Open Journal of Soil Science
Vol.3 No.8(2013), Article ID:41596,5 pages DOI:10.4236/ojss.2013.38043
Glomalin and Soil Aggregation under Six Management Systems in the Northern Great Plains, USA
![]()
1USDA-ARS Northern Great Plains Research Laboratory, Mandan, USA; 2Precision Soil Management, Redfield, USA.
Email: Kristine.nichols@ars.usda.gov
Copyright © 2013 Kristine A. Nichols, James Millar. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received September 21st, 2013; revised October 21st, 2013; accepted October 28th, 2013
Keywords: Glomalin; Arbuscular Mycorrhizal Fungi; Soil Aggregation; Grazing Systems; Crop Production Systems
ABSTRACT
The soil environment is linked to aboveground management including plant species composition, grazing intensity, levels of soil disturbance, residue management, and the length of time of a living plant is growing. Soil samples were collected under rangeland [native grass, rotational grazing (NGRG); tame grass, heavy grazing (TGRG); and tame grass, rotational grazing (TGHG)] and cropland [conventional till (CT); CT plus manure (CTM); and long term no till (NT)] systems. The rangeland systems were hypothesized to have higher glomalin content [measured as Bradford-reactive soil protein (BRSP)] and water stable aggregation (WSA) than the cropland systems. In addition, within both rangeland and cropland systems, BRSP and WSA were expected to decline with increased disturbance due to grazing or tillage and going from native to introduced plant species. Differences were detected for BRSP with NGRG and CTM having the highest values in range and cropland systems, respectively. However, the CTM system had higher BRSP values than one or both of the tame grass systems while the CT and NT systems had similar values. Correlation analysis showed strong relationships between all of the BRSP values and WSA.
1. Introduction
The symbiotic relationship between arbuscular mycorrhizal fungi (AMF) and about 90% of all vascular plants provides a direct linkage between above and belowground activities [1,2]. Soil aggregation is a mechanism used to engineer the soil environment to maximize water and nutrient use efficiency [3]. Glomalin is a glycoprotein produced by AMF which is strongly linked to aggregation [4-6]. The aggregation process and glomalin production are both a carbon cost to AMF and may be hindered by soil disturbance through tillage [4-11] or overgrazing [12-20]. Therefore, the level of mycorrhizal dependency [1] of the host plant as well as photosynthetic activity may impact glomalin content and water stable aggregation (WSA) [2].
In rangeland systems, Liebig et al. [21] found that systems under heavy grazing intensity had higher soil C values than systems under moderate intensity. However, Klumpp et al. [19] and Ingram et al. [20] found that increases in grazing intensity decreased soil C. Unlike the Liebig et al. [21] study, these authors also examined microbial community and soil C dynamics under different grazing treatments and found that the rhizosphere environment was the driver behind changes in soil C [19,20]. These results are worth noting because often soil C levels are related to soil aggregation and glomalin concentrations [2, 22].
In cropland systems, no till (NT) treatments had higher soil organic C levels, glomalin concentrations, and more water stable aggregates than conventional till (CT) treatments [5-8,11]. These differences tended to increase with longer periods of time in NT [5,6]. Pikul et al. [4] found that particulate organic matter (POM) increased under NT while aggregate wettability decreased. Greater amounts of POM may support higher aggregate stability and water holding capacity while a decrease in aggregate wettability may negatively impact water infiltration and water holding capacity. In addition to C and N levels and aggregate stability, glomalin values also may be related to other AMF parameters such as spore counts, hyphal length or root colonization but these relationships are inconsistent [7,8,13,15,18,23].
Bradford-reactive soil protein (BRSP) and water stable aggregation (WSA) were measured at six sites near Platte, SD, USA. These sites included three rangelands [native grass, rotational grazing (NGRG); tame grass, heavy grazing (TGRG); and tame grass, rotational grazing (TGHG)] and three croplands [conventional till (CT); CT plus manure (CTM); and long term no till (NT)] systems. We hypothesized that the rangeland systems would have higher BRSP and WSA values than the cropland systems and values would increase across these systems in the following order: CT < CTM < NT < TGHG < TGRG < NGRG.
2. Materials and Methods
2.1. Site Description
Soil samples were collected on June 7, 2004 near Platte, SD, USA from under six management systems (NGRG, TGRG, TGHG, CT, CTM, and NT). The sites were selected to show a wide range of management scenarios on the same soil type (an Eakin silty clay loam or fine-silty, mixed, superactive, mesic Typic Argiustolls) and under the same climatic conditions (i.e. all six sites within a 10-km radius). The major grass species at the NGRG was big bluestem (Andropogon gerardii) with a small amount of switchgrass (Panicum virgatum L.). The grass species at the TGRG sites were predominantly bromegrass (Bromus inermus) and crested wheatgrass (Agropyron cristatum) with small amounts of intermediate wheatgrass (Thinopyrum intermedium). The grass species at the TGHG was Kentucky bluegrass (Poa pratensis) with some bromegrass. The cropping systems all had a cornwheat rotation. Two samples were collected from each site at the 0 to 10 cm depth to create a bulk sample. A portion of the soil was sequentially passed through a series of screens to collect 1 to 2 mm aggregates.
2.2. Soil Analyses
Six 2-g subsamples were removed from both bulk soil and 1 to 2 mm aggregate samples and extracted for glomalin-related soil protein (GRSP). Soil was extracted with 50 mM sodium citrate, pH 8.0, for 15 to 26 1-hr cycles at 121˚C [24] followed by extraction with 100 mM sodium pyrophosphate, pH 9.0, for four to nine 1-hr cycles at 121˚C to remove a recalcitrant fraction of glomalin [2]. For both the citrate and the sodium pyrophosphate extractions, samples were centrifuged after each 1-hr extraction cycle to pellet the soil and collect the supernatant. All citrate or pyrophosphate extractions were combined, and the total volume measured.
The Bradford total protein assay was used to measure GRSP as Bradford reactive soil protein (BRSP) [2]. For the samples extracted with pyrophosphate, an aliquot equal in volume to the aliquot used for the unknowns of 100 mM pyrophosphate was added to the BSA standard to remove background cross-reactivity. Total protein values are calculated on a mg Bradford-reactive soil protein (BRSP) g−1 soil or 1 to 2 mm aggregates basis. Both the bulk soil and 1 to 2 mm aggregates were corrected for coarse material in the sample using forced water to pass the extracted soil or aggregates through a 1 mm screen leaving the coarse fraction on the screen. Water stable aggregation (WSA) was measured on four subsamples of the 1 to 2 mm aggregates using methodology modified from Kemper and Rosenau [25] and described by Nichols and Wright [2].
2.3. Statistical Analysis
Means and standard error of the mean (SEM) were calculated for the BRSP and WSA. All means comparisons were made at the a ≤ 0.05 level using PROC MIXED (SAS software, ver. 9.1, SAS Institute, 2004) after the residuals met the assumptions for normality and homogeneity of variance. The correlations between BRSP, for both the citrate and pyrophosphate extractions and WSA were measured using PROC CORR after meeting the assumption for normal data distribution (SAS software, ver. 9.1, SAS Institute, 2004).
3. Results and Discussion
Concentrations of BRSP in both the bulk soil and 1 to 2 mm aggregate samples were similar for each of the individual management scenarios with the exception of the CT site which was lower in the bulk soil than in the 1 to 2 mm aggregates (Table 1). The three grazing systems had the highest glomalin concentrations with the NGRG system having the significantly highest values. Because most studies have shown that as grazing intensity increases, soil carbon levels and microbial activities decrease [19], it was unexpected that the TGRG site would have lower BRSP values than the TGHG system. However, a study by Liebig et al. [21] showed that a heavily grazed treatment had higher soil carbon levels than a moderately grazed treatment at sites in North Dakota which were in the same semi-arid ecoregion as the sites in this study but were approximately 560 km apart. These studies did indicate that the results were probably dependent upon if the plant species composition was dominated by native or tame grasses which impacts the intensity of the mycorrhizal relationship [1,13] and impacts rooting depth and rhizosphere dynamics [19-21].
In the crop production systems, the CTM site had the highest values while there was no difference between the CT and NT systems in the bulk soil but the CT site had significantly higher BRSP in the 1 to 2 mm aggregates. This was also unexpected since most studies have shown glomalin, or BRSP, increases with longer periods of NT [4-6]. The addition of manure has been shown to increase
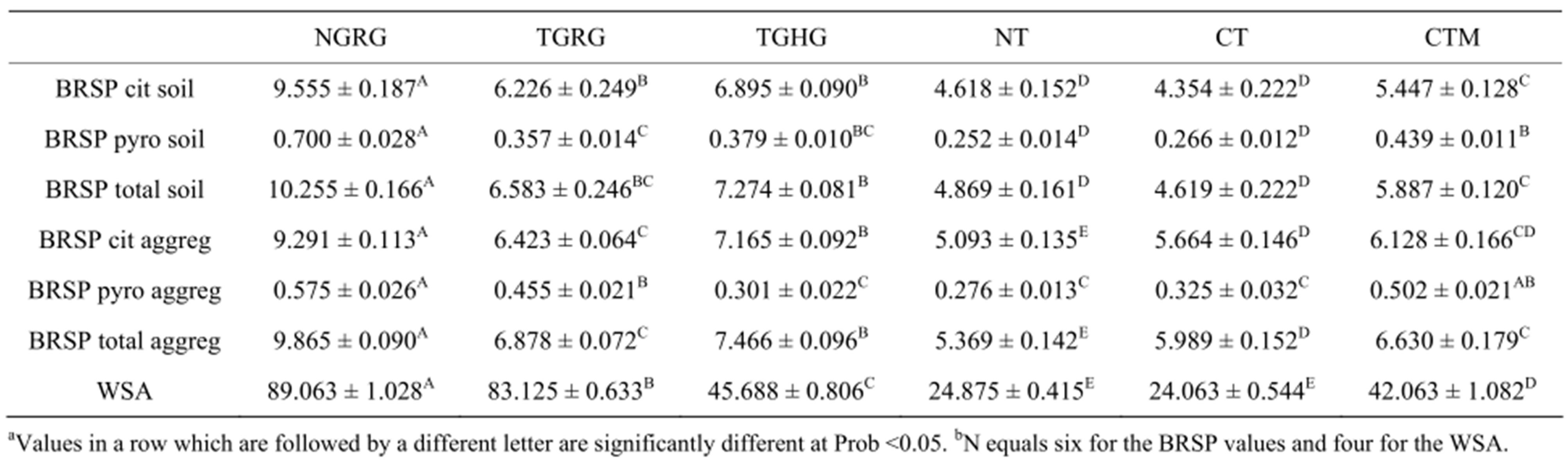
Table 1. Valuesa for glomalin extracted with sodium citrate (cit) followed by sodium pyrophosphate (pyro) from either bulk soil or 1 to 2 mm aggregates (aggreg) and measured as Bradford reactive soil protein (BRSP) on a mg protein g−1 soil or aggregate weight basis and for water stable aggregation (WSA) at a% of total aggregates. Values are meansb ± SEs for each management system: native grass, rotational grazing (NGRG); tame grass, heavy grazing (TGRG); tame grass, rotational grazing (TGHG); long term no till (NT); conventional till (CT); and CT plus manure (CTM).
soil C, which may stimulate aggregation, and BRSP (Wright, personal communication). In addition, other organic compounds, such as humic substances and tannins may be co-extracted with glomalin and measured with the Bradford assay [2,26,27]. This might explain the CTM treatment values, but it was unclear in this study why the long term NT system did not have higher BRSP values than the CT site.
The authors speculate that the crops grown at the cropland sites might have impacted carbon allocation to AMF and AMF allocation to glomalin production and aggregation.
Despite these unexpected results from BRSP and WSA values, the Pearson Correlation Coeffcients showed BRSP was strongly and significantly correlated WSA, r > 0.7 in almost all cases (Table 2). This is similar to many other studies where BRSP and WSA are correlated.
Native warm-season grasses, such as big bluestem and switchgrass, are all highly mycorrhizal while cool-season grasses are less dependent upon the mycorrhizal relationship [1,13]. Mycorrhizal fungi and glomalin production are sensitive to carbon exudates from the plant and soil disturbance. Additionally, AMF and glomalin are important in forming and stabilizing aggregates which play significant roles in water infiltration and water retention [5,6,14]. Finally, grazing pressure has been shown to decrease the amount of carbon fixed by photosynthesis and exuded from the roots [19,20]. For the most part, this did occur in this study but given the non-replicated design and the limited number of samples collected at each site, and the limited number of studies on glomalin and WSA in rangeland systems in the literature, more research is needed. Future studies will focus on the biological, chemical, and physical interactions occurring in the soil, particularly in the rhizosphere, which may be used to model above and belowground relationships. These models will help us understand the linkage between levels of soil disturbance and the types and amounts of photosynthetically-derived carbon going belowground, especially as it relates to the plant-mycorrhizal relationship. Ongoing research projects in the northern Great Plains are measuring GRSP and WSA in other range and cropland studies. The range studies include systems with long-term (over 60 years) continuous management different levels of grazing intensity from no grazing to heavy grazing while the cropland studies incorporate perennial and cover crops which may be used as forages.
4. Conclusion
Similar to many other studies, glomalin and aggregate stability were strongly correlated in this study. However, the BRSP and WSA values did not increase from CT < CTM < NT < TGHG < TGRG < NGRG as expected. This was noted in particular in the cropland sites where CTM had the highest values and CT and NT were similar. The complex relationship between AMF and host plant especially as it relates to carbon allocation needs more studies to understand glomalin production and how it relates to aggregation.
5. Abbreviations and Acronyms
BRSP, Bradford reactive soil protein; C, carbon; CT, conventional till; CTM, conventional till plus manure; GRSP, glomalin-related soil protein; IR, infiltration rate; NGRG, native grass rotational grazing; NT, long-term no-till; TGHG, tame grass heavy grazing; TGRG, tame grass rotational grazing; WHCs, water holding capacity; WSA, water stable aggregation.
6. Acknowledgements
The authors would like to acknowledge Crystal Jundt and Gary Brucker for their assistance with the laboratory
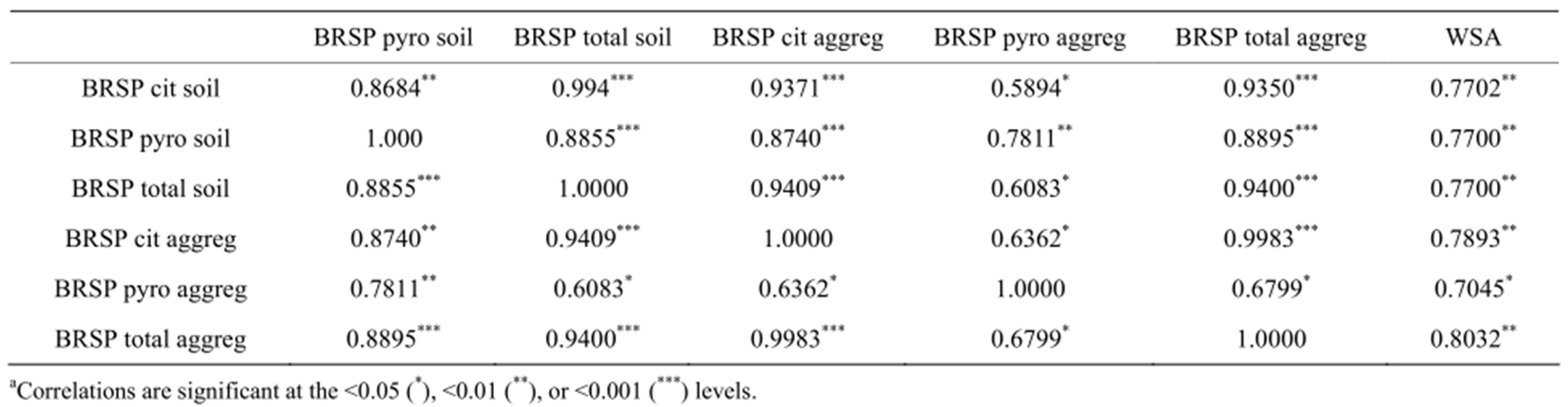
Table 2. Correlation coefficientsa for glomalin extracted with sodium citrate (cit) followed by sodium pyrophosphate (pyro) from either bulk soil or 1 to 2 mm aggregates (aggreg) and measured as Bradford reactive soil protein (BRSP) on a mg protein g−1 soil or aggregate weight basis, for water stable aggregation (WSA) at a% of total aggregates, for the infiltration rate (IR) on a ml per hr basis for the first (1) and second (2) inches of water, and for the water holding capacity at saturation (WHCs) for the first two inches of water.
analysis, and Drs. Rebecca Phillips and Shannon Osborne for their invaluable comments on earlier drafts of this manuscript.
REFERENCES
- M. F. Allen, W. Swenson, J. I. Querejeta, L. M. EgertonWarburton and K. K. Treseder, “Ecology of Mycorrhizae: A Conceptual Framework for Complex Interactions among Plants and Fungi,” Annual Review of Phytopathology, Vol. 41, 2003, pp. 271-303. http://dx.doi.org/10.1146/annurev.phyto.41.052002.095518
- K. A. Nichols and S. F. Wright, “Contributions of Soil Fungi to Organic Matter in Agricultural Soils,” In: F. R. Magdoff and R. R. Weil, Eds., Soil Organic Matter in Sustainable Agriculture, CRC Press, New York, 2004, pp. 179-198. http://dx.doi.org/10.1201/9780203496374.ch6
- M. C. Rillig and P. D. Steinberg, “Glomalin Production by an Arbuscular Mycorrhizal Fungus: A Mechanism of Habitat Modification?” Soil Biology and Biochemistry, Vol. 34, No. 9, 2002, pp. 1371-1374. http://dx.doi.org/10.1016/S0038-0717(02)00060-3
- J. Pikul Jr., G. Chilom, J. Rice, A. Eynard, T. E. Schumacher, K. A. Nichols, J. M. F. Johnson, S. Wright, T. C. Caesar-Tonthat and M. Ellsbury, “Organic Matter and Water Stability of Field Aggregates Affected by Tillage in South Dakota,” Soil Science Society of America Journal, Vol. 73, 2009, pp. 197-206. http://dx.doi.org/10.2136/sssaj2007.0184
- S. F. Wright and R. L. Anderson, “Aggregate Stability and Glomalin in Alternative Crop Rotations for the Central Great Plains,” Biology and Fertility of Soils, Vol. 31, No. 3/4, 2000, pp. 249-253. http://dx.doi.org/10.1007/s003740050653
- S. F. Wright, J. L. Starr and I. C. Paltineanu, “Changes in Aggregate Stability and Concentration of Glomalin during Tillage Management Transition,” Soil Science Society of America Journal, Vol. 63, No. 6, 1999, pp. 1825-1829. http://dx.doi.org/10.2136/sssaj1999.6361825x
- F. Borie, R. Rubio, J. L. Rouanet, A. Morales, G. Borie and C. Rojas, “Effects of Tillage Systems on Soil Characteristics, Glomalin and Mycorrhizal Propagules in a Chilean Ultisol,” Soil and Tillage Research, Vol. 88, No. 1-2, 2006, pp. 253-261. http://dx.doi.org/10.1016/j.still.2005.06.004
- T. Caesar-Tonthat, U. M. Sainju, S. F. Wright, W. L. Shelver, R. L. Kolberg and M. West, “Long-Term Tillage and Cropping Effects on Microbiological Properties Associated with Aggregation in a Semi-Arid Soil,” Biology and Fertility of Soils, Vol. 47, No. 2, 2010, pp. 157-165. http://dx.doi.org/10.1007/s00374-010-0508-2
- M. C. Rillig, “Arbuscular Mycorrhizae, Glomalin, and Soil Aggregation,” Canadian Journal of Soil Science, Vol. 84, No. 4, 2004, pp. 355-363. http://dx.doi.org/10.4141/S04-003
- A. Roldán, J. R. Salinas-García, M. M. Alguacil and F. Caravaca, “Soil Sustainability Indicators Following Conservation Tillage Practices under Subtropical Maize and Bean Crops,” Soil and Tillage Research, Vol. 93, No. 2, 2007, pp. 273-282. http://dx.doi.org/10.1016/j.still.2006.05.001
- S. B. Wuest, T. C. Caesar-Tonthat, S. F. Wright and J. D. Williams, “Organic Matter Addition, N, and Residue Burning Effects on Infiltration, Biological, and Physical Properties of an Intensively Tilled Silt-Loam Soil,” Soil and Tillage Research, Vol. 84, No. 2, 2005, pp. 154-167. http://dx.doi.org/10.1016/j.still.2004.11.008
- K. M. Batten, K. M. Scow, K. F. Davies and S. P. Harrison, “Two Invasive Plants Alter Soil Microbial Community Composition in Serpentine Grasslands,” Biological Invasions, Vol. 8, No. 2, 2006, pp. 217-230. http://dx.doi.org/10.1007/s10530-004-3856-8
- M. A. Bingham and M. Biondini, “Mycorrhizal Hyphal Length as a Function of Plant Community Richness and Composition in Restored Northern Tallgrass Prairies (USA),” Rangeland Ecology and Management, Vol. 62, No. 1, 2009, pp. 60-67. http://dx.doi.org/10.2111/08-088
- S. B. Bird, J. E. Herrick, M. M. Wander and S. F. Wright, “Spatial Heterogeneity of Aggregate Stability and Soil Carbon in Semi-Arid Rangeland,” Environmental Pollution, Vol. 116, No. 3, 2002, pp. 445-455. http://dx.doi.org/10.1016/S0269-7491(01)00222-6
- J. S. Buyer, D. A. Zuberer, K. A. Nichols and A. J. Franzluebbers, “Soil Microbial Community Function, Structure, and Glomalin in Response to Tall Fescue Endophyte Infection,” Plant and Soil, Vol. 339, No. 1-2, 2011, pp. 401-412. http://dx.doi.org/10.1007/s11104-010-0592-y
- A. J. Franzluebbers, S. F. Wright and J. A. Stuedemann, “Soil Aggregation and Glomalin under Pastures in the Southern Piedmont USA,” Soil Science Society of America Journal, Vol. 64, No. 3, 2000, pp. 1018-1026. http://dx.doi.org/10.2136/sssaj2000.6431018x
- E. R. Lutgen, D. Muir-Clairmont, J. Graham and M. C. Rillig, “Seasonality of Arbuscular Mycorrhizal Hyphae and Glomalin in a Western Montana Grassland,” Plant and Soil, Vol. 257, No. 1, 2003, pp. 71-83. http://dx.doi.org/10.1023/A:1026224209597
- M. C. Rillig, F. T. Maestre and L. J. Lamit, “Microsite Differences in Fungal Hyphal Length, Glomalin, and Soil Aggregate Stability in Semiarid Mediterranean Steppes,” Soil Biology and Biochemistry, Vol. 35, No. 9, 2003, pp. 1257-1260. http://dx.doi.org/10.1016/S0038-0717(03)00185-8
- K. Klumpp, S. Fontaine, E. Attard, X. Le Roux, G. Gleixner and J.-F. Soussana, “Grazing Triggers Soil Carbon Loss by Altering Plant Roots and Their Control on Soil Microbial Community,” Journal of Ecology, Vol. 97, No. 5, 2009, pp. 876-885. http://dx.doi.org/10.1111/j.1365-2745.2009.01549.x
- L. J. Ingram, P. D. Stahl, G. E. Schuman, J. S. Buyer, G. F. Vance, G. K. Ganjegunte, J. M. Welker and J. D. Derner, “Grazing Impacts on Soil Carbon and Microbial Communities in a Mixed-Grass Ecosystem,” Soil Science Society of America Journal, Vol. 72, No. 4, 2008, pp. 939-948. http://dx.doi.org/10.2136/sssaj2007.0038
- M. A. Liebig, J. R. Gross, S. L. Kronberg, R. L. Phillips and J. D. Hanson, “Grazing Management Contributions to Net Global Warming Potential: A Long-Term Evaluation in the Northern Great Plains,” Journal of Environmental Quality, Vol. 39, No. 3, 2010, pp. 799-809. http://dx.doi.org/10.2134/jeq2009.0272
- J. Six, S. D. Frey, R. K. Thiet and K. M. Batten, “Bacterial and Fungal Contributions to Carbon Sequestration in Agroecosystems,” Soil Science Society of America Journal, Vol. 70, No. 2, 2006, pp. 555-569. http://dx.doi.org/10.2136/sssaj2004.0347
- X. He, Y. Li and L. Zhao, “Dynamics of Arbuscular Mycorrhizal Fungi and Glomalin in the Rhizosphere Ofartemisia Ordosica Krasch. In Mu Us Sandland, China,” Soil Biology and Biochemistry, Vol. 42, No. 8, 2010, pp. 1313-1319. http://dx.doi.org/10.1016/j.soilbio.2010.03.022
- S. F. Wright, M. Franke-Snyder, J. B. Morton and A. Upadhyaya, “Time-Course Study and Partial Characterization of a Protein on Hyphae of Arbuscular Mycorrhizal Fungi During Active Colonization of Roots,” Plant and Soil, Vol. 181, No. 2, 1996, pp. 193-203. http://dx.doi.org/10.1007/BF00012053
- W. D. Kemper and R. C. Rosenau, “Aggregate Stability and Size Distribution,” In: Methods of Soil Analysis, Part I. Physical and Mineralogical Methods, American Society of Agronomy-Soil Science Society of America, Madison, WI, 1986, pp. 425-442.
- J. J. Halvorson and J. M. Gonzalez, “Tannic Acid Reduces Recovery of Water-Soluble Carbon and Nitrogen from Soil and Affects the Composition of Bradford-Reactive Soil Protein,” Soil Biology and Biochemistry, Vol. 40, No. 1, 2008, pp. 186-197. http://dx.doi.org/10.1016/j.soilbio.2007.07.022
- K. A. Nichols and S. F. Wright, “Comparison of Glomalin and Humic Acid in Eight Native U.S. Soils,” Soil Science, Vol. 170, No. 12, 2005, pp. 985-997. http://dx.doi.org/10.1097/01.ss.0000198618.06975.3c