O. BOCK, G. SCHMITZ
The present study scrutinizes two predictions of the pub-
lished model. First, practised and unpractised motor systems
are assigned by different weights, while practised and unprac-
tised sensory modalities are not; if so, adaptation should trans-
fer partially between arms but fully between targets from dif-
ferent modalities. It has indeed been shown that adaptation with
visual targets transfers to acoustic targets (Zwiers, van Opstal,
& Paige, 2003) and that aftereffects are equal in both modalities
(Kagerer & Contreras-Vidal, 2009); however the latter study
always tested acoustic targets first and visual targets thereafter,
so that a possible visual advantage might have dissipated by the
time of testing. To overcome this problem, we now test both
arms and modalities in counterbalanced order.
According to the second prediction of the published model,
both arms can concurrently adapt to different distortions with-
out interference, and this dual adaptation should again transfer
fully between sensory modalities. The first part of this predict-
tion has been confirmed in literature (Bock, Worringham, &
Thomas, 2005; Prablanc et al., 1975), and we now address the
second part.
Methods
25 male and 23 female subjects aged 22.0 1.6 years parti-
cipated after signing their informed consent to this study, which
was approved in advance by the first author’s Ethics Committee.
Subjects were right-handed, healthy, and had no prior experi-
ence with adaptation research. As shown by the inset of Figure
1, subjects faced an opaque horizontal panel with a wooden
dowel protruding downwards underneath their chin. Targets
were presented on the upper surface of the panel, along a semi-
circle of 36 cm radius around the dowel. They were presented
in balanced order at 30, 18, and 6 deg about straight-ahead,
either as light dots of 1.5 cm radius, or as sounds (mix of 0.45,
1.35, 2.30 and 3.20 kHz) from loudspeakers of 1.5 cm radius;
the loudspeaker array was hidden from view by a fabric screen.
Subjects pointed with their index fingertip underneath the panel,
moving from the dowel towards each target where the radial
response component was stopped by a semicircular barrier.
Subjects then moved the finger along that barrier until it was
aligned with the target. They couldn’t see their arm and hand
because of the opaque panel, but index fingertip position was
registered by the Fastrak® system (resolution 1 mm and 17 ms),
and could be displayed as a cursor on the upper surface of the
panel to provide real-time visual feedback.
The experiment was subdivided into pointing episodes of 45
s duration (or about 25 responses), separated by rest breaks of 5
s. Each experiment started with six baseline episodes. In the
first two, subjects pointed with their right hand at visual targets
and received veridical visual feedback. In the remaining four
baseline episodes, they pointed without visual feedback at vis-
ual targets with their right hand (VR), at visual targets with
their left hand (VL), at acoustic targets with their right hand
(AR), and at acoustic targets with their left hand (AL); the order
of these episodes was counterbalanced across subjects. In the
subsequent adaptation episodes, subjects pointed at visual tar-
gets under rotated visual feedback. One half of them formed
group “single”, which used their right hand under a +30 deg
rotated feedback. The other half formed group “dual”, which
alternately used their right hand under a +30 deg rotated feed-
back, and their left hand under a −30 deg rotated feedback.
Group “single” performed 20 and group “dual” 40 adaptation
episodes, i.e., there were 20 adaptation episodes per hand in
both groups. The experiment concluded with four aftereffect
episodes, which replicated VR, VL, AR and AL, again in coun-
terbalanced order.
The registered finger position data were analysed by an in-
teractive computer routine which determined the directional
error of each response, defined as angle between target and
cursor direction 166 ms after movement onset. This quantifies
the feedforward component of motor control without confound-
ing it with feedback-based error corrections, which emerge later
during the response. For graphical presentation and statistics,
we calculated the mean error of each adaptation episode minus
that of the second baseline episode, and the mean errors of each
aftereffect episode minus that of the pertinent baseline episode.
To facilitate comparisons, data yielded with the left hand of
group “dual” were sign-reversed. The outcome was submitted
to analyses of variance (ANOVAs) with Greenhouse-Geyser
corrections if variances were unequal.
Results
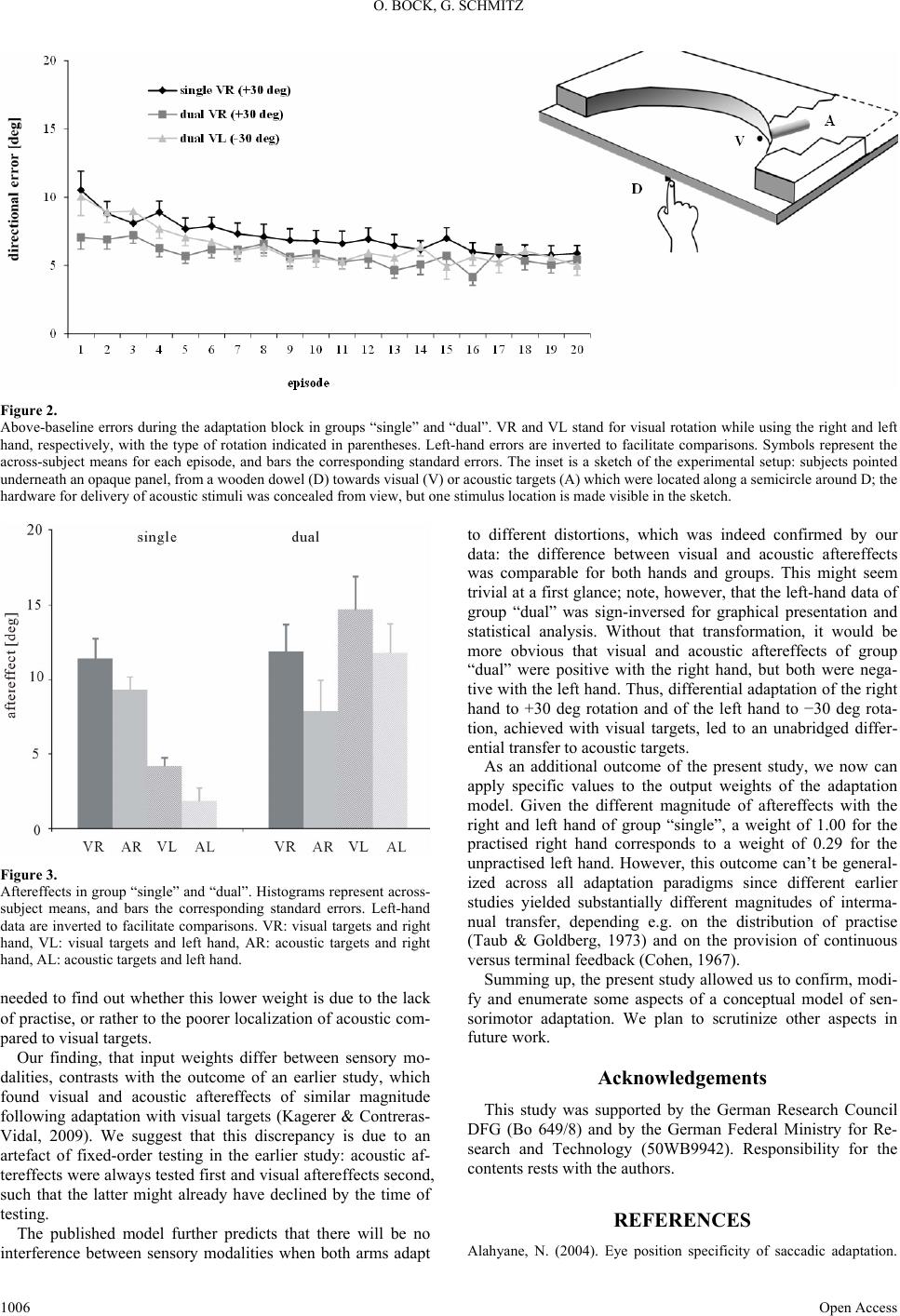
Figure 2 illustrates the errors of group “single” and “dual”
throughout the adaptation block. For clarity, the left hand of
group “dual” is plotted in same episodes as the right hand al-
though the data actually came from separate, interleaved epi-
sodes. ANOVA with the between-factor Task (single, dual right,
dual left) and the within-factor Episode (1, 2, …, 20), yielded
significance only for Episode (F(19,1292) = 13.15, p < 0.001).
The aftereffects of both groups are summarized in Figure 3.
ANOVA with the between-factor Group (single, dual) and the
within-factors Hand (R, L) and Modality (V, A) yielded signi-
ficance for Group (F(1,46) = 5.83; p < 0.05), Hand (F(1,46) =
8.46; p < 0.01) and Group*Hand (F(1,46) = 64.80; p < 0.001):
aftereffects with the right hand reached a similar magnitude in
both groups, but only 29% of that magnitude with the unprac-
tised left hand of group “single”; the practised left hand of
group “dual” even showed slightly larger aftereffects than the
right hand. Furthermore, ANOVA yielded significance for
Modality (F(1,46) = 15.39; p < 0.001): aftereffects with acous-
tic targets were only 66% of those with visual targets. No other
interactions reached significance.
Discussion
The present study evaluates the transfer of sensorimotor ad-
aptation to an unpractised limb and to an unpractised sensory
modality, to scrutinize a recently published conceptual model
(Bock, 2013). Our discussion will focus on the observed after-
effects rather than on the time-course of adaptation, since after-
effects are thought to be a pure indicator of sensorimotor re-
calibration while the time-course of adaptation may also reflect
workaround strategies (McNay & Willingham, 1998; Redding,
1996; Werner & Bock, 2007).
The above model predicts that inputs from different sensory
modalities are weighted equally, which implies that adaptation
will fully transfer between modalities. This, however, was not
the case in our study: following adaptation with visual targets,
the magnitude of aftereffects with acoustic targets was only
66% of that with visual ones. The model therefore must be
amended; specifically, if the practised visual modality in our
study is given a weight of 1.00, then the unpractised acoustic
modality should obtain a weight of only 0.66. Further work is
Open Access 1005