P. Wu et al. / Agricultural Science 2 (2011) 201-207
Copyright © 2011 SciRes. Openly accessible at http://www.scirp.org/journal/AS/
207207
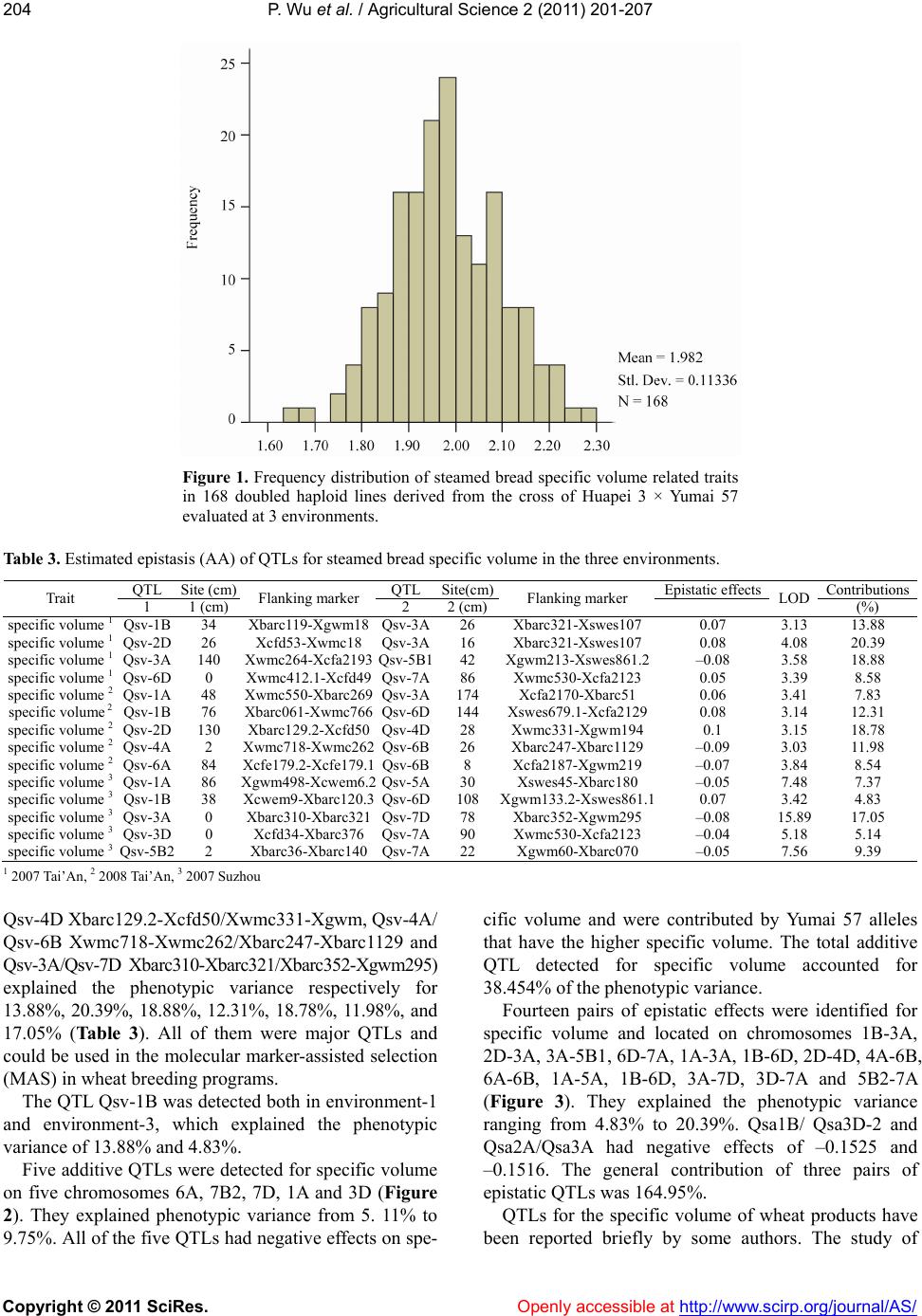
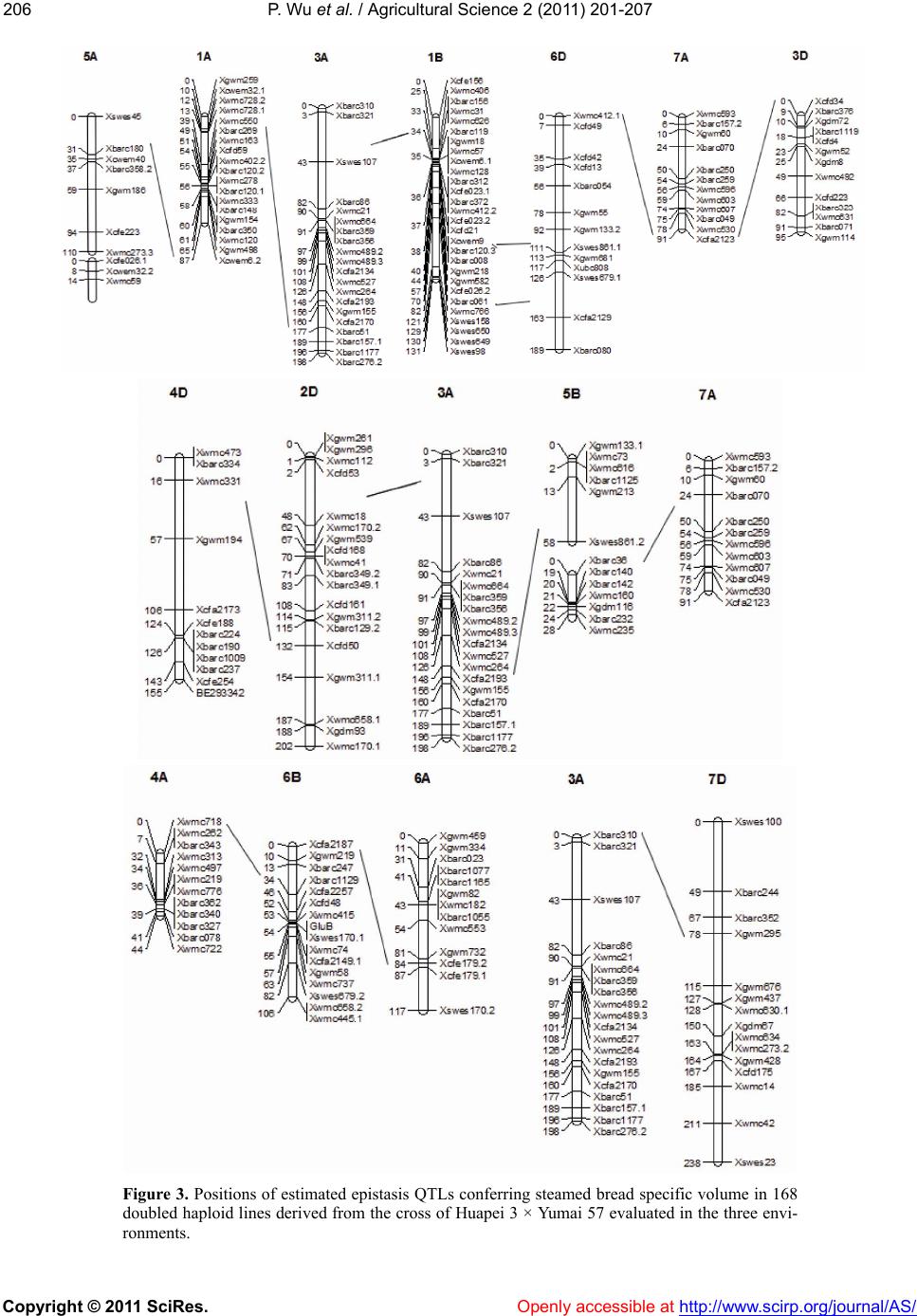
12.31%, 18.78%, 11.98% and 17.05% of the phenotypic
variation of specific volume. The information obtained
in this study will be useful for manipulating the QTLs
for CNSB specific volume property by molecular
marker-assisted selection (MAS).
5. ACKNOWLEDGEMENTS
The authors thank gratefully Dr. C.E.(Chuck)Walker (Kansas State
University) for his editorial suggestions and careful reading of the
manuscript. The DH population and parents were kindly provided by
Professor Yan Hai, Henan Academy of Agricultural Sciences, Zheng-
zhou, China. Fin ancial support was provided by the 973 program of
the National Nature Science Foundation of China (2009CB118301),
and the National Natural Science Foundation of China (No. 30971764)
from the Ministry of Science and Technology of the People’s Republic
of China.
REFERENCES
[1] He, Z.H., Liu, A.H., Peña, R.J. and Rajaram, S. (2003)
Suitability of Chinese wheat cultivars for production of
northern style Chinese steamed bread. Euphytica, 131,
155-163. doi:10.1023/A:1023929513167
[2] Guo, B.L., Wei, Y.M., Zhang, G.Q.,Yang, S.H., Hu, X.Zh.
(2002) Study on the quality judging methods of steamed
bread. Journal of Triticeae Crops, 22, 7-10
[3] Hai, Y., and Kang, M.H. (2007) Breeding of a new wheat
variety Huapei 3 with high yield and early maturing.
Henan Agriculture Science, 5, 36-37.
[4] Nelson, J.C. Andreescu, C., Breseghello, F., Finney, P.L.,
Gualberto, D.G., Bergman, C.J., Pena, R.J., Perretant,
M.R., Leroy, P., Qualset, C.O. and Sorrells, M.E. (2006)
Quantitative trait locus analysis of wheat quality traits,
Euphytica, 149, 145-159.
doi:10.1007/s10681-005-9062-7
[5] Hai, Y. and Kang, M.H. (2007) Breeding of a new wheat
variety Huapei 3 with high yield and early maturing.
Henan Agriculture Science, 5, 36-37.
[6] Guo, C.Q., Bai, Z.A., Liao, P.A. and Jin, W.K. (2004)
New high quality and yield wheat variety Yumai 57.
China Seed Industry, 4, 54.
[7] SB/T10139-93 (1993) wheat flour for steamed bread,
Appendix A.
[8] Somers, D.J., Isaac, P. and Edwards, K. (2004) A high-
density microsatellite consensus map for bread wheat
(Triticum aestivum L.). Theoretical and Applied Genetics,
109, 1105-1114. doi:10.1007/s00122-004-1740-7
[9] Doerge, R.W. (2002) Multifactorial genetics: Mapping
and analysis of quantitative trait loci in experimental
populations. Nature Reviews, 3, 43-52.
doi:10.1038/nrg703
[10] Lincoln, S.E., Daly, M.J. and Lander, E.S. (1993) Con-
structing genetic maps with MAPMAKER/EXP version
3.0: A tutorial and reference manual. Whitehead Institute
for Biomedical Research, Cambridge.
[11] Kosambi, D.D. (1944) The estimation of map distances
from recombination values. Annals of Human Genetics,
12, 172-175. doi: 10 .1111 /j. 1469-1809.1943.tb02321.x
[12] Voorrips, R.E. (2002) MapChart: Software for the
graphical presentation of linkage maps and QTL. Journal
of Heredity, 93, 77-78. doi:10.1093/jhered/93.1.77
[13] Yang, J. and Zhu, J. (2005) Predicting superior genotypes
in multiple environments based on QTL effects. Theo-
retical and Applied Genetics, 11 0, 1268-1274.
doi:10.1007/s00122-005-1963-2
[14] Wang, D.L., Zhu, J., Li, Z.K. and Paterson, A.H. (1999)
Mapping QTLs with epistatic effects and QTL environ-
ment interactions by mixed linear model approaches.
Theoretical and Applied Genetics, 99, 1255-1264.
doi:10.1007/s001220051331
[15] Cao, G., Zhu, J., He, C., Ga o , Y. , Yan, J. and Wu, P. (2001)
Impact of epistasis and QTL environment interaction on
the developmental behavior of plant height in rice (Oryza
sativa L.). Theoretical and Applied Genetics, 103, 153-
160. doi:10.1007/s001220100536
[16] Li, H., Li, Z. and Wang, J. (2008) Inclusive composite
interval mapping (ICIM) for digenic epistasis of quanti-
tative traits in biparental populations. Theoretical and
Applied Genetics, 116, 243-260.
doi:10.1007/s00122-007-0663-5
[17] Elangovan, M.R., Rai, B.B., Dholakia, M.D., Lagu, R.,
Tiwari, R.K., Gupta, V.S. Rao, M.S. and Gupta V.S.
(2008) Molecular genetic mapping of quantitative trait
loci associated with loaf volume in hexaploid wheat
(Triticum aestivum). Journal of Cereal Science, 47, 587-
598. doi:10.1016/j.jcs.2007.07.003
[18] Fan, Y. D., Sun, H.Y., Zhao, J.L., Ma, Y.M., Li, R.J., Li,
S.Sh. (2009) QTL mapping for quality traits of north-
ern-style hand-made Chinese steamed bread. Journal of
Cereal Science, 49, 225-229.
doi:10.1016/j.jcs.2008.10.004
[19] Zhang K.P., Tian J.C., Zhao L., Wang S.S. (2008) Map-
ping QTLs with epistatic effects and QTL environment
interactions for plant height using a doubled haploid
population in cultivated wheat. Journal of Genetics &
Genomics, 35, 119-127.
doi:10.1016/S1673-8527(08)60017-X