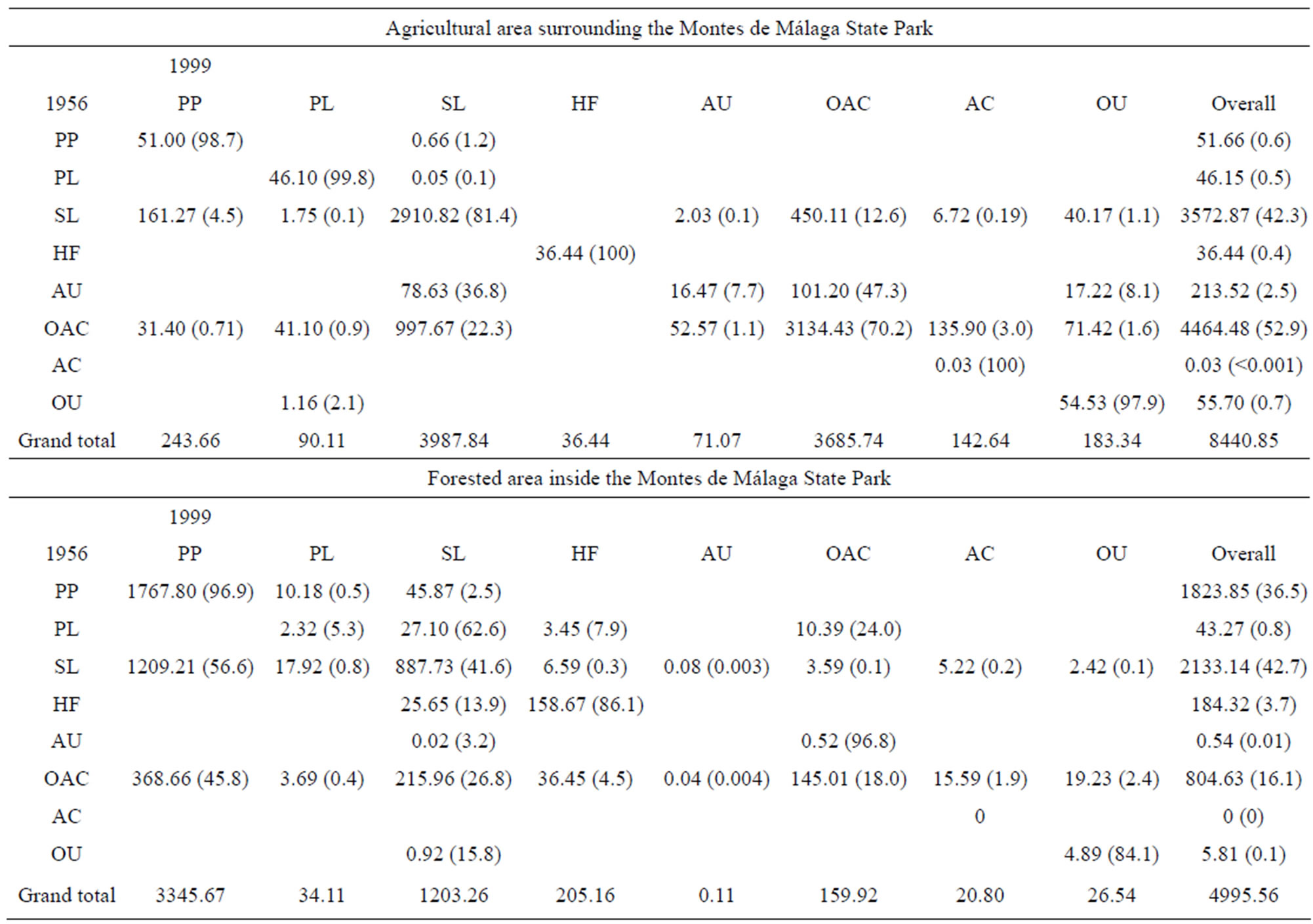
Table 2. The transition matrix of the surface of landscape patch types in the agricultural area surrounding and inside the Montes de Málaga State Park from 1956 to 1999. Values are area in hectares, with percent of total area in parentheses. PP: Pine plantation; PL: Pasture land; SL: Shrub land; HF: Holm oak forest; AU-: Agricultural and other uses; OAC: Olive and almond cultivation; AC: Abandoned olive and almond cultivation; OU: Other uses.
In the agricultural area around the park, 2362.7 hectares or 28% of the entire area underwent changes. One of the most important processes that occurred in the region during this period was the large-scale cessation of agricultural activity, which led to an increase in shrub cover and secondary shrubby growth. This vegetation type represented 22% of the area covered by woody species in 1956. Olive trees continued to form an important part of the canopy of secondary growth whereas almond trees died shortly agricultural abandonment.
3.2. Species Richness and Diversity
Plant diversity in the agricultural area around the reserve was lower than in the forested area within the reserve. A total of 56 woody species were recorded in the canopy and understory vegetation of both areas, with 31 species occurring in forested areas and 25 species in agricultural land, while 23 species were encountered in both areas. Mean species richness was significantly higher in forest than in abandoned olive and almond plantations (P = 0.027: paired T-test), Holm oak forests (P < 0.001), and pasture (P < 0.001) (Table 3). Similar to mean species richness, diversity indices were also significantly higher in plantation forests than in other habitats (Table 3). In the agricultural area, mean richness (S)
was highest in Holm oak forest (HF = 8.5 ± 0.05), shrub (SL = 7.2 ± 0.7), and abandoned olive and almond tree plantations (AC = 4.6 ± 1.5), while active olive and almond tree plantations (OAC = 1.2 ± 0.4) had the lowest values (Table 3). Despite higher species richness (S), the Shannon-Wiener diversity index indicated low evenness and high variation among all vegetation types ranging from 1.33 for Holm oak forest (HF) to 0.06 for olive and almond tree cultivation (OAC). Simpson’s diversity index (D) was higher in Holm oak (HF = 2.8 ± 0.03) and shrub (SL = 2.7 ± 0.2) than olive and almond tree plantations (OAC = 1.0 ± 0.04) and pasture (PL = 1.32 ± 0.74).
In contrast, in forested areas. , Species richness (S) was highest in pine plantations (PP = 10.8 ± 1.8), followed by pasture (PL = 9.5 ± 2.5), shrub land (SL = 8.2 ± 1.2), and abandoned olive tree plantations (AC = 7 ± 1.0) (Table 3). The Shannon-Wiener index in forested areas varied less than that of agricultural areas with only slight variation among all vegetation types ranging from 1.5 in pine plantations (PP) and 1.2 in Holm oak forest (HF) to 1.06 in shrub land (SL). Simpson’s diversity index (D) indicated lower diversity for shrub land (SL = 2.3 ± 0.2), and pasture (PL = 2.7 ± 0.6) and higher diversity for pine plantations (PP = 3.8 ± 0.6), abandoned olive and almond tree cultivation (AC = 3.3 ± 0.6) and Holm oak forest (HF = 2.9 ± 0.6).
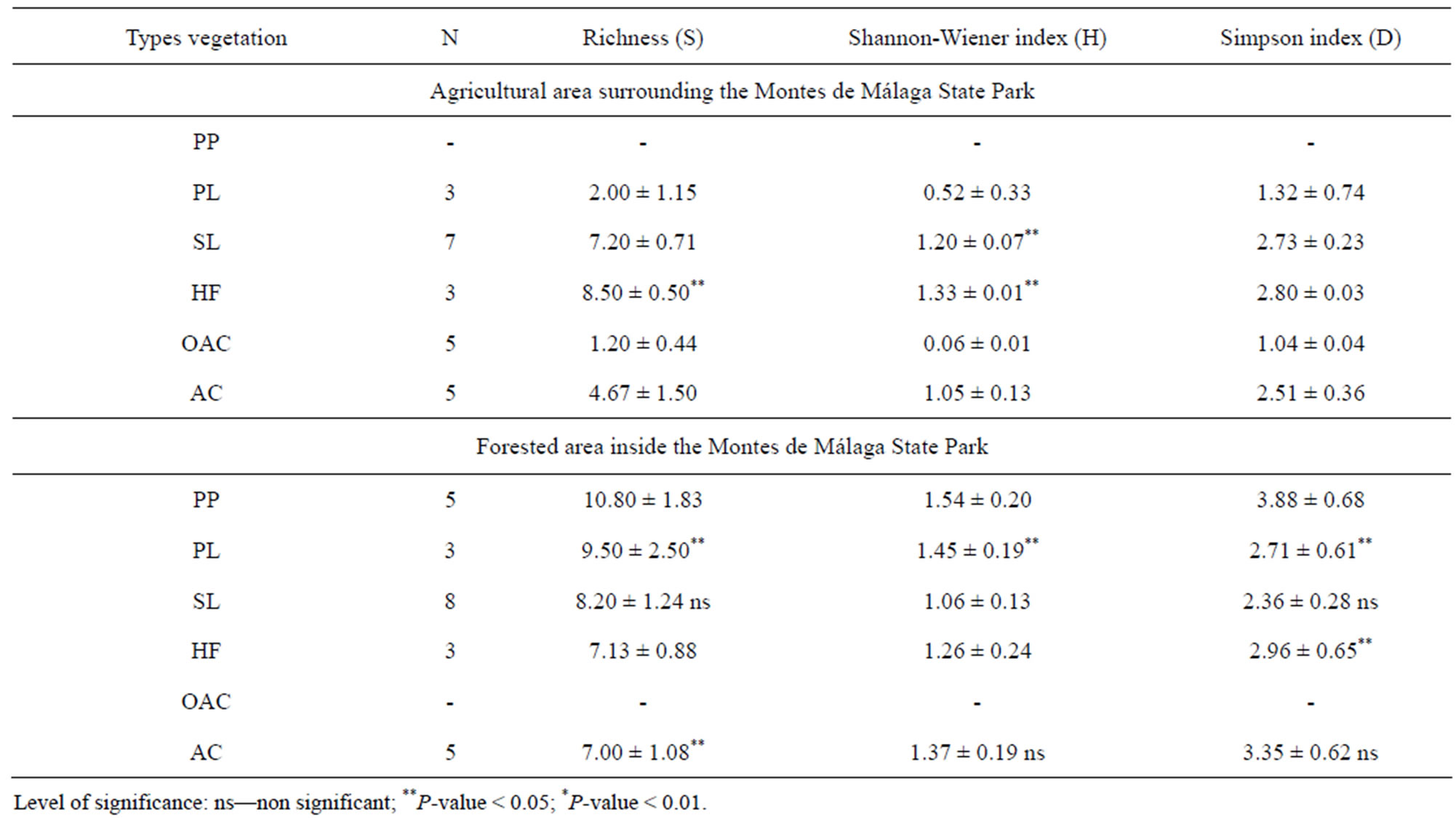
Table 3. Mean values of richness (S, in number of species), Shannon-Wiener, and Simpson’s diversity indices calculated for tree and woody understory species in the six types of sampled land use/vegetation types (PP: Pine plantation; PL: Pasture land; SL: Shrub land; HF: Holm oak forest; AU-: Agricultural and other uses; OAC: Olive and almond cultivation; AC: Abandoned olive and almond cultivation) in the agricultural area surrounding the reserve and forested area inside the Montes of Malaga State Park (Andalusia, southern Spain).
3.3. Changes in Landscape Metrics at Landscape Level
A comparison of changes in landscape metrics between 1956 and 1999 in both forested and agricultural areas at the landscape level is outlined below (Table 4).
The number of different vegetation types present within the forested area increased from 9 in 1956 to 12 in 1999. This change was paralleled by an increase in the number of patches from 274 in 1956 to 325 in 1999, and a decrease in mean patch area by about 3 231 ha. In addition, largest patch index (LPI) values showed a large increase from 21.19% to 52.8% indicating that the percentage of the landscape covered by the largest patch almost doubled. Similarly, the area-weighted mean index indicated a correlation between increases in the largest patch as the shape size changed from 3.51 to 5.66.
The decrease in the interspersion and juxtaposition indices (IJI) and increase in contagion indicated that the different patch types became less interspersed as the number of small dispersed patches declined resulting in decreased fragmentation of the landscape.
However, the extent of landscape interspersion does not necessarily imply that patch types were dispersed. It is also important to note that IJI and Contagion index (CONTAG) were inversely related.
Any indication of a decline in landscape heterogeneity was mainly due to the significant increase in pine plantations (PP). Shannon’s diversity and evenness indices, both of which decreased, also suggest that the landscapes in the study area became less fragmented and more homogeneous.
In the agricultural area surrounding the park, the number of patches increased from 478 in 1956 to 555 in 1999 leading to a decline in the relative area index (AREA_MN) from 17.6 to 15.2 in the same period. The rise in IJI from 44.9 in 1956 to 50.5 in 1999 and the decline in CONTAG from 74.0 to 72.5 indicates that the 1999 landscape was more fragmented and more heterogeneous than in 1956. Diversity indices increased in 1999, as well as the number of different land uses, rising from 11 in 1956 to 13 in 1999.
3.4. Changes in Landscape Metrics at Land Use Type
At the class level, the most significant change in Montes de Málaga State Park was the decrease in percent cover of almond and olive plantations, as well as shrub land (Table 5). Between 1956 and 1999, almond plantations, olive plantations, and shrub land declined from 11% to 3%, 5% to less than 1%, and 43% to 24%, respectively. In contrast, the area occupied by pine plantations increased from 37% to 67%. In 1956, secondary growth dominated most of the park, occupying 2130.9 ha or 43% of the landscape (PLAND). By 1999, this habitat had diminished as a result of conversion to other land uses such as pine forest, abandoned olive groves, Industrial and commercial zones and other forestry uses.
Pine plantations were the only land use type that increased significantly in area during the study period, and cover indices indicated a rapid rate of change. However, the number of patches of pine plantations decreased from 15 in 1956 to 12 in 1999. A similar pattern occurred for patches of Holm oak forest, which decreased from 8 to 6. By comparison, 14 patch types that did not exist in 1956 appeared in the study area in 1999, including abandoned olive plantations (7 patches), other forest uses (3 patches), and industrial/infrastructure zone (4 patches). Correspondingly, the mean patch area increased from 122 ha to 279 ha for pine plantations and from 23 ha to 34 ha for Holm oak forest.
When comparing all land use types with the analogous area-weighted mean shape index, pine plantations showed the greatest change from less than 4 ha in 1956 to about 7 ha in 1999. The increased patch number and decreased mean patch area of shrub and pasture suggest that these two land use types became more fragmented.
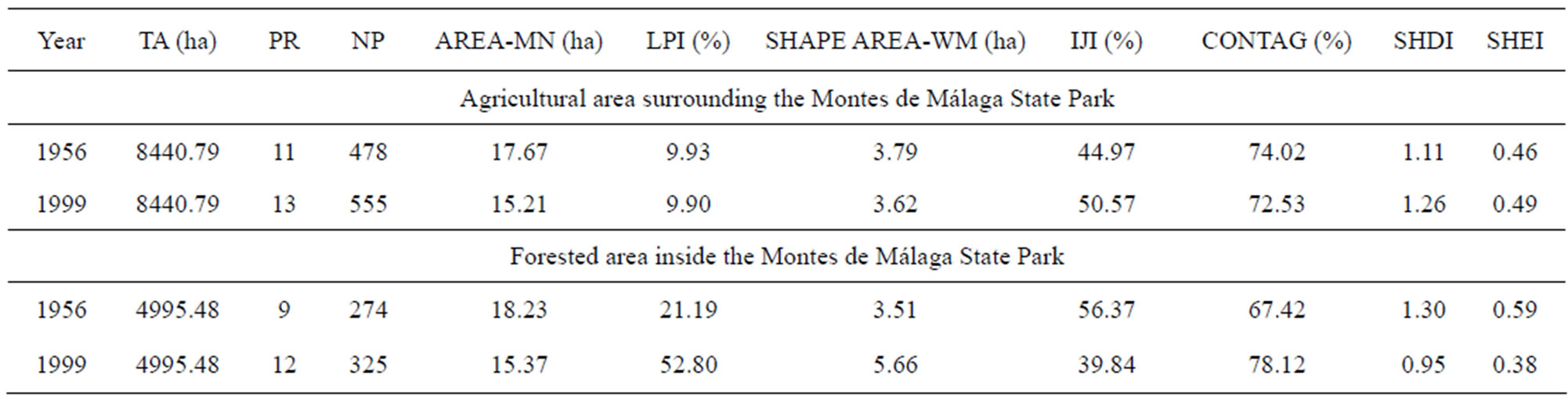
Table 4. Landscape metrics of 1956 and 1999 periods in the agricultural area surrounding the reserve and forested areas inside Montes de Málaga State Park (Andalusia, southern Spain). TA (ha): total area of landscape; PR: patch richness (number of patches/100 ha); NP: number of patches; AREA-MN (ha): mean patch area; LPI (%): largest patch index; SHAPE AREA-WM (ha): area-weighted mean shape index; IJI (%): interspersion and juxtaposition index; CONTAG (%): contagion index; SHDI: Shannon’s diversity index; and SHEI: Shannon’s evenness index.
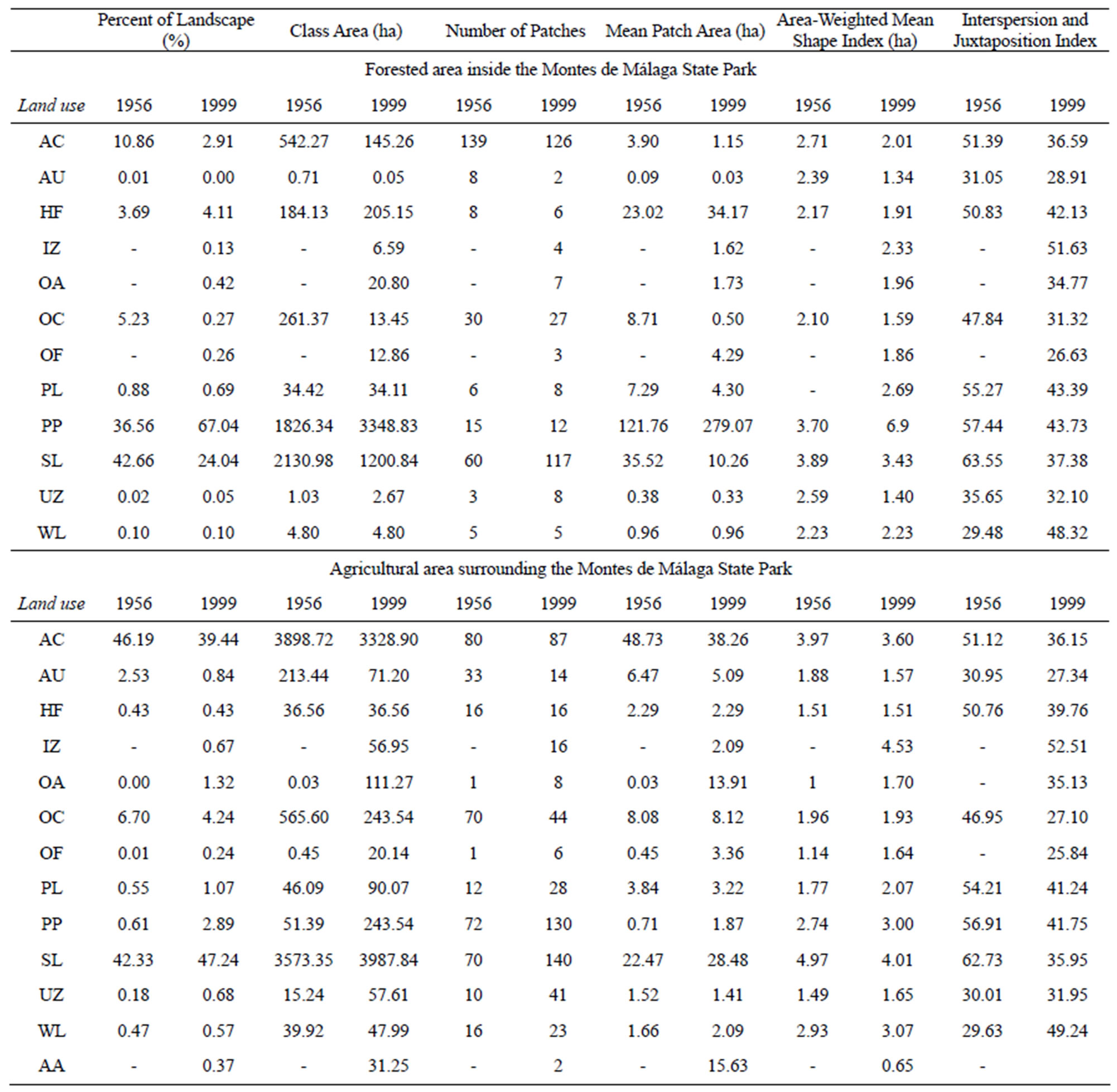
Table 5. Comparison of class metrics in 1956 and 1999 between the agricultural area surrounding the reserve and the forested areas inside of Montes de Málaga State Park (Andalusia, southern Spain). AC: Almond cultivation; AU: Agriculture-other use; HF: Holm oak forest; IZ: Industrial/infrastructure zone; OA: Abandoned olive cultivation; OC: Olive cultivation; OF: Other forest use; PL: Pasture land; PP: Pine plantation; SL: Shrub land; UZ: Urban zone; WL: Wetlands; AA: Crop land.
The interspersion and juxtaposition indices for pine plantation and Holm oak forest patch types decreased from 57 to 42 and 51 to 40, respectively. These results indicate that other patch types were converted into new forest types, mainly pine plantations and Holm oak between 1956 and 1999, and that these newly formed patches were more evenly distributed throughout the study area.
Surrounding the park, the area occupied by scrubland increased from 3573.3 ha in 1956 to 3,987.8 ha in 1999, whereas the number of tessellations dropped from 159 to 140. Almond tree orchards covered the largest area in 1956, but these diminished from 46% to 39% by 1999. A similar pattern occurred with the insertion and juxtaposetion indices, which decreased from 1956 to 1999, in contrast to the number of tessellations which increased from 80 to 87 in the same time period. The cause of the decline in both area and percent landscape cover was due to the transformation of the scrubland the abandoned almond trees plantations, and to the development of Industrial and commercial zones.
Abandoned almond and olive plantations behaved identically due to the number of tessellations, the relative area index and the insertion and juxtaposition indices increasing in 1999. As can be observed in Table 5, abandoned almond trees were not present in 1956 but they were in 1999, and although the area and number of tessellations, 31.25 and 2, respectively, were not very high, the relative area index reached a high value.
4. DISCUSSION
4.1. Species Richness in Pine Plantations
Changes in land cover and in other ecosystem features have a direct impact on societal expectations of forests, especially their links with nature conservation, global climate change, and recreation activities [5,30]. Understanding landscape and species patterns in restoration studies are emerging as important issue to identify and evaluate whether restoration goals have been met [U4] [31,32]. Forest plantations have been an important driving force of landscape change in Mediterranean areas. Therefore, changes in vegetation diversity due to forest plantations are a relevant ecological issue at regional level. [U5] However, the lack of any consistent information about the ecological attributes of these artificial environments, and specifically Aleppo pine plantations (P. halepensis), in Mediterranean countries makes it difficult to understand the changes on structural and ecological components. In this study, vegetation dynamics of Pinus halepensis plantations have been evaluated, indicating changes in species composition and vegetation structure.
Pinus halepensis forests in 1999 showed a higher species richness in the understory than those in agricultural land, which dominated the landscape before reforestation activities commenced. Forest plantations contain understory vegetation that is similar in diversity and composition to Quercus ilex forests, which can be considered a reference ecosystem. Vegetation dynamics of P. halepensis stands were driven by two factors: intensive forest management and natural succession, and the community gradually comes to resemble that of natural forests. There are two possible reasons for the greater species richness in the plantations compared to natural forests [33]. First, the soil seed bank, resprouts and seed rain affect the species richness of understory vegetation in forest plantations [13,14,34]. In this study, pine plantations were adjacent to natural forests (Figure 1), thus seed rain was likely higher than in the agricultural land, which was under cultivation during the same period. In addition, this may indicate that silviculture of P. halepensis activates germination of shade-tolerant species such as Quercus ilex. In surrounding patches, the influence of the seed sources on species richness should be considered a key factor in managing plantations for biodiversity [13,35,36,13], which can justify differences found between this study and others in semiarid Mediterranean areas in the Iberian Peninsula [37,38]. Second, species richness increases in response to silvicultural treatments such as thinning [39]. Pine plantations with lower canopy cover contain greater species richness than those with relatively closed canopies. Good light conditions and a thin litter layer promote the germination and recruitment of pine seeds as well as understory species, such as Arbutus unedo L., Quercus suber L., Olea europaea L. var. sylvestris Brot., . and Pistacia lentiscus L. For example, in Aleppo pine plantations in a dry Mediterranean climate, more open plantations were accompanied by greater oak abundance and species richness [35]. This suggests that canopy cover is a key structural factor when designing and managing plantations to increase biodiversity. The forests surveyed have been correctly managed during the past 50 years [23], and the beneficial effects of thinning on natural regeneration can be assumed [40]. This recolonization was likely possible only because of the vegetative structure of natural vegetation patches and plantations. Spatial interaction across patch boundaries (edge effect) between vegetation patches, therefore, acts as a trigger for patch dynamics [11].
These results suggest that forest plantations have an important role in promoting ecosystem functionality, and the floristic composition of these plantations will become similar to the surrounding evergreen oak forests due to the edge effect [19,41]. Patches of almond and olive cultivations were mainly located in the surrounding area and appeared as gaps in the forested landscape (Figure 1). However, if they are abandoned, tree crops will be invaded by species from surrounding patches. This is one of the features of vegetation dynamics in pine-dominated landscapes in Europe [42].
4.2. Changes in Landscape Metrics
In general, the mosaic landscape affects sink-source interactions such as seed flow and individual movement of species from diverse habitat patches [20,43,20]. Spatial information on landscape change is required for defining spatial patterns and ecological processes of landscape systems [44]. The land use dynamics explored in this study showed that a large deforested area (approximately 5,000 ha of the Guadalmedina river basin and Montes de Málaga State Park) was converted to mainly Aleppo pine plantations for revegetation and soil protection since the 1940s. A clear relationship between spatial vegetation pattern and dryland degradation has been shown by several authors [45,46].
Analyses of the land use changes that have generated the patterns observed in this study revealed that the establishment of P. halepensis in Montes de Málaga State Park has successfully met all the silvicultural and restoration objectives. First, a forest dominated by a Mediterranean and local native species, P. halepensis, was successfully created. Second, a transition process from the current dominant pine species to Holm oak forests has been initiated by natural regeneration in the study area. It is highly probable that the regenerating evergreen species will facilitate and extend the formation of mixed forest stands [13,14]. In Montes de Málaga State Park, major land use changes were due to the significant increase in pine plantations (1,522 ha) and Holm oak (21 ha), and the decreases in shrub land (931 ha), and almond (396 ha) and olive tree (Olea europea L.) cultivation (248 ha) between 1956 and 1999.
The effects of forest landscape changes on forest species were also demonstrated by fragmentation analysis. Landscape analyses using several landscape indices (Tables 1 and 4) were adapted for spatial pattern size using the spatial model FRAGSTAT [25]. These metrics were also used for measuring spatial heterogeneity [44]. Although the indices generated are partially correlated, each of them may make a unique contribution to the interpretation of the different spatial pattern aspects for the uses studied [47]. The landscape metrics calculated for Montes de Málaga and the transition maps of landscape patch types between 1956 and 1999 were helpful tools for analyzing the changes in landscape/vegetation structure and composition, and developing effective forest management strategies.
Indices related to patch size, TA (Total landscape area, ha), LPI (Largest patch index, %), NP (Number of patches, AREA MN (Mean patch area, ha), and SHAPE AM (Area weighted mean shape index) were used as an indicator of habitat function, such as habitat fragmentation (Table 1). Those indices showed the changes in landscape heterogeneity from 1956 to 1999 through landscape element and vegetation structure in agriculture and forests areas. A comparison of the landscape structure of the forested and agricultural land suggests that habitat heterogeneity was higher in the crop land than in the plantations. Several authors consider environmental heterogeneity to be an important factor, which promotes diversity [4,32]. However, the higher structural heterogeneity of the agricultural land is made up of crop areas with a lesser diversity than is observed in forest plantations. On the contrary, the structural homogeneity of the plantations might cover a lower number of available niches, but include greater species diversity.
Therefore, patches of a ‘ “natural unevenness’ were” replaced to ‘“regular evenness’ ” of a managed forest matrix of woody species communities. Edge indices (PR: Patch richness, SHDI: Shannon’s diversity index, and SHEI: Shannon’s evenness index), and shape metrics (IJI, Interspersion and juxtaposition index, and CONTAG, Contagion index) (Table 1) have also indicated greater complexity of landscape mosaics in agricultural land. However, interspersion and juxtaposition indices, showing the patch shape by patch area and perimeter, were similar in both land uses, which had complicated boundaries. In Mediterranean ecosystem, the edge effect at the landscape level is very important, particularly on ecotone-edge relationships [19,41]. Habitat connectivity has been identified as being one of the most important factors preserveing seed dispersal among populations [19,30,32,30].
4.3. Management Implications
In comparison to the surrounding agricultural land, the understory vegetation of pine plantations contained greater species richness, but also a more homogeneous landscape structure. However, because forest succession is influenced by various factors such as land management practices, canopy cover or seed sources, further studies in regions with these elements are required to define the general relationship between potential and current patterns of forest succession in plantations.
Both regeneration treatments and thinning used in Montes de Málaga State Park have been consciously designed to accelerate the formation of complex forest structures, such as secondary successional conditions [48]. The objective of increasing secondary successional conditions is a primary consideration in silviculture practices in forest plantations in the Mediterranean Basin. Currently, most stands contain dense, simple structures forming a low fragmented geographic pattern. Hence, landscape management intends to increase fragmentation and edge effects while increasing frequency and complexity of secondary successional structures at a rate that results in measurable, positive, changes in ecological functions.
Enriching species composition was helped by the presence of remaining Holm oak stands, integrated with P. halepensis. The presence of these stands of evergreen and shrub species facilitates diversification.[U6] A design following landscape ecology principles has permitted the development of a management plan and silvicultural activities that appear to be economically and biologically feasible, judging from presented results in the Montes de Málaga State Park, while the overall forest structure is beginning to regain a reasonable amount of secondary successional structures.
5. ACKNOWLEDGEMENTS
This project was funded through a Junta de Andalucía and project INTERBOS (CGL2008-04503-CO3-02) and DIVERBOS (CGL2011- 30285-C02-02) Diversidad funcional y resiliencia del bosque mediterráneo”.. . We also acknowledge the financial support from University of Córdoba-Campus de Excelencia CEIA3. We thank José Lopez Quintanilla for their contributions to the initial research proposal.
REFERENCES
- Myers, N., Mittermeier, R.A., Mittermeier, C.G., Fonseca, G.A., and Kent, J. (2000). Biodiversity hotspots for conservation priorities. Nature, 403, 853-858. doi:10.1038/35002501
- Fabbio, G., Merlo, M., and Tosi, V. (2003). Silvicultural management in maintaining biodiversity and resistance of forests in Europe—-the Mediterranean region. Journal of Environmental Management, 67 (1), 67-76. doi:10.1016/S0301-4797(02)00189-5
- Bielsa, I., Pons, X., and Bunce, B. (2005). Agricultural abandonment in the North Eastern Iberian Peninsula: the use of basic landscape metrics to support planning. Journal Environmental Planning and Management, 48 (1), 85–- 102. doi:10.1080/0964056042000308166
- Jomaaa, I., Audab, Y., Abi Salehc, B., Hamze, M., and Safi, S. (2008). Landscape spatial dynamics over 38 years under natural and anthropogenic pressures in Mount Lebanon. Landscape and Urban Planning, 87 (1), 67–-75.
- Lasanta, T., Gonzalez-Hidalgo, J.C., Vicente-Serrano, S., and Sferi, E. (2006). Using landscape ecology 516 to evaluate an alternative management scenario in abandoned Mediterranean mountain areas. Landscape and Urban Planning, 78 (1-2), 101–-114. doi:10.1016/j.landurbplan.2005.06.003
- Vélez, R. (1986). Fire prevention in Aleppo pine forests. Options Mediterraneennes, 1, 167-178.
- Mosser, D., Zechmeister, H.G., Plutzar, C., Sauberer, N., Wrbka, T., and Grabherr, G. (2002). Landscape patch shape complexity as an effective measure for plant species richness in rural landscapes. Landscape Ecology, 17, 657–-669. doi:10.1023/A:1021513729205
- Felton, A., Lindbladh, M., Brunet, J., and Fritz, O. (2010). Replacing coniferous monocultures with mixed-species production stands: An assessment of the potential benefits for forest biodiversity in northern Europe. Forest Ecology and Management, 260, 939-947. doi:10.1016/j.foreco.2010.06.011
- Stephens, S.S., and Wagner, M.R. (2007). Forest plantations and biodiversity: A fresh perspective. Journal of Forestry, 105, 307-313.
- Brockerhoff, E.G., Jactel, H., Parrotta, J.A., Quine, C.P., and Sayer, J. (2008). Plantation forests and biodiversity: oxymoron or opportunity? Biodiversity and Conservation, 17, 925-951. doi:10.1007/s10531-008-9380-x
- Bremer, L,. and Farley, K. (2010). Does plantation forestry restore biodiversity or create green deserts? A synthesis of the effects of land-use transitions on plant species richness. Biodiversity and Conservation, 19, 3893-3915. doi:10.1007/s10531-010-9936-4
- Atauri, J.A., de Pablo, C.L., de Agar, P.M., Schmitz, M.F., and Pineda, F.D. (2004). Effects of management on understory diversity in the forest ecosystems of northern Spain. Environmental Management, 34 (6), 819-828. doi:10.1007/s00267-004-0180-0
- Gomez-Aparicio, L., Zavala, M.A., Bonet, F.J. and Zamora, R. (2009) Are pine plantations valid tools for restoring Mediterranean forests? An assessment along abiotic and biotic gradients. Ecological Applications 19, 2124-2141. doi:10.1890/08-1656.1
- Urbieta, I., Garcia, L., Zavala, M.A., and Marañón, T. (2010). Mediterranean pine and oak distribution in the southern of Spain: Is there a mismatch between regeneration and adult distributions? Journal Vegetation of Science, 22, 18-31. doi:10.1111/j.1654-1103.2010.01222.x
- Mendoza, I., Gómez-Aparicio, L., Zamora, R., and Matías, L. (2009). Recruitment limitation of forest communities in a degraded Mediterranean landscape. Journal of Vegetation Science, 20, 367-376. doi:10.1111/j.1654-1103.2009.05705.x
- Diaci, J. (2002). Regeneration dynamics in a Norway spruce plantation on a silver fir— – beech forest site in the Slovenia Alps. Forest Ecology and Management, 161 (1-3), 27-38. doi:10.1016/S0378-1127(01)00492-3
- Swindel, B.F., and Grosenbaugh, L.R. (1988). Species diversity in young Douglas-fir plantations compared to old growth. Forest Ecology and Management, 23 (2-3), 227–- 231. doi:10.1016/0378-1127(88)90084-9
- Brockerhoff, E.G., Ecroyd, C.E., Leckie, A.C., and Kimberley, M.O. (2003). Diversity and succession of adventine and indigenous vascular understorey plants in Pinus radiata plantation forests in New Zealand. Forest Ecology and Management, 185 (3), 307–-326. doi:10.1016/S0378-1127(03)00227-5
- Alados C.L., Pueyo Y., Barrantes O., Escós J., Giner L. and Robles, A. (2004). Variations in landscape patterns and vegetation cover between 1957 and 1994 in a semiarid Mediterranean ecosystem. Landscape ecology, 19 (5), 543-559. doi:10.1023/B:LAND.0000036149.96664.9a
- Ferrier, S., and Drielsma, M. (2010). Synthesis of pattern and process in biodiversity conservation assessment: a flexible whole-landscape modelling framework. Diversity and Distributions, 16 (3), 386–-402. doi:10.1111/j.1472-4642.2010.00657.x
- CSIC (1989). Mapa de Suelos de Andalucía. Junta de Andalucía., Sevilla.
- Rivas Martínez, S. (1987). Memoria del mapa de series de vegetación de España. (Vegetation map of Spain). ICONA, Ministerio de Agricultura, Pesca y Alimentación., Madrid.
- Consejería de Medio Ambiente, (2003). Plan de ordenación de los recursos naturales y el plan rector de uso y gestión del parque natural montes de Málaga. (Management Plan of Natural Resources of Montes de Málaga State Park) Consejería de Medio Ambiente, Junta de Andalucía., Sevilla.
- Elzinga, C.L., Salzer, D., and Willoughby, J. (1999). Measuring and monitoring plant populations. United States Government PrintingBLM Technical Reference 1730–1. U.S. Department of the Interior, Arlington, VirginiaWashington DC.
- McGarigal, K., and Marks, B.J. (1995). FRAGSTATS: spatial pattern analysis program for quantifying landscape structure. USDA Forest Service General Technical Report PNW-351., Corvallis, Oregon, U.S.A.
- Tinker, D.B., Romme, W.H., and Despain, D.G. (2003). Historic range of variability in landscape structure in subalpine forests of the Grater Yellowstone Area, USA. Landscape ecology, 18 (4), 427- 439. doi:10.1023/A:1026156900092
- Matsushita, B., Xu, M., and Fukushima, T. (2005). Characterizing the changes in landscape structure in the Lake Kasumigaura Basin, Japan using a high-quality GIS dataset. Landscape and Urban PlanningLandscape Urban Planning, 78 (3), 241-250. doi:10.1016/j.landurbplan.2005.08.003
- Leitao, A.B., and Ahem, J. (2002). Applying landscape ecological concepts and metrics in sustainable landscape planning. Landscape and Urban Planning, 59 (2), 65-93. doi:10.1016/S0169-2046(02)00005-1
- Peet, R.K. (1975). Relative diversity indices. Ecology, 56 (2), 496-498. doi:10.2307/1934984
- Laliberté, E., Wells, J.A., DeClerck, F., Metcalfe, D.J., Catterall, C.P., Queiroz, C., Aubin, I., Bonser, S.P., Ding, Y., Fraterrigo, J.M., McNamara, S., Morgan, J.W., Sánchez Merlos, D., Vesk, P.A., and Mayfield, M.M. (2010). Land-use intensification reduces functional redundancy and response diversity in plant communities. Ecology Letters, 13 (1), 76–-86. doi:10.1111/j.1461-0248.2009.01403.x
- Korb, J.E., Covington, W.W., and Fulé, P.Z. (2003). Sampling techniques influence understory plant trajectories after restoration: an example from Ponderosa pine restoration. Restoration ecology, 11 (4), 504-515. doi:10.1046/j.1526-100X.2003.rec0170.x
- Mayfield, M.M., Bonser, S.P., Morgan, J.W., Aubin, I., McNamara, S., and Vesk, P.A. (2010). What does species richness tell us about functional trait diversity? Predictions and evidence for responses of species and functional trait diversity to land-use change. Global Ecology and Biogeography, 19, 423–-431.
- Igarashi, T., and Kiyono, Y. (2008). The potential of hinoki (Chamaecyparis obtusa [Sieb. et Zucc.] Endlicher) plantation forests for the restoration of the original plant community in Japan. Forest Ecology and Management, 255 (1), 183–-192. doi:10.1016/j.foreco.2007.09.003
- Zamora, R., Hódar, J.A., Matías, L., and Mendoza, I. (2010.) Positive adjacency effects mediated by seed disperser birds in pine plantations. Ecological Applications, 20, 1053-1060. doi:10.1890/09-0055.1
- Zavala, M.A., and Zea, E. (2004) Mechanisms maintaining biodiversity in Mediterranean pine-oak forests: insight from a spatial simulation model. Plant ecology, 171(1-2), 197-207. doi:10.1023/B:VEGE.0000029387.15947.b7
- Purves, D.W., Zavala, M.A., Ogle, K., Prieto, F., and Benayas, J.M.R. (2007). Environmental heterogeneity, bird-mediated directed dispersal and oak woodland dynamics in Mediterranean Spain. Ecological Monographs, 77, 77-97. doi:10.1890/05-1923
- Maestre, F.T., and Cortina, J. (2004). Are Pinus halepensis plantations useful as a restoration tool in semiarid Mediterranean areas? Forest Ecology and Management, 198 (1-3), 303-317. doi:10.1016/j.foreco.2004.05.040
- Navarro-Cano, J.A., Barberá, G.G., Ruiz-Navarro, A., and Castillo, V.M. (2009). Pine plantation bands limit seedling recruitment of a prennial grass under semiarid conditions. Journal of Arid Enviroment, 73 (1), 120-126. doi:10.1016/j.jaridenv.2008.09.002
- Battles, J.J., Shlisky, A.J., Barrett, R.H., Heald, R.C., and Allen-Diaz, B.H. (2001). The effects of forest management on plant species diversity in a Sierran conifer forest. Forest Ecology and Management, 146 (1-3), 211–-222. doi:10.1016/S0378-1127(00)00463-1
- Thomas, S.C., Halpern, C.B., Falk, D.A., Liguori, D.A., and Austin, K.A. (1999). Plant diversity in 562 managed forests: understory responses to thinning and fertilization. Ecological Application, 9(3), 864–-879.
- Yue, T.X., and Li, Q.Q. (2010). Relationship between species diversity and ecotope diversity. Annals of the New York Academy of Sciences, 1195, E40–-E51. doi:10.1111/j.1749-6632.2009.05417.x
- Rabbinge, R., and van Diepen, C.A. (2000). Changes in agriculture and land use in Europe. European Journal of Agronomy, 13, 85–-100. doi:10.1016/S1161-0301(00)00067-8
- Jeanneret, P., Schüpbach, B., and Luka, H. (2003). Quantifying the impact of landscape and habitat features on biodiversity in cultivated landscape. Agricultural Ecosystem Environment, 98, 311–-320. doi:10.1016/S0167-8809(03)00091-4
- Turner, M.G., Gardner, R.H., and O’Neill, R.V. (2001). Landscape ecology: theory and practice. Springer, Berlin Heidelberg New York.
- Kefi, S., Rietkerk, M., Alados, C., Pueyo, Y., Papanastasis, V., Elaich, A., and de Ruiter, P. (2007). Spatial vegetation patterns and imminent desertificatiion in Mediterranean arid ecosystems. Nature, 507, 213-217. doi:10.1038/nature06111
- Bautista, S., Mayor, A., Bourakhouadar, J., and Bellot, J. (2007). Plant spatial pattern predicts hillslope runoff and erosion in a semiarid mediterranean landscape. Ecosystems, 10(6), 987-998. doi:10.1007/s10021-007-9074-3
- Garrabou, J., Riera, J., and Zabala, M. (1998). Landscape pattern indices applied to Mediterranean subtidal rocky benthic communities. Landscape ecology, 13 (4), 225-247. doi:10.1023/A:1007952701795
- Osem, Y., Zangy, E., Bney-Moshe, E., Moshe, Y., Karni, N. and Nisan, Y. (2009) The potential of transforming simple structured pine plantations into mixed Mediterranean forests through natural regeneration along a rainfall gradient. Forest Ecology and Management, 259, 14-23.