J. M. MENTER ET AL. 161
quences. Although the consequences of these interactions
are not well known with respect to keloids, one might
gain insight by considering the closely-related case of
systemic sclerosis, where imbalances in NO metabolism,
type I collagen expression, and the occurrence of nitrated
proteins have been known to occur for certain disease
states [24-26]. Melanin sequestration of NO and its role
in generating toxic NOx might have similar effects on
normal fibroblasts that could possibly result in their
transformation to keloid fibroblasts. This latter question
is currently under investigation in our laboratory (Nok-
kaew et al., in progress).
5. Conclusions
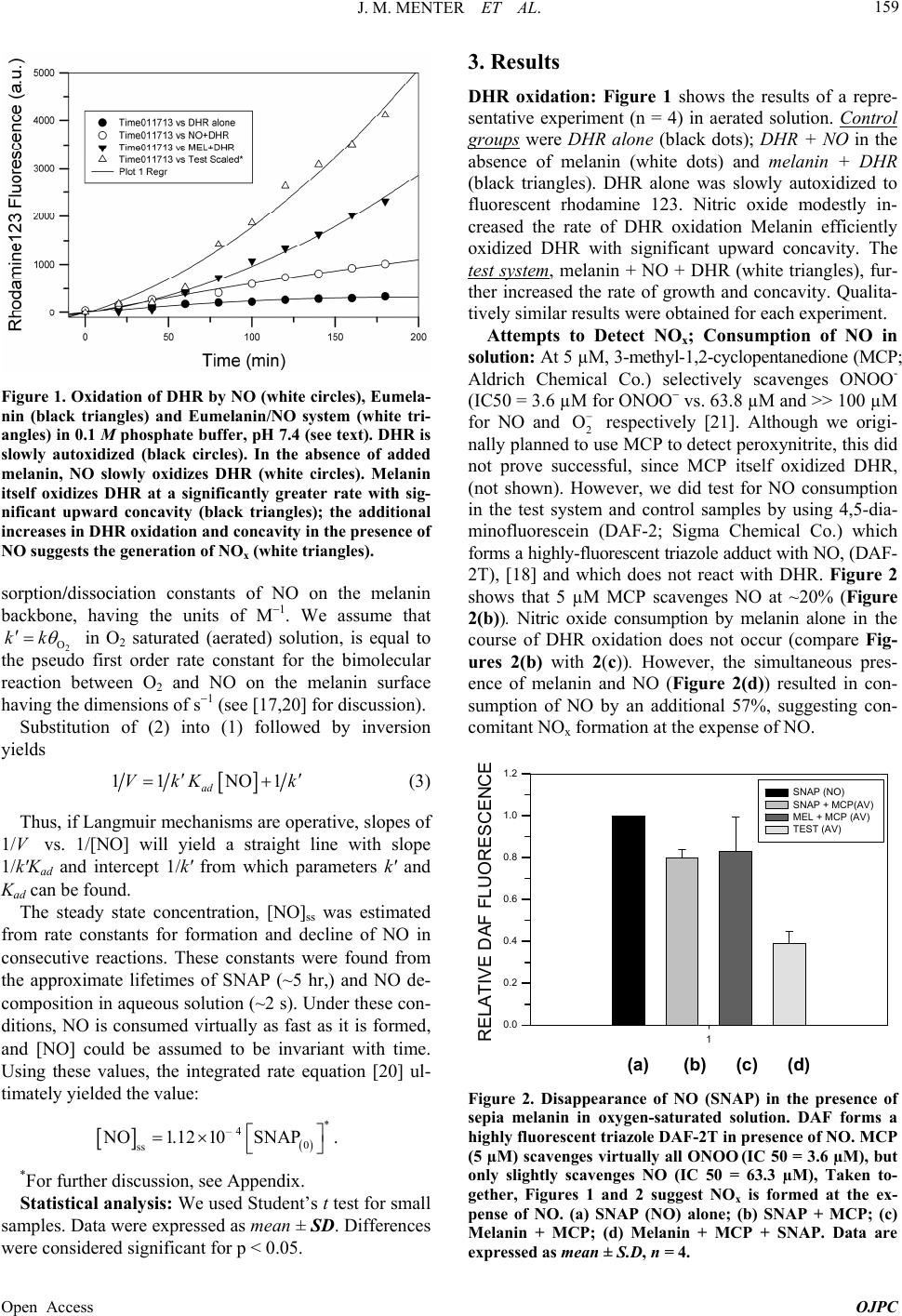
a) Sepia melanin accelerates (couples) the oxidation of
NO to NOx and the reduction of molecular O2 in buffered
aqueous solution.
b) The reaction kinetics fit well to a simple Langmuir
adsorption isotherm, and the reaction therefore appears to
take place between adsorbed NO and O2 via a chemi-
sorption mechanism. We obtained a value of pseudo first
order constant 7
8.2 10 sk1
and adsorption
coefficient Kad = 4.04 M1.
c) Such a reaction occurring in situ in dermal tissue
might have significant epigenetic consequences in keloid
formation or related pathologies.
6. Acknowledgements
This work was funded in part by GRANTS: MBRS
#GM08248, RCMI #8G12MD007602, and DOD # 911
NF-10-1 0448. There are no conflicts of interest.
REFERENCES
[1] L. Hong and J. D. Simon, “Current Understanding of the
Binding Sites, Capacity, Affinity, and Biological Signi-
ficance of Metals in Melanin” The Journal of Physical
Chemistry B, Vol. 111, No. 8, 2008, pp. 7938-7947.
http://dx.doi.org/10.1021/jp071439h
[2] W. Korytowski, P. Hintz, R. C. Sealy and B. Kalyanara-
man, “Mechanism of Dismutation of Superoxide Produc-
ed during Autoxidation of Melanin Pigments,” Bioche-
mical and Biophysical Research Communications, Vol.
131, No. 2, 1985, pp. 659-665.
http://dx.doi.org/10.1016/0006-291X(85)91288-4
[3] W. Korytowski, B. Pilas, T. Sarna and B. Kalyanaraman,
“Photoinduced Generation of Hydrogen Peroxide and Hy-
droxyl Radicals in Melanins,” Photochemistry and Photo-
biology, Vol. 45, No. 2, 1987, pp. 185-190.
http://dx.doi.org/10.1111/j.1751-1097.1987.tb05362.x
[4] J. M. Menter, D. Eatman, M. Bayorh, A. M. Dawaghreh
and I. Willis, “Pigment Melanin Scavenges Nitric Oxide
in Vitro: Possible Relevance to Keloid Formation,” Re-
search Letters in Physical Chemistry, Vol. 2008, 2008,
Article ID: 210616.
http://dx.doi.org/10.1155/2008/210616
[5] M. R. Chedekel, “Photophysics and Photochemistry of
Melanin,” In: L. Zeise, M. R. Chedekel and T. B. Fitzpa-
trick, Eds., Melanin: Its Role in Human Photoprotection,
Valdenmar, Overland Park, 1994, pp. 11-22.
[6] J. M. Menter and I. Willis, “Electron Transfer and Photo-
protective Properties on Melanins in Solution,” Pigment
Cell Research, Vol. 10, No. 4, 1997, pp. 214-217.
http://dx.doi.org/10.1111/j.1600-0749.1997.tb00487.x
[7] J. M. C. Gutteridge, I.-Z. Nagy, L. Maidt and R. A. Floyd,
“ADP-Iron as a Fenton Reagent: Radical Reactions De-
tected by Spin-Trapping, Hydrogen Abstraction, and Aro-
matic Hydroxylation,” Archives of Biochemistry and Bio-
physics, Vol. 277 No. 2, 1990, pp. 422-428.
http://dx.doi.org/10.1016/0003-9861(90)90599-T
[8] B. Roznowki, J. M. Burke, M. E. Boulton, T. Sarna and
M. Roznowka, “Human RPE Melanosomes Protect from
Photosensitized and Iron-Mediated Oxidation but Become
Pro-Oxidant in the Presence of Iron upon Photodegra-
dation,” Investigative Ophthalmology and Visual Science,
Vol. 49, No. 7, 2008, pp. 2838-2847.
http://dx.doi.org/10.1167/iovs.08-1700
[9] L. Louw, “The Keloid Phenomenon: Progress toward a
Solution,” Clinical Anatomy, Vol. 20, No. 1, 2007, pp. 3-
14. http://dx.doi.org/10.1002/ca.20374
[10] P. D. Butler, M. T. Longaker and G. P. Yang, “Current
Progress in Keloid Research and Treatment,” Journal of
the American College of Surgeons, Vol. 206, No. 4, 2008,
pp. 731-741.
http://dx.doi.org/10.1016/j.jamcollsurg.2007.12.001
[11] M. C. Naylor and A. E. Brissett, “Current Concepts in
Etiology and Treatment of Keloids,” Facial Plastic Sur-
gery, Vol. 28, No. 5, 2012, pp. 504-512.
http://dx.doi.org/10.1055/s-0032-1325644
[12] A. S. Halim, A. Emami, I. Salahshourifar and T. Ponnuraj,
“Keloid Scarring: Understanding the Genetic Basis, Ad-
vances,” Archives of Plastic Surgery, Vol. 39, No. 3, 2012,
pp. 184-189. http://dx.doi.org/10.5999/aps.2012.39.3.184
[13] S. B. Russell, J. D. Russell, K. M. Trupin, A. E. Gayden,
S. R. Latha Raju and S. M. Williams, “Epigenetically Al-
tered Wound Healing in Keloid Fibroblasts,” Journal of
Investigative Dermatology, Vol. 130, No. 10, 2012, pp.
2489-2496. http://dx.doi.org/10.1038/jid.2010.162
[14] Y. W. Wirohadidjojo, S. Radiono, A. Budiyanto and H.
Soebono, “Cellular Viability, Collagen Deposition and
Transforming Growth Factor β1 Production among Ultra-
violet B-Irradiated Keloids Fibroblasts,” Aesthetic Plastic
Surgery, Vol. 35, No. 6, 2011, pp. 1050-1055.
http://dx.doi.org/10.1007/s00266-011-9732-x
[15] Y.-C. Hsu, M. Hsiao, Y. W. Chien and W.-R. Lee, “Ex-
ogenous Nitric Oxide Stimulated Collagen Type I Expres-
sion in Keloid Fibroblasts by a cGMP-Dependent Man-
ner,” Nitric Oxide, Vol. 16, No. 2, 2006, pp. 258-265.
[16] Y.-C. Hsu, M. Hsaio, L.-F. Wang, Y.-W. Chien and W.-R.
Lee, “Nitric Oxide Produced by iNOS Is Associated with
Collagen Synthesis and Keloid Scar Formation,” Nitric
Oxide, Vol. 14, No. 4, 2006, pp. 327-334.
http://dx.doi.org/10.1016/j.niox.2006.01.006
Open Access OJPC