American Journal of Climate Change
Vol.07 No.02(2018), Article ID:84917,16 pages
10.4236/ajcc.2018.72012
Variation in Expression of Sub1 Gene and Association with Submergence Stress Related Traits in Advance Rice ( Oryza sativa L.) Genotypes as a Tool of Climate Change Adaptation
Zulqarnain Haider1, Muhammad Akhter1, Abid Mahmood2, Usman Saleem1
1Rice Research Institute, Kala Shah Kaku, Lahore, Pakistan
2Ayub Agricultural Research Institute, Faisalabad, Pakistan

Copyright © 2018 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/



Received: December 18, 2017; Accepted: May 27, 2018; Published: May 30, 2018
ABSTRACT
Floods have now become most detrimental natural catastrophe worldwide due to radical climatic fluxes. Therefore, there is a dire necessity to develop a high yielding rice lines to deal with this scenario. For this purpose, a large scale experiment was conducted including one hundred and fifteen (115) rice genotypes having SUB1 gene imported from International Rice Research Institute (IRRI) Philippines, six local cultivars/approved varieties and three high yielding rice varieties i.e. Sabitri, IR6 and NSICRC222 being used as potential varieties in different countries of Asia as susceptible check and IR64-SUB1 as tolerant check. The genotypic screening was performed using two PCR-based DNA markers i.e. ART5 and SC3. Phenotypic screening was conducted in a natural pond to assess the interaction of SUB1 gene in natural stagnant flood water as well as the suitability of introgression of SUB1 gene into approved varieties and elite rice lines. The genotypes were assessed in terms of plant survival percentage, submergence tolerance index, physical condition, stem elongation, number of grains per panicle, thousand grain weight, grain yields and deviations in these traits after submergence stress. The PCR results suggested that both the primers ART5 and SC3 may be used as potential PCR-based markers for molecular screening of rice genotypes for SUB1 QTL. Furthermore, it confirmed the presence of SUB1 gene in all the lines imported from IRRI, while it was absent in all the local cultivars studied. All the genotypes with submergence tolerant gene (SUB1) showed significantly greater tolerance level in submergence stress of 14 days, as compared to other local cultivars/varieties, authenticating the effectiveness of SUB1QTL in conferring submergence tolerance. Significantly different performances of all the SUB1 genotypes in terms of all the studied traits indicate high Genotypic and Genotypic Environment Interaction (GEI) of SUB1QTL. Employment of SUB1 lines such as R105479:149-18, IR64-SUB1 and Rl05469:81-22-3 in breeding programs for developing flood tolerant rice varieties might further upsurge rice yields in flash flood areas. Correlation analysis revealed that plant survival percentage after submergence, reduced stem elongation during submergence and submergence tolerance index are very important traits for developing submergence tolerant lines.
Keywords:
Oryza sativa L., Sub1 QTL, Floods, Climate Change, Phenotypic Expression, Association Analysis, Stress Related Traits

1. Introduction
Climate change is termed as variation in statistical distribution of weather patterns (temperature variations, rainfall, light intensity etc.) when that change persists for a prolonged duration. Erratic fluctuations in rainfall and other types of precipitation will be one of the most perilous aspects determining the overall sway of climate change scenario. It is also predicted that some regions will be prone to extreme drought while already wet regions will be wetter due to extreme rainfalls causing flooding.
World population is increasing tremendously and so is the case with food demand. Rice is basic food item of nearly 50% of the world population. This demand necessitates a significant enhancement of rice production even in marginal-farmlands of the world. Summer monsoon rains damage rice crops drastically every year in terms of yield and grain quality [1]. Out of total area under rice, more than 1.6 million hectares have been affected by excessive rains and floods [2]. In Pakistan, approximately 70% rice farms are prone to seasonal flooding. Rice crop may be the only crop which can effectively be grown in flood prone ecosystems [3]. These floods are sometimes so damaging that rice farmers in some parts of the country lose their entire rice crop. According to annual flood reports of Pakistan, Punjab’s paddy production had suffered an estimated loss of 217 thousand tons due to seasonal floods during 2014-15 and as a result overall national paddy production remained below the target of previous year. According to Toojinda et al. [4] , flooding is a serious constraint to rice crop and its survival in rainfed lowland and deep-water areas. Recently, extreme weather conditions and unpredicted heavy rains due to drastic climatic changes in Pakistan have resulted increased extent of submergence during rice season [1] [2]. Major rice-damaged districts are Hafizabad, Jhang, Gujranwala and Chiniot that are major rice growing areas of Pakistan. The experts say that the situation may become worse as climate change progresses. Flash floods, in which muddy water recedes in three to seven days, prohibit the sunlight to reach plants resulting in termination of photosynthesis, food making process in plants. Low oxygen level in deep floods also causes submerged plants to suffocate without oxygen. This rapid decline in the oxygen (~10,000-fold less) during prolonged submergence in accompanied by drastic lessening in cellular O2 levels and an energy crisis, becomes severe under absence of photosynthesis [5] [6]. All this results in severe damages to rice crop in terms of yield loss as well as quality deterioration.
Therefore, submergence tolerance has become an important breeding objective in order to reduce yield losses [7] [8] [9]. Recently, in Pakistan, all the rice cultivars being cultivated are much sensitive to flooding and anaerobic water-logged situation. With the new developments, it has become possible to cope with such flooded and water-logged situation by incorporating genes such as SUBMERGENCE 1 (SUB1), a robust quantitative trait locus from a submergence tolerant FR13A landrace, into local high yielding cultivars using Marker-assisted Breeding (MAB). Although, rice (Oryza sativa L.) is considered a flood-tolerant crop, its level of tolerance differs significantly and only limited cultivars show tolerance to prolonged submergence, most of them with SUB1 QTL [10]. SUB1 is a variable polygenic locus that encodes two to three ethylene responsive factors (ERF) DNA binding proteins [11]. However, the response of this SUB1 QTL differs with the genotypes and other environmental factors [3]. In order to study these responses, an experiment was organized to screen that included 115 rice genotypes with SUB1 QTL from International Rice Research Institute (IRRI), Philippines along with six local high yielding rice cultivars being cultivated in Pakistan.
2. Material and Methods
1) Phenotypic Screening
The study was carried out at the experimental fields of the Rice Research Institute (RRI), Kala Shah Kaku Pakistan during rice (Kharif) season of 2015. One hundred and fifteen (115) rice genotypes having SUB1 genes imported from International Rice Research Institute (IRRI) Philippines, six (06) local cultivars/released varieties and three high yielding varieties i.e. Sabitri, IR6 and NSICRC222 being used as potential variety in many countries of Asia, were used as experimental material. All these genotypes were evaluated in a natural water pond that allows maintenance of water depth of 150 cm for a period of 15 days. The experiment was laid out in a triplicated randomized complete block design (RCBD). Seeds were sown on the wet raised beds and 21 days seedlings were transplanted to well puddle soil in the deep pond/submergence tank at 20 cm × 20 cm with two seedlings per hill. Plot size was maintained in two rows of 5 m2 length for each entry. Non-experimental lines of Super Basmati were also planted on each side of the pond to observe the damages due to submergence. Nitrogen fertilizer was applied at 30:30:30 kg/ha as basal dose a day before starting transplanting. Gap-filling was also practice as required 7 days after transplanting (DAT) in order to ensure 100% plant establishment. Plant was counted before applying submergence stress. The transplanted seedlings were allowed to establish their roots for 20 DAT before submergence. The pond was filled with normal canal water up to depth level of 1500 cm so that all the plants of all the entries were completely submerged within a day. The same water depth was maintained for the period of 14 days by adding water regularly. After completing 14 days of complete submergence, the treatment was terminated. The water was drained out of pond to water level of 1 - 2 cm. After a day, the water was again filled up to a level of 5 cm. Second split of nitrogen fertilizer was applied at the rate of 30 kg/ha after 10 days and the third split of 30 kg/ha was applied after 20 days of second split. All the following agronomic and plant protection measures were adopted as normal in order to allow the plant to recover from submergence stress.
Another experiment was also conducted, at parallel with the submergence experiment, in order to ascertain the percentage yield losses along with other yield components. All the entries were laid out in triplicated RCB design. The same nursery was used for this experiment. Two seedlings were transplanted per hill in order to maintain plant population with 20 cm × 20 cm plot size of each entry. Inorganic fertilizer was applied in two splits, a basal doze of 200 kg/ha before transplanting using NPK (15:15:15 kg/ha) and top dressed with Urea fertilizer @ 65 kg/ha and 35 kg/ha at tillering and booting stage respectively. For this experiment, 5 cm water depth was maintained with normal irrigations. Weeds were controlled by applying post-emergence herbicides as required.
Data for number of plants before and after submergence treatment, plant height, number of grains per panicle, 1000 grain weight, number of tillers, yield per plant were recorded for all the entries in both treated (submergence tank) as well as normal (irrigated field) experiments. Then the percentage loss of yield was calculated using the formula as given below:
where
= Percentage yield loss
= Yield under normal irrigated condition
= Yield under submergence stress
Similarly, percentage survival after submergence stress was calculated by the following formula also used by Ranawake et al., 2014 [12]
where
= Number of plants before submergence stress
= Number of plants after submergence stress
Stem elongation/reduction or height gain/loss of stressed plants was calculated by the following formula:
where
= Average height of plants under normal condition/non-stress environment
= Average height of plants in submergence tank after water submergence stress
= Stem elongation/reduction after submergence stress
2) Molecular Screening
The molecular screening was conducted in Marker Assisted Selection (MAS) lab of Rice Research Institute, Kala Shah Kaku. The objective of the experiment was to confirm the presence or absence of SUB1gene in 128 rice genotypes. Grains of all the genotypes were grown in germination trays under controlled environmental conditions (temperature 30˚C - 32˚C and humidity 75% - 80%) in an automated growth chamber. Seedlings were taken for DNA extraction using Miniprep protocol by Mackill [13]. In order to examine the quality of extracted genomic DNA (3 µL) mixed with 6x loading dye (2 µL) was loaded in 1% agarose gel.
Polymerase chain reaction (PCR) was executed [13] using total reaction mixture of 10 µL (15 ng of DNA template, 1 µL 10x TB buffer, 1 µL of 1 mM dNTP, 0.50 µL each of 5 µM forward and reverse primers (SC3 and ART5) and 0.25 µL of Taq DNA polymerase (4 U/µL) using Qantarus (Q-cycler 96 model) thermocycler. Initial denaturation was completed after 5 minutes at 94˚C, each cycle was programmed for 1-minute denaturation at 94˚C, 1-minute annealing at 55˚C, and 2-minute extension at 72˚C. After completing 35 repeated cycles, final elongation was programmed for 5 minutes at 72˚C. The final PCR products were mixed with loading dye (bromophenol blue) and were evaluated using vertical electrophoresis (8% polyacrylamide gel). The gels were visualized using Gel Documentation System (UVI-tech.) after staining in 0.5 mg/ml ethidium bromide. The data were scored for SUB1 gene using “++/−−” signs for presence/absence respectively (Table 1).
3) Statistical analyses
Statistical analyses were conducted using Statistical software STATISTIX (v. 8) and Microsoft Excel (v. 2016).
3. Results and Discussion
1) Molecular screening
Results of PCR revealed that ART5 which is a closely linked marker in the promoter region of SUB1C a transcription-factor gene [3] showed bands in some of the genotypes while the presence of this marker was nullified in all the rest of the genotypes used in this study. Like-wise, SC3 which is a closely linked marker with SUB1A showed polymorphism in almost all the genotypes. Only

Table 1. Reverse and forward sequences of primers used for SC3 and ART5 to investigate sub1 gene in rice cultivars.
few genotypes showed distinct bands in case of both the markers. Therefore, concluding these results it may be suggested that both the markers i.e. ART5 and SC3 may be used as potential PCR-based markers for molecular screening of rice genotypes for SUB1 QTL [3] [11]. Genotyping of all the lines imported from IRRI confirmed the presence of SUB1gene. Furthermore, both the primers did not show polymorphism in other local cultivars as well as exotic cultivars.
2) Phenotypic screening in submergence tank
The obtained results clearly indicated that all the genotypes with submergence tolerant gene (SUB1) showed significantly greater tolerance level as compared to other local cultivars/varieties, authenticating the effectiveness of SUB1QTL in conferring submergence tolerance [14]. However, a significant variation was observed among the studied cultivars in tolerance for submergence as well as all other traits studied.
a) Physical condition after treatment
Irradiance due to turbidity and limited CO2 supply are two major factors for tissue death in case of stagnant floodwater submergence. As a result of these factors, rice plants become unable to photosynthesize and continue to maintain supply of carbohydrate for respiration and alcoholic fermentation [15]. Therefore, data for physical condition of the plants of each genotype were collected after submergence stress in terms of tissue death due to prolonged condition of submergence stress. Comparatively healthiest plants were given 5 ranking while the plants having poorest health condition were given 0 ranking and so on. Results clearly showed that genotypes without SUB1gene showed highest rate of tissue death as compared to genotypes with SUB1gene. Furthermore, the SUB1genotypes also showed variation in expressivity of the SUB1gene. Five genotypes showed 5 ranking (very good) in physical condition after submergence stress, thirty one showed 4 (good), Nineteen showed 3 (fair) while the rest showed <3 (below average and poor) ranking. Only five plants that maintained their health status and showed very good health condition after de-submergence will be considered for being used as donor parents (Figure 1).
b) Plant survival percentage
The data for plant survival percentage was collected 60 days after submergence treatment when submergence susceptible checks had completely dried. Plant survival percentage varied from 0% (all plants dried) to 100% (all plants

Figure 1. Distribution pattern of physical condition of genotypes after de-submergence (14 days of submergence period) shown by 4th order polynomial trend lines.
were green after submergence treatment) survival. Considering IR64-SUB1line (showing 90.6% survival) as submergence tolerant while IR6, NSICRC-222 and Sabitri (showing survival percentage of 21.9%, 18.8% - 21.9% and 9.4% respectively) as submergence susceptible lines, criterion for evaluating the lines for submergence tolerance rice lines may be designed. It can be perceived from Figure 2 that the entries showing above 80% survival percentage can be presumed as submergence tolerant, while below 20% as submergence susceptible lines. Results indicated that all the studied lines with submergence tolerance gene (SUB1) has significant higher survival rate than other varieties under evaluation [14]. All the local cultivars could not survive at all.
c) Submergence Tolerance Index
Similarly, Submergence Tolerance Index (STI) was calculated in order to measure the level of tolerance to submergence stress. Submergence tolerant lines showed above 8.0 index while the susceptible lines had less than 2.0 tolerance index value. IR64-SUB1 (IRRI variety) showed STI value of 0.906 indicating that it is a submergence tolerant line whereas IR6, NSICRS222 and Sabitri, with no SUB1 QTL showed STI value of 0.219, 0.188 and 0.094 respectively. All the local approved varieties of Pakistan showed zero tolerance level as indicated by the lowest STI values. Further, the lines with SUB1QTL (submergence tolerant gene) showed significantly higher STI value as compared to local cultivars deprive of the gene. It was further observed that there was also a significant variability among lines with SUB1gene. This reconfirms the previous findings that the flood-tolerant SUB1 gene, when transferred into popular farmer-preferred rice varieties, enables them to survive even after two weeks of total submergence [13] [16]. Radar graph (Figure 3) shows the performance of the genotypes under submerged condition of 14 days in terms of Tolerance Index Values from highest to lowest. It shows that IR64-SUB1 (Tolerance check) line earns 17th position

Figure 2. Distribution pattern of survival percentage of genotypes after de-submergence (submergence duration of 14 days).

Figure 3. Radar graph showing performance of genotypes in terms of tolerance index (TI) values.
with TI value of 0.906. Among all, 18 lines showed TI value above 0.90 (Highly tolerant), 13 lines more than 0.80 TI values (Moderately Tolerant), 13 lines between 0.60 to 0.80 TI values (Moderately Susceptible) lines, and rest of the lines showed highly susceptible performance with TI values below 0.6.likewise among the susceptible checks (without SUB1gene), IR6 got the 85th position, both NSICRC222 lines got 86th and 87th positions, both lines of Sabitri got 103rd and 104th position. All the local cultivars poorly performed in submerged condition with TI values below 0.2 indicating that there is no tolerance to submergence and there is dire need to transfer SUB1QTL into our local high yielding cultivars in order to cope with growing frequency of floods in imminent era.
d) Stem elongation
Findings clearly indicated that almost all the studied genotypes showed more stem elongation in submerged condition as compared to those in normal condition (Table 2). Under normal condition, plants showed average height of 105 cm, minimum and maximum observed plant height was 84 and 132 respectively. As compared to this, plants showed average height of 121 cm, minimum and maximum observed plant height was 100 and 151 respectively under submergence stress. The plants that showed higher stem elongation, also showed less yield due to rapid consumption of reserved carbohydrates. Fragile stem with extra height made the plants more prone to lodging. On the basis of this information, only 45 plants that showed less than 10 cm elongation in submergence stress were selected for further use in breeding programs (Figure 4). The same response of rice plants was also observed by Ranawake et al. [12] , Redona and Mackill [17] and Ismail et al. [18]. They also observed that submergence stress significantly promotes stem elongation in young rice seedlings. Won and Yoshida [19] observed in a study that enhanced stem elongation in submerged condition is an adaptive reaction of water-seeded rice for attaining more oxygen. However, the genotypes with SUB1QTL elongated less as compared to those without SUB1QTL. Moreover, among the SUB1 genotypes, there was also a significant difference in stem elongation under submergence stress. This is due to the blocking mechanism of SUB1gene to express Ethylene Responsive Factors (ERF) which are genetically produced under submerged condition as a defensive strategy and these ERFs are responsible for stem elongation in rice [12].
e) Number of tillers
There was no significant change in tillering ability of studied genotypes due to submergence stress. The number of tillers ranged from 10 to 24 tillers per plant with the variety IR105469:43-51-16 showing the highest number of tillers of 24 while the lowest number of tillers (10 tillers) was recorded for variety IR102885-1666-11-3-11.
f) Number of grains per panicle
The genotypes showed significant difference for number of grains per panicle. It varied from 45 grains in IR105463:24-13-12to 171 in IR102885-1666-7-1 -12, followed by R102885-1653-1-1-19, IR105469:72-17-21 and Sabitri with 149, 149 and 143 grains per panicle under normal/control condition. On the other hand, Rl05463:16-14-3, IR102885-1651-7-12 and R105462:4-2-22 developed minimum 0, 0 and 1 grains under stress while maximum grains per panicle were observed in IR105495:10-12, R102885-1666-11-3-14 and IR105479:82-11 with 96, 91 and 88 grains per panicle respectively. The study showed that grains per panicle reduced significantly due to submergence stress.
g) Thousand grain weight
Genotypes also showed significant difference in thousand grain weight among themselves as well as between the two environments. The TGW ranged from 17.31 g to 37.89 g under normal while 2.98 g to 36.28 g under stressed condition. Average TGW was calculated as 23.24 g and 18.79 g under normal and stressed condition respectively.
Table 2. Average (±S.E), Minimum and Maximum values of different traits of genotypes under normal and submerged conditions.

Figure 4. Distribution pattern for Stem Elongation of 128 genotypes after submergence of 14 days.
h) Yield
The genotypes showed significant difference for grain yield within and between both the conditions. Under control, average yield was observed as 4.9 t/ha and it varied from 1.0 t/ha to 11.1 t/ha, while under stressed condition average yield was 2.54 t/ha and it ranged from 0.089 t/ha to 8.0 t/ha. Graph 1(a) and Graph 1(b) clearly show the performance of genotypes under control and submerged condition respectively. Results show that genotypes IR105479:149-18 showed the maximum yield (11151 kg/ha), followed by IR105469:43-51-16 (9039 kg/ha), IR102885-1642-7-6 (8790 kg/ha) and NSICRC222 (8381 kg/ha) under normal condition. Whereas, under stress R105479:149-18 was the highest yielder (8000 kg/ha), followed by IR64-SUB1 (6034 kg/ha) and Rl05469:81-22-3 (5800 kg/ha) as depicted in Graph 1(a) and Graph 1(b). These results suggest that genotype IR105479:149-18 may be used as potential donor parent for local environment as it performed best in both normal as well as under stressed condition. IR64-SUB1 may also be used as donor parent in breeding programs for developing high yielding submergence tolerant rice varieties.
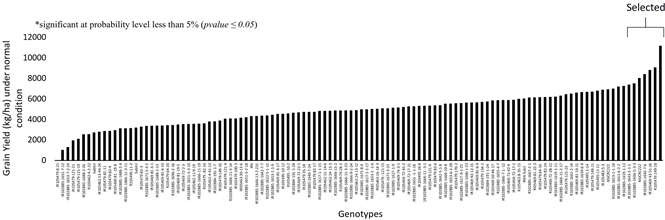
(a)
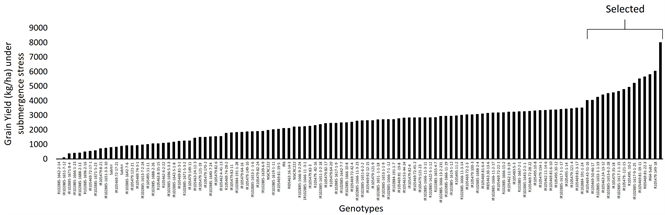
(b)
Graph 1. (a) Yield (kg/ha) performance of all the studied genotypes under normal condition; (b) Yield (kg/ha) performance of all the studied genotypes under submergence stress of 14 days.
i) Correlation Analysis:
Results of correlation analysis (Table 3) suggested that yield reduction after submergence stress was positively correlated with grain yield under normal condition (0.6668**), plant height under stress (0.6567**), stem elongation due to stress (0.6840**) and reduction in thousand grain weight (0.2723*), while negatively correlated with yield under submergence stress (−0.4705**). Yield under normal or control condition showed positive correlation with yield under stress (0.3439*), plant height under stress (0.3553**) and Stem elongation after stress (0.3331**). Grain yield under stress condition showed positive correlation with thousand grain weight under normal (0.2914**) and under stress (0.3333**) while negative correlation with plant height under stress condition (−0.4068**). Stem elongation was negatively correlated with plant height under normal (−0.3224**) and positively correlated with plant height under stress (0.6471**). Reduction in thousand grain weight was positively correlated with plant height under stress (0.3195*). Same results are also depicted in Figures 5(a)-(h).
It can be suggested from the findings of correlation analysis that reduction in yield after submergence stress was mainly attributed to stem elongation as depicted

Figure 5. Results of correlation analysis among yield and stress related traits in rice observed under flood conditions.
Table 3. Correlation (Pearson) analysis of all the studied traits of 128 genotypes under normal and natural submerged condition (TGW = Thousand Grain Wight, GPP = Grains per panicle, Y = Yield, PH = Plant Height, SE = Stem Elongation; n = normal condition, s = submerged condition) ɼ = correlation coefficient; p = probability.
in Figure 5(b). Figure 5(e) and Figure 5(f). The genotypes that showed higher stem elongation in stress were less yielder under stress. All the local cultivars elongated rapidly under stress and lead to lodging after de-submergence. However, genotypes with SUB1 gene elongated less and showed better performance. Furthermore, the expression of SUB1 QTL was also significantly variable resulting in a range in values of yield, yield reduction and elongation ability after stress. This suggests the high interaction of SUB1 QTL with the genome as well as with the environment. Elongation was observed more in those genotypes that were already taller as compared to shorter plants. Moreover, genotypes with better yield performance in control were also better yielders in stress condition. However, reduction in yield was observed more in genotypes that were very high yielding under normal condition. Therefore, it may be concluded that while selecting the genotypes for submergence stress, plant height may be considered at priority.
4. Conclusion
The results clearly showed that all approved rice varieties are sensitive to submergence stress, elucidating the urgent need for flood/submergence tolerant cultivars of rice. Approved rice varieties couldn’t survive at all after two weeks of complete submergence in water. The PCR results suggested that both the markers ART5 and SC3 may be used as potential PCR-based markers for molecular screening of rice genotypes for SUB1 QTL. Furthermore, it confirmed the presence of SUB1 gene in all the lines imported from IRRI, while it was absent in all the local cultivars studied. All the genotypes with submergence tolerant gene (SUB1) showed significantly greater tolerance level in submergence stress of 14 days, as compared to other local cultivars/varieties, authenticating the effectiveness of SUB1 in conferring submergence tolerance. Significantly different performances of all the SUB1 genotypes in terms of all the studied traits indicate high Genotypic and Genotypic Environment Interaction (GEI) of SUB1QTL. Correlation analysis revealed that plant survival percentage after submergence, reduced stem elongation during submergence and submergence tolerance index are very important traits for developing submergence tolerant lines. Employment of SUB1 lines such as R105479:149-18, IR64-SUB1 and Rl05469:81-22-3 in breeding programs for developing flood tolerant rice varieties might further increase yields in flash flood areas. This screened material can be used as a new cultivar or as parent material to breed new flood-tolerant high yielding local rice varieties to be grown under water-logged and flood prone areas [20].
Acknowledgements
The genetic material used in this study was imported from International Rice Research Institute (IRRI), Philippines, to be used in different breeding programs under Agriculture Innovation Program (AIP) Philippines. The same material was used in this investigation for developing flood tolerant lines of rice and the sole purpose of this investigation is the alleviation of livelihood of farmers living in areas prone to frequent floods and coping with the climate change scenario in future.
Cite this paper
Haider, Z., Akhter, M., Mahmood, A. and Saleem, U. (2018) Variation in Expression of Sub1 Gene and Association with Submergence Stress Related Traits in Advance Rice (Oryza sativa L.) Genotypes as a Tool of Climate Change Adaptation. American Journal of Climate Change, 7, 171-186. https://doi.org/10.4236/ajcc.2018.72012
References
- 1. Disaster Management Committee (DMC) (2002) Economic Analysis of Reclamation Measures for Salt Affected and Waterlogged Soils. Development and Management Consultants, Lahore, Pakistan 112-125.
- 2. Pakistan Economic Survey (PES) (2016) Pakistan Bureau of Statistics (PBS), Ministry of Finance, Government of Pakistan, Agric., 28-29.
- 3. Sarkar, R.K. and Bhattacharjee, B. (2011) Rice Genotypes with SUB1 QTL Differ in Submergence Tolerance, Elongation Ability during Submergence and Re-Generation Growth at Re-Emergence. Rice, 5, 2-11.
- 4. Toojinda, T., Siangliw, M., Tragoonrung, S. and Vanavichit, A. (2003) Molecular Genetics of Submergence Tolerance in Rice: QTL Analysis of Key Traits. Annals of Botany, 91, 243-253. https://doi.org/10.1093/aob/mcf072
- 5. Bailey, S.J. and Voesenek, L.A.C.J. (2008) Flooding Stress: Acclimations and Genetic Diversity. Annual Review of Plant Biology, 59, 313-339. https://doi.org/10.1146/annurev.arplant.59.032607.092752
- 6. Licausi, F. and Perata, P. (2009) Low Oxygen Signaling and Tolerance in Plants. Advances in Botanical Research, 50, 139-198. https://doi.org/10.1016/S0065-2296(08)00804-5
- 7. Kawano, N., Ella, E., Ito, O., Yamauchi, Y. and Tanaka, K. (2002) Metabolic Changes in Rice Seedlings with Different Submergence Tolerance after De-Submergence. Environmental and Experimental Botany, 47, 195-203. https://doi.org/10.1016/S0098-8472(01)00126-5
- 8. Mackill, D.J. (1986) Progress in Rainfed Lowland Rice. Vol. 2, International Rice Research Institute, Rainfed Lowland Rice Improvement in South and South East Asia, Results of a Survey, Los Baños, 115-144.
- 9. Mohanty, H.K. and Chaudhary, R.C. (1986) Progress in Rainfed Lowland Rice. International Rice Research Institute, Breeding for Submergence Tolerance in Rice in India, Los Baños, 192-200.
- 10. Niroula, R.K., Pucciariello, C., Ho, V.T., Novi, G., Fukao, T. and Perata, P. (2012) SUB1-Dependant and -Independent Mechanisms Are Involved in the Flooding Tolerance of Wild Rice Species. The Plant Journal, 72, 282-293. https://doi.org/10.1111/j.1365-313X.2012.05078.x
- 11. Julia, B.S., Fukao, T., Ronald, P., Ismail, A., Heuer, S. and Mackill, D. (2010) Submergence Tolerant Rice: SUB1’s Journey from Landrace to Modern Cultivar. Rice, 3, 138-147. https://doi.org/10.1007/s12284-010-9048-5
- 12. Ranawake, A.L., Samarasinghe, U.G. and Senanayake, S.G.J.N. (2014) Submergence Tolerance of Some Modern Rice Cultivars at Seedling and Vegetative Stages. Journal of Crop and Weed, 10, 240-247.
- 13. Mackill, D.J. (2006) Breeding for Resistance to Abiotic Stresses in Rice: The Value of Quantitative Trait Loci. In: Lamkey, K.R. and Lee, M., Eds., Plant Breeding: The Arnel R Hallauer International Symposium, Blackwell Publishing, Ames, 201-212. https://doi.org/10.1002/9780470752708.ch14
- 14. Akinwale, M.G., Akinyele, B.O., Odiyi, A.C., Nwilene, F., Gregorio, G. and Oyetunji, O.E. (2012) Phenotypic Screening of Nigerian Rainfed Lowland Mega Rice Varieties for Submergence Tolerance. Proceedings of the World Congress on Engineering, 1, 4-6.
- 15. Setter, T.L., Ellis, M., Laureles, E.V., Ella, E.S., Semdhira, D., Mishra, S.B., Sarkarung, S. and Datta, S. (2016) Physiology and Genetics of Submergence Tolerance in Rice. Annals of Botany, 79, 67-77.
- 16. Xu, K.N., Deb, R. and Mackill, D.J. (2004) A Microsatellite Marker and a Codominant PCR-Based Marker for Marker-Assisted Selection of Submergence Tolerance in Rice. Crop Science, 44, 248-253. https://doi.org/10.2135/cropsci2004.2480
- 17. Redona, E.D. and Mackill, D.J. (1996) Genetic Variation for Seedling-Vigor Traits in Rice. Crop Science, 36, 285-290. https://doi.org/10.2135/cropsci1996.0011183X003600020012x
- 18. Ismail, A.M., Ella, E.S., Vergara, G.S. and Mackill, D.J. (2009) Mechanisms Associated with Tolerance to Flooding during Germination and Early Seedling Growth in Rice (Oryza sativa). Annals of Botany, 103, 197. https://doi.org/10.1093/aob/mcn211
- 19. Won, J.G. and Yoshida, T. (2000) Screening Cultivars at Low Dissolved Oxygen Level for Water Seeded Rice. Plant Production Science, 3, 112-132. https://doi.org/10.1626/pps.3.112
- 20. Zaman, S.B. and Ahmad, S. (2009) Salinity and Waterlogging in the Indus Basin of Pakistan: Economic Loss to Agricultural Economy. Managing Natural Resources for Sustaining Future Agriculture, 1, 4-8.