 Open Journal of Genetics, 2013, 3, 201-215 OJGen http://dx.doi.org/10.4236/ojgen.2013.33023 Published Online September 2013 (http://www.scirp.org/journal/ojgen/) An update on the role of prokineticins in human reproduction-potential therapeutic implications Kulvinder Kochar Kaur1*, Gautam Allahbadia2, Mandeep Singh3 1Dr Kulvinder Kaur Centre for Human Reproduction, Jalandhar, India 2Rotunda—A Centre for Human Reproduction, Mumbai, India 3Swami Satyanand Hospital, Jalandhar, India Email: *kulvinder.dr@gmail.com, drallah@gmail.com, gundeep26@hotmail.com Received 8 July2013; revised 5 August 2013; accepted 13 August 2013 Copyright © 2013 Kulvinder Kochar Kaur et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. ABSTRACT Objective: Prokineticin-1 (PROK1) is a recently de- scribed protein with a wide range of functions in- cluding tissue specific angiogenesis, modulation of inflammatory responses and regulation of haemato- poiesis. PROK1 has been found in the steroidogenic organs like ovary, testis, adrenal and specially pla- centa and they have been found to have a role in de- velopment of the olfactory system and GnRH system. The aim was to update the role of PROK1 and PROK2 in human reproduction since the review was provided by Maldono-Perez (2007) on the potentials of prokineticins in reproduction. Design: A review of international scientific literature by a search of Pub- med and the authors files was done for citation of articles relevant to prokineticins in reproduction, be it its role in ovary, testis, uterus with special emphasis on implantation, normal pregnancy, in labour, patho- physiological states like tubal pregnancy, pcos, vari- ous genital tumours, and cases of isolated hypogo- nadotropic hypogonadism with mutations with PROK2/ PROKR2 and studies detailing functional mechanisms. Results: In the normal cycle, PROK1 has been found to have important roles in implantation, regulating several genes like COX-2, IL-8, IL-11, CTGF related to imp lantation. Initially m urine studies revealed a critical role of PROK2 pathway on olfac- tory bulb morphogenesis and GnRH secretion which was accidentally discovered and since then several studies on mutations in PROK2/PROKR2 showed that they underlie some case of KS in humans. Al- though in mouse heterozygote state is not associated with clinical phenotype, most of huma n mu tat ion s ar e heterozygous. Conclusions: Role of PROK-1 in the process of implantation, with a deeper understanding of the process success rates in IVF and ART can be improved, besides understanding the pathophysiology of tubal pregnancy. Further presence in ovarian folli- cles of PROK1 can be used to plan a strategy for treating pcos. Developme nt of a ntagoni sm of PROK’ S may be a helpful strategy in treating preterm labour. Keywords: Prokineticin 1; Prokineticin Receptor 2; Kallmanns Syndrome; Implantation; GnRH Development 1. INTRODUCTION The prokineticins (PROK) are a family of two multifunc- tional secreted proteins called prokineticin 1 (PROK1) [1] and PROK2, alias Bombina variegate 8 (Bv 8) [2]. The names PROK1 and PROK2 were assigned to these pro- teins by Li et al. to reflect their functions in inducing specific and potent contractions of the gastrointestinal tract (GIT). Le Courtier et al. described a growth factor which induced strong and reproducible mitogenic re- sponse in endocrine gland-derived endothelial cells [3]. A similar effect induced by this protein and by vascular endothelial growth factor (VEGF) lead it to be named endocrine gland VEGF (EG-VEGF) [4]. The two pro- teins are structurally unrelated despite several similarities in the function and control mechanisms. The gene that encodes human PROK1 is located on chromosome 1 (1p.13.3) and is encoded by three exons. The mature human PROK1 peptide consists of 86 amino acids. The most active PROK2 peptide consists of 81 amino acids and is encoded by a four-exon gene on chromosome 3 (3p21.1). The additional exon of PROK2 gene can be actively spliced resulting in longer isoform PROK2 L (102 amino acids) whose function is not well understood [5]. The PROK have been shown to regulate *Corresponding author. OPEN ACCESS  K. K. Kaur et al. / Open Journal of Genetics 3 (2013) 201-215 202 Angiogenesis [6], neuron genesis [7], pain sensation [8] intestinal contraction [1], haematopoiesis [4], immune response [9] and reproduction [10]. PROK bind to two closely related G-protein coupled receptors (GPCR) known as prokineticin receptor1 (PROKR1) and PROKR2; with both receptors being able to bind PROK1 and PROK2 with similar affinities [11]. Mature PROK1 and PROK2 are ligands for the highly homologous (85%) GPCR, (PROKR1 and PROKR 2 formerly known as GPR73a and GPR73b respectively). In contrast to high homology exhibited by PROK recap- tors, the ligands PROK1 and PROK2 share only 44% aminoacid identity. Respecting the conserved N terminal AVITGA sequence Kaser et al. (2003) proposed the term AVIT family to classify the prokineticins and their non- mammalian orthologs [12]. Despite only 45% homology PROK1 and PROK2 share two conserved feautures dur- ing molecular evolution essential for bioactivity, 1) a highly conserved hexapeptide AVITGA sequence and 2) their N terminal and a distinctive structural motif con- sisting of ten cysteine residues with five disulphide crosslinking. The striking differential expression in pro- kineticins results in PROK1 being predominately expres- sed in steroidogenic endocrine cells [3], while PROK2 is mainly expressed in nonsteroidogenic cells of the testis and the central nervous system. 2. PROKINETICINS IN FEMALE REPRODUCTIVE FUNCTION Ovary: Prokineticin 1 (PROK1) is expressed in a dy- namic way in elements of sex-cord stromal lineage [13], whereas prokineticin 2 (PROK2) expression is unde- tectable [14]. During follicle maturation, PROK1 and vasculal endothelial growth factor (VEGF) expression are inversely related. In primordial and primary follicles, there is a high expression of PROK1 in granulosa cells but no VEGF expression. 1) Maturing secondary follicles maintain strong PROK1 expression and weak to moder- ate VEGF expression. 2) In contrast, in the antral folli- cles PROK1 is expressed at low levels in theca cells, whereas VEGF expression is very strong in granulosa cells and moderate in theca cells. 3) In the mature atretic follicle PROK1 expression is strong again in residual theca and VEGF expression is weak. The high expres- sion of PROK1 in atretic follicles might relate to hy- poxia (via HIF-α) secondary to regressive/apoptotic changes occurring in these follicles and serves as a signal for remodeling. 4) In the corpus luteum (CL) the mRNA expression of PROK1 increases as the CL matures, whereas VEGF expression is already maximal at the early luteal phase [14,15]. These differential expression patterns suggest that VEGF and PROK1 have different roles in the vascular and nonvascular structures in the CL. The actions of PROK1 in the ovary are likely to be me- diated by PROKR1 and PROKR2 which are expressed in the ovary [11,16]. However their precise localization has not yet been elucidated. Studies in vitro suggest that PROK1 has a role in the proliferation and survival of endothelial cells of bovine corpus luteum [17]. An indi- rect role for angiogenesis in the CL also has been sug- gested following the observations that PROK1 can stimulate the expression of VEGF [18,19]. Practical Implications: Ferrara et al. 2003 studying 13 PCOS human ovaries in comparison to 13 normal ones found a particular high expression of PROK1 in the Ley- dig like hilus cells found in the highly vascularized ovarian hilus. In PCOS ovaries they found strong ex- pression of PROK1 mRNA in theca interna and stroma, which are spatially related to new blood vessel. In con- trast VEGF mRNA expression was most consistently associated with the granulosa cell layer and sometimes the theca, but rarely the stroma. These findings of ex- pression of both VEGF and PROK1 expression in PCOS ovaries but in different cell types, at different stages of differentiation, suggested a complementary functions for the two factors in angiogenesis and possibly cyst forma- tion [14]. 3. POTENTIAL ROLE OF PROKINETICINS IN PREGNANCY 3.1. Implantation PROK1 and PROK2 show differential expression across the menstrual cycle. One of the earliest signs of implan- tation are hyperaemia and endothelial leakage at the im- plantation site [20,21]. There is increased endometrial expression of PROK1 in the midsecretory phase and both PROK1 and PROKR1 increase in first trimester decidua. PROK1 and PROKR1 immuno localize to stromal endo- thelial and glandular epithelial cells of the endometrium and smooth muscle and endothelial cells in the myo- metrium [22,23]. Expression of PROK1 has been shown to be highest during the secretory phase of the menstrual cycle, and it has been proposed that its role maybe in vascular differentiation and spiral artery formation dur- ing the secretory phase. Also, its presence in myometrial smooth muscle as well as intestinal smooth muscle, sug- gests that it may also play a role in myometrial contrac- tions [22]. PROK1 but not PROK2, PROKR1, or PROKR2 ex- pression peaks during the midluteal window of implanta- tion. Evans et al. (2008) demonstrated elevated expres- sion of PROK1 and PROKR1 in first trimester deciduas in comparison to nonpregnant endometrium. Expression of both proteins in first trimester deciduas was localized to glandular epithelium and various compartments within the stroma and endothelial cells of the microvasculature. In addition PROK1 but not PROKR1 was detected in Copyright © 2013 SciRes. OPEN ACCESS  K. K. Kaur et al. / Open Journal of Genetics 3 (2013) 201-215 203 uterine natural killer cells [23]. Gene array analysis of an endometrial epithelial line stably expressing PROKR1 (PROKR1-Ishikawa) demonstrated, PROK1-PROKR1 signalling regulated genes involved in endometrial re- ceptivity and implantation of early pregnancy. These genes included cyclooxy genase 2 (COX2), laeukemia inhibitory factor (LIF) [23,24], interleukin-8 (IL-8) [25], and interleukin 11 (IL-11). Studies have demonstrated regulation of PROK1 by progesterone (P) and human chorionic gonadotropin (hCG) in the endometrium [22,24,26]. Dual immunohistochemical analysis co-lo- calized expression of luteinizing hormone (LH)/hCGre- ceptor, PROK1, PROKR1 and LIF to the glandular epi- thelial cells of the first trimester decidual tissue. PROK1 enhances adhesion of trophoblast cells to fibronectin and laminin matrices, which are mediated predominantly via LIF induction. Hence maternal-embryonic crosstalk in which embryonic hCG via endometrial PROK1 may play a pivotal role in enhancing receptivity and maintaining early pregnancy [24]. Further Cook et al. (2010) demon- strated the mechanism by which PROK1 modulated IL-11 expression via a PROKR1 and a calcineurin/nu- clear factor of activated T cells (NFAT) signaling path- way, on a calcium, guanine nucleotide binding protein (Gq/11) and extracellular signal related kinase (ERK) dependent manner in human endometrium and first tri- mester deciduas [27]. Overexpression of regulator of calcineurin 1isoform 4 (RCAN 1-4)—a negative regulator of calcineurin signaling leads to a reduction in PROK1 induced IL-11, indicating that RCAN 1 - 4 is acting as a negative regulator of the signaling pathway mediating IL-11. IL-11 is known to be essential for successful de- cidualization and implantation. In human endometrial stromal cells IL-11 has been shown to advance proges- terone induced decidualization implying a role for IL-11 in preparing endometrium for implantation [28]. Relaxin and PGE2 have been shown to increase IL-11 mRNA and protein secretion in decidualized endometrial stromal cells [29]. The same gene array analysis also identified connective tissue growth factor (CTGF) as a target for PROK1 [23]. CTGF is a heparin binding 38 kDa cysteine rich peptide that belongs to the CCN (Cyr 61, CTGF, Nov) family of secretory proteins, with biological activi- ties related to cellular proliferation, differentiation, adhe- sion, chemotaxis, migration, apoptosis and extracellular matrix production. It also has a role in regulating im- plantation and placentation [30,31] with expression being increased from placentae from women with preeclamsia compared with normal pregnancy [32]. CTGF expression was upregulated by PROK1 in early pregnancy decidua via activetion of the Gq, PLC, cSrc, EGFR, MAPK/ERK kinase pathway. Treatment of trophoblast derived HTR-8/Svneo cells with 1 µg/ml CTGF significantly in- creased adhesion to collagen IV, and differentiation of the cells into tube like structures in matrigel suggesting CTGF may contribute to the regulation of trophoblast conversion of maternal spiral arteries [33]. Mcdonald et al. 2011 showed that PROK1 signalling via PROKR1 regulated Dickkopf 1 (DKK1) expression, a negative regulator of canonical Wnt signaling, and its function in the nonpregnant endometrium and first trimester decid- uas [34]. DKK1 mRNA expression is elevated during midsecretory phase of the menstrual cycle and expres- sion increases further in first trimester deciduas. DKK1 protein expression is localized to glandular epithelium and stromal cells during the proliferative, early and sec- retory phases. However expression is confined to the stroma in the late secretory phase and first trimester de- ciduas. PROK1 has been shown to regulate the expres- sion of IL-8 and IL-11 via a Gq-calcium-calcineu- rin-NFAT signaling pathway. PROK1 induced DKK1 expression in endometrial epithelial cells and decidual- izes stromal cells stably expressing PROKR1by same pathway. In this pathway calcium dependent activation of calcineurin causes dephosphorylation of NFAT, al- lowing it to translocate to the nucleus and activate NFAT regulated gene transcription. The calcineurin-NFAT sig- naling pathway is regulated by RCAN1-4, an endoge- nous inhibitor which acts to bind to calcineurin and pre- vent its activation of NFAT (Figure 1). The study by Mcdonald et al. (2011) confirmed that RCAN1-4 is a negative regulator of PROK1 mediated DKK1 expres- sion in epithelial cells proliferation, and in the decidua it regulate decidualization of the stroma. The calcineu- rin-NFAT pathway has previously been shown to be in- volved in regulating endometrial epithelial cell prolif- eration [35] and endometrial expression of IL-11. DKK1 expression is known to be increased upon decidualization of human endometrial stromal cells (HESC) in culture [36,37], and was demonstrated to be elevated in first tri- mester decidual tissue where it localizes primarily to the stromal compartment. Recently PROK levels have been shown to increase in stromal cells decidualized in vitro [38,39], and PROK1 is increased in decidualized tissue [23]. When expression of either DKK 1 or PROK1 is knocked down on primary ESC, there is a decrease in expression of the markers of decidualization i.e. -IGFBP1, prolactin, and IL-11 in response to a decidual- izing stimulus. Both DKK1 and PROK1 lie downstream of the progesterone (P)/cyclic AMP signaling cascade with potential for DKK1 to be regulated by P directly and indirectly via P mediated regulation of PROK1. It has been proposed that via a negative regulation of cel- lular proliferation and decidualization, PROK1 mediated DKK 1 expression contributes to the generation of a re- ceptive endometrium and dysregulation of PROK1 me- diated expression of DKK1 may be a contributing factor to infertility and recurrent pregnancy loss. Su et al. dem- Copyright © 2013 SciRes. OPEN ACCESS  K. K. Kaur et al. / Open Journal of Genetics 3 (2013) 201-215 204 Figure 1. Schematic Representation of prokineticin 1 (PROK1) induction of regulator of calcineurin 1 isoform 4 (RCAN1-4)) and interleukin 11 (IL-11). Activation of PROKR1 by PROK1 results in the induction of IL-11 expression. This occurs via coupling to Gq/11 protein. This results in an intracellular in- crease in calcium which activates calcineurin and subsequently dephosphorylates cytoplasmic NFAT. This allows NFAT to migrate to the nucleus and bind to NFAT binding motifs in the promoter of IL-11 and induce its transcription. PROK 1 also upregulates RCAN1-4 expression which acts as a negative regulator and reduces the level of IL-11 transcription by bind- ing to calcineurin and hence inhibiting NFAT dephosphoryla- tion. Gq = Gq protein alpha subunit, Ca2+ = intracellular ion- ized calcium ERK = extracellular signal regulated kinase, Cam-calmodulin, can = Calcineurin catalytic subunit, CnB = Calcineurin regulatory subunit and NFAT = nuclear factor of activated T cells. Courtesy ref no-16 with permission. onstrated a correlation between recurrent miscarriages and genetic polymorphisms in PROK1 and its receptors [40]. Aberrant elevation of PROK1 expression has also been associated with impaired decidualization and re- current miscarriages [38]. EG-VEGF/PROK1 has been identified as one of the five new biomarkers of human endometrial receptivity in the natural cycle besides laminin-β3, microfil associated protein 5, angiopoietin like 1, and nuclear localized factor 2 [41]. 3.2. Proks and the Feto Placental Unit During the first trimester of pregnancy PROK1 and PROKR1 are predominantly expressed in syncytiotro- phoblasts with the highest expression found during cru- cial hypoxic period of placentation i.e. from 8 - 10 wks gestation in contrast to VEGF which is mostly localized to the cytotrophoblast and extravillous trophoblast cells. PROK1 is also expressed in specialized macrophages called hofbauer cells in the placental villi from 6 wks of gestation. PROKR1 mRNA expression was 80 times more as compared to PROKR2 mRNA in trophoblast [42] Both PROK and PROKR1 mRNA appear to be regulated by hypoxia, as supported by the presence of a hypoxic inducible factor (HIF-α) binding sites in the promoter of both PROK1 and PROKR1 [3]. Supported bystudies in the mouse it has therefore been suggested that PROK1 may have a role in trophoblast differentiation and pla- cental angiogenesis during early pregnancy, negatively regulate trophoblast invasion and that its circulating lev- els were significantly higher in preeclampsia patients [43,44] 3.3. Prok in Third Trimester—Human Placenta-Inflammatory Mediator PROK is thought to have a role in immune regulation affecting differentiation of human bonemarrow cells onto distinct monocyte derived cell population primed for release of proinflammatory cytokines. Furthermore on stimulation with LPS, PROK1 primed monocyte. Macro- phages expressed higher levels of TNF-α and IL-12 with a simultaneous decrease of anti-inflammatory IL-10, demonstrating that PROK, not only stimulates differen- tiation of monocytes, but also alters their functions by enhancing their proinflammatory potential [45]. Also in mouse tumour model and in isolated human immune cells it has been demonstrated that PROK2 and PROKR2 are upregulated in peripheral monocytes and neutrophils in response to G-CSF and GM-CSF [46,47]. Encouraged by their findings e.g. upregulation of IL-8 and COX-2 in a PROKR1 overexpressing human endometrial epithelial cell lines, Denison et al. postulated that PROK1 maybe a novel mediator of inflammatory response in term pla- centa and found PROK1 and PROKR1 expressed in term placenta, immunolocalizing to syncytiotrophoblast, cyto- trophoblast, foetal endothelium and macrophages [48]. PROK1 induced a time dependent increase in expression of IL-8 and COX-2 which was dependent on Gq, phos- phorylation of cSrc, epidermal growth factor receptor (EGFR) and MAPK kinase. PROK1 colocalized with IL-8 and COX-2 in placenta as revealed by double im- munoflorescent immunohistochemistry. COX-2 derived prostaglandins alongside with chemokines such as IL-8 act to activate immune cells, enable vascular permeabil- ity and inflammatory cell infiltration during labour. Be- sides this prostaglandins are involved in cervical ripening and uterine contractions and thereby elevated COX-2 is an important marker of ongoing labour. Based on this and microarray analysis revealing expression of PROK2 increasing with the onset of labour in both the myo- metrium and cervix [49], it was proposed by Gorwiec et al. (2011) that PROK1 and PROKR1 may constitute an initiatory pathway for an inflammatory response in third trimester placenta [50]. PROK’S have also been shown to directly induce contractility of smooth muscles. Ana- lysing the promoter regions of PROKS and PROKR’S highlights their potential regulation by pathways acti- vated by infectious agents. Hence Catalano et al. (2010) further proposed that infection could result in premature Copyright © 2013 SciRes. OPEN ACCESS 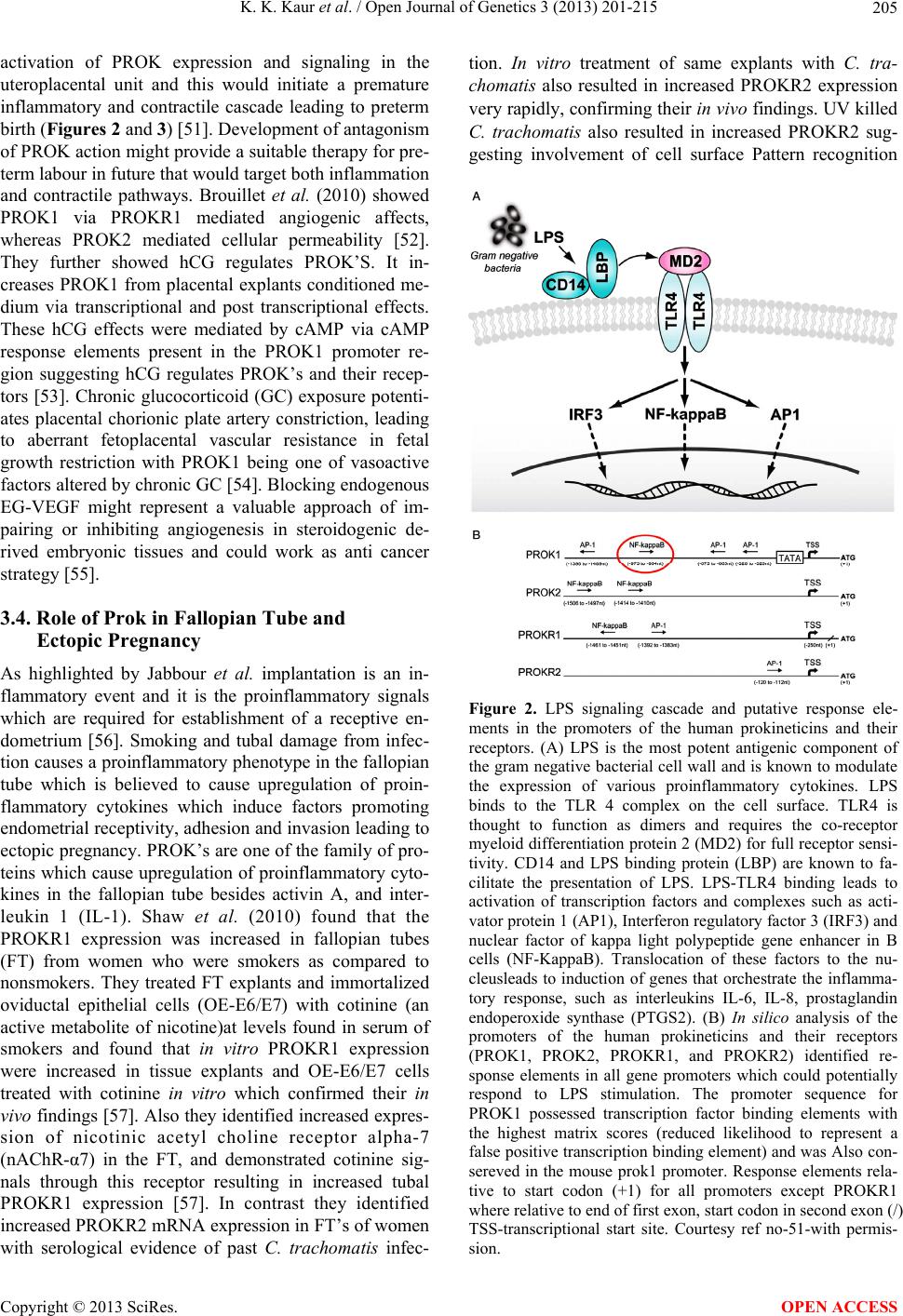 K. K. Kaur et al. / Open Journal of Genetics 3 (2013) 201-215 205 activation of PROK expression and signaling in the uteroplacental unit and this would initiate a premature inflammatory and contractile cascade leading to preterm birth (Figures 2 and 3) [51]. Development of antagonism of PROK action might provide a suitable therapy for pre- term labour in future that would target both inflammation and contractile pathways. Brouillet et al. (2010) showed PROK1 via PROKR1 mediated angiogenic affects, whereas PROK2 mediated cellular permeability [52]. They further showed hCG regulates PROK’S. It in- creases PROK1 from placental explants conditioned me- dium via transcriptional and post transcriptional effects. These hCG effects were mediated by cAMP via cAMP response elements present in the PROK1 promoter re- gion suggesting hCG regulates PROK’s and their recep- tors [53]. Chronic glucocorticoid (GC) exposure potenti- ates placental chorionic plate artery constriction, leading to aberrant fetoplacental vascular resistance in fetal growth restriction with PROK1 being one of vasoactive factors altered by chronic GC [54]. Blocking endogenous EG-VEGF might represent a valuable approach of im- pairing or inhibiting angiogenesis in steroidogenic de- rived embryonic tissues and could work as anti cancer strategy [55]. 3.4. Role of Prok in Fallopian Tube and Ectopic Pregnancy As highlighted by Jabbour et al. implantation is an in- flammatory event and it is the proinflammatory signals which are required for establishment of a receptive en- dometrium [56]. Smoking and tubal damage from infec- tion causes a proinflammatory phenotype in the fallopian tube which is believed to cause upregulation of proin- flammatory cytokines which induce factors promoting endometrial receptivity, adhesion and invasion leading to ectopic pregnancy. PROK’s are one of the family of pro- teins which cause upregulation of proinflammatory cyto- kines in the fallopian tube besides activin A, and inter- leukin 1 (IL-1). Shaw et al. (2010) found that the PROKR1 expression was increased in fallopian tubes (FT) from women who were smokers as compared to nonsmokers. They treated FT explants and immortalized oviductal epithelial cells (OE-E6/E7) with cotinine (an active metabolite of nicotine)at levels found in serum of smokers and found that in vitro PROKR1 expression were increased in tissue explants and OE-E6/E7 cells treated with cotinine in vitro which confirmed their in vivo findings [57]. Also they identified increased expres- sion of nicotinic acetyl choline receptor alpha-7 (nAChR-α7) in the FT, and demonstrated cotinine sig- nals through this receptor resulting in increased tubal PROKR1 expression [57]. In contrast they identified increased PROKR2 mRNA expression in FT’s of women with serological evidence of past C. trachomatis infec- tion. In vitro treatment of same explants with C. tra- chomatis also resulted in increased PROKR2 expression very rapidly, confirming their in vivo findings. UV killed C. trachomatis also resulted in increased PROKR2 sug- gesting involvement of cell surface Pattern recognition Figure 2. LPS signaling cascade and putative response ele- ments in the promoters of the human prokineticins and their receptors. (A) LPS is the most potent antigenic component of the gram negative bacterial cell wall and is known to modulate the expression of various proinflammatory cytokines. LPS binds to the TLR 4 complex on the cell surface. TLR4 is thought to function as dimers and requires the co-receptor myeloid differentiation protein 2 (MD2) for full receptor sensi- tivity. CD14 and LPS binding protein (LBP) are known to fa- cilitate the presentation of LPS. LPS-TLR4 binding leads to activation of transcription factors and complexes such as acti- vator protein 1 (AP1), Interferon regulatory factor 3 (IRF3) and nuclear factor of kappa light polypeptide gene enhancer in B cells (NF-ΚappaB). Translocation of these factors to the nu- cleusleads to induction of genes that orchestrate the inflamma- tory response, such as interleukins IL-6, IL-8, prostaglandin endoperoxide synthase (PTGS2). (B) In silico analysis of the promoters of the human prokineticins and their receptors (PROK1, PROK2, PROKR1, and PROKR2) identified re- sponse elements in all gene promoters which could potentially respond to LPS stimulation. The promoter sequence for PROK1 possessed transcription factor binding elements with the highest matrix scores (reduced likelihood to represent a false positive transcription binding element) and was Also con- sereved in the mouse prok1 promoter. Response elements rela- tive to start codon (+1) for all promoters except PROKR1 where relative to end of first exon, start codon in second exon (/) TSS-transcriptional start site. Courtesy ref no-51-with permis- ion. s Copyright © 2013 SciRes. OPEN ACCESS 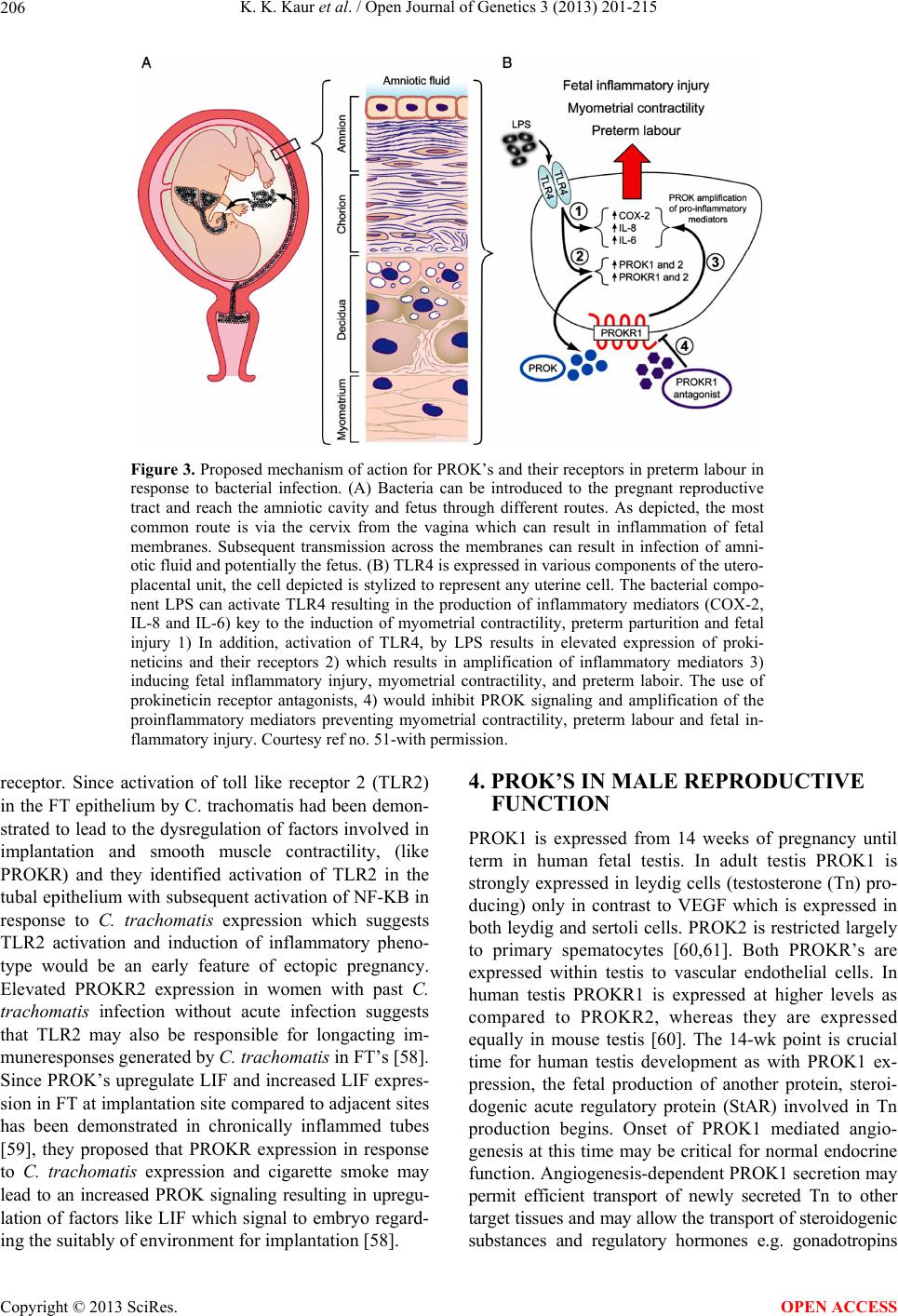 K. K. Kaur et al. / Open Journal of Genetics 3 (2013) 201-215 Copyright © 2013 SciRes. 206 Figure 3. Proposed mechanism of action for PROK’s and their receptors in preterm labour in response to bacterial infection. (A) Bacteria can be introduced to the pregnant reproductive tract and reach the amniotic cavity and fetus through different routes. As depicted, the most common route is via the cervix from the vagina which can result in inflammation of fetal membranes. Subsequent transmission across the membranes can result in infection of amni- otic fluid and potentially the fetus. (B) TLR4 is expressed in various components of the utero- placental unit, the cell depicted is stylized to represent any uterine cell. The bacterial compo- nent LPS can activate TLR4 resulting in the production of inflammatory mediators (COX-2, IL-8 and IL-6) key to the induction of myometrial contractility, preterm parturition and fetal injury 1) In addition, activation of TLR4, by LPS results in elevated expression of proki- neticins and their receptors 2) which results in amplification of inflammatory mediators 3) inducing fetal inflammatory injury, myometrial contractility, and preterm laboir. The use of prokineticin receptor antagonists, 4) would inhibit PROK signaling and amplification of the proinflammatory mediators preventing myometrial contractility, preterm labour and fetal in- flammatory injury. Courtesy ref no. 51-with permission. 4. PROK’S IN MALE REPRODUCTIVE FUNCTION receptor. Since activation of toll like receptor 2 (TLR2) in the FT epithelium by C. trachomatis had been demon- strated to lead to the dysregulation of factors involved in implantation and smooth muscle contractility, (like PROKR) and they identified activation of TLR2 in the tubal epithelium with subsequent activation of NF-ΚB in response to C. trachomatis expression which suggests TLR2 activation and induction of inflammatory pheno- type would be an early feature of ectopic pregnancy. Elevated PROKR2 expression in women with past C. trachomatis infection without acute infection suggests that TLR2 may also be responsible for longacting im- muneresponses generated by C. trachomatis in FT’s [58]. Since PROK’s upregulate LIF and increased LIF expres- sion in FT at implantation site compared to adjacent sites has been demonstrated in chronically inflammed tubes [59], they proposed that PROKR expression in response to C. trachomatis expression and cigarette smoke may lead to an increased PROK signaling resulting in upregu- lation of factors like LIF which signal to embryo regard- ing the suitably of environment for implantation [58]. PROK1 is expressed from 14 weeks of pregnancy until term in human fetal testis. In adult testis PROK1 is strongly expressed in leydig cells (testosterone (Tn) pro- ducing) only in contrast to VEGF which is expressed in both leydig and sertoli cells. PROK2 is restricted largely to primary spematocytes [60,61]. Both PROKR’s are expressed within testis to vascular endothelial cells. In human testis PROKR1 is expressed at higher levels as compared to PROKR2, whereas they are expressed equally in mouse testis [60]. The 14-wk point is crucial time for human testis development as with PROK1 ex- pression, the fetal production of another protein, steroi- dogenic acute regulatory protein (StAR) involved in Tn production begins. Onset of PROK1 mediated angio- genesis at this time may be critical for normal endocrine function. Angiogenesis-dependent PROK1 secretion may permit efficient transport of newly secreted Tn to other target tissues and may allow the transport of steroidogenic substances and regulatory hormones e.g. gonadotropins OPEN ACCESS  K. K. Kaur et al. / Open Journal of Genetics 3 (2013) 201-215 207 from periphery towards testis [62]. Lin et al. 2002 pro- posed that like PROK1, PROK2 functions as a mitogen, chemoattractant, survival factor in adrenal cortical capil- lary epithelial cells (ACE) [63]. Thus PROK’s function as regulators of proliferation and formation of fenestrae in human testis vasculature [60]. In a pilot genome wide association study, atagged single nucleotide polymor- phism in close proximity to PROK2 gene has been shown to be associated with oligozoospermia/azoosper- mia in men [64]. Collectively these observations suggest a role of PROK1 pathway in regulating testicular func- tion and spermatogenesis. Besides that Samsung et al. 2004 found PROK1 expressed in leydig cell tumours but not in seminomas whereas VEGF a powerful angiogenic factor was strongly expressed in seminoma but very weakly in leydig cell tumours [62]. PROK2 expression has been found to be increased by varicocoele induction in rat testis and it may have a role in varicocoele induced infertility [65]. PROK’s expression have been reported in prostate along with their receptors [1,16,66]. But at the protein level PROK1 expression has been reported only in hy- perplastic and cancerous tissue, localized in glandular epithelial cells and progressively increased with the prostate cancer [65]. The role innormal prostate is un- certain as yet. 5. ROLE OFPROK2 PATHWAY IN REPRODUCTION PROK2 is localized in hypothalamic regions critical for GnRH action e.g., preoptic area, arcuate nucleus, and me- dian eminence. It is also expressed in nucleus accumbens, premammilary nucleus, islands of calleja and amyg- dala-regions associated with reproductive and feeding behavior. It is also present in suprachiasmatic nucleus and PROK2 expressing neurons extend their connections to the preoptic area where GnRH neurons reside. There is ample evidence that circadian signals contribute di- rectly to neuroendocrine control of reproduction [67,68]. 5.1. GnRH Deficiency in PROK2 and PROKR Knockout Mice The role of PROK pathway was accidentally discovered in the murine knock out of prok2 and prokr2 while studying the role in gastrointestinal motility and a dis- ruption of neurogenesis of their olfactory bulbs accom- panied by a dramatic reduction in GnRH expressing cells in the median POA along with absence of GnRH neu- ronal projections in the median eminence [69] was found. These findings were a phenocopy of the anatomical ob- servations seen in Kallmanns syndrome (KS) in humans although till then nomurine model to study KS was available as KAL1 gene had never been located in the mouse genome. Approximately 50% of prok2 knockout mice show asymmetrical development of olfactory bulb. The GnRH neurons that do manage to reach hypothala- mus are insufficient in numbers/function to initiate re- productive axis competency. This implies that PROK2 may impact on GnRH neuronal integrity through addi- tional mechanisms besides olfactory bulb neurogenesis [7,69]. Since PROKR2 is not expressed in GnRH neu- rons, elucidation of molecular mechanisms by which PROK2 system regulates GnRH neuronal development and function remains a big challenge. The arrested GnRH neurons formed a fibrocellular mass just beyond cribriform plate immediately prior to their entry into the forebrain [70]. Although all prokr2 mice showed a dra- matic decrease in the olfactory bulb (OB) size [70] only half exhibit an asymmetric olfactory bulb development [69], suggesting a potential redundancy between the two ligands PROK1 and PROK2 in the neurogenesis of OB. prok2 and prokr2 knockout mice with reduced GnRH neurons have a low GnRH secretion resulting in low go- nadotropins and impairment of sexual maturation in both male and female mice. Male prok2 and prokr2 knockouts show small seminiferous tubules which lack lumens, absent haploid spermatocytes and spermatids [69]. Under normal conditions prok2 is heavily expressed in diploid spermatocytes after meiotic division, suggesting a possi- ble role of prok2 in final stages of spermatogenesis. Al- though in female mice incomplete follicular development occurs in mice and humans ovarian function gets re- stored with gonadotropin replacement. 5.2. Genetic Causes of Isolated GnRH Deficiency and PROK2 To date roughly 32% of a large cohort of GnRH deficient patients (n = 397) at the Massachussets general hospital have been linked to atleast one gene mutation knwn to cause human GnRH deficiency. These include a broad spectrum of phenotypes: 1) mild defect of GnRH secre- tion affecting only timing of puberty (delayed puberty), 2) an intermediary defect presenting as spontaneous puberty with subsequent development of permanent hypo- gonadism (acquired HH) or a, 3) severe defect with com- plete/partial absence of puberty (reviewed in [71]). Early developmental genes such as KAL1, FGF8, FGFR1, NELF, CHD7, PROK2 and PROKR2 play a critical role in embryonic neuronal development and subjects with mutations in these genes present primarily with KS. GnRH deficient patients also display a broadspectrum of nonreproductive phenotypes including facial midline defects, skeletal abnormalities and renal agenesis that can provide key clues to the underlying causal gene. Copyright © 2013 SciRes. OPEN ACCESS  K. K. Kaur et al. / Open Journal of Genetics 3 (2013) 201-215 208 5.3. PROK2 and PROKR2 Mutations in Isolated GnRH Deficiency in Humans Following the murine models, Dode et al. (2006) screened 192 unrelated KS patients and found several DNA se- quence changes in both PROK2 and PROKR2 without Any functional studies in the missense cases [72]. In contrast to murine knockout model majority of these rare sequence variants existed only in heterozygous state with four patients with heterozygous mutations in PROK2 and ten patients with heterozygous PROKR2 variants in pa- tients with overt clinical phenotype. Only four patients showed a homozygous/compound heterozygous state, Following that Pittleloud et al. 2007 reported 3 siblings with GnRH deficiency (two brothers and one sister of Portuguese ethnicity and all of them harbouredloss of function homozygous deletion in the ligand, PROK2 which resulted in a biologically inactive 27 amino acid truncated protein [69]. Subsequently a large number of predominantly heterozygous loss of function mutations in both PROK2 and PROKR2 have now been reported in patients with both KS and nIHH by several groups. (Cole et al. (2008) [73], Leroy et al. (2008) [74], Sinisi et al. (2008) [75], Abreu et al. (2008) [76], Canto et al. (2009) [77], Sarfati et al. (2010) [78], Monnier et al. (2009)) [79]. Balasubriam et al. (2011) found a lot of puzzling observa- tions after studying combined analysis of murine and hu- man phenotypes [80]. 1) Although neurodevelopmental role of PROK2 pathwayis key in GnRH development there is conspicu- ous absence of PROKR2 in both developing and mature GnRH neuron. This is further complicated by the recent findings of isolated congenital anosmia (ICA) without gonadotropin deficiency in 25 patients with ICA and olfactory bulb agenesis in whom detailed phenotype analysis and coding sequences of KAL1, FGR1, FGF8, PROK2 and PROKR2 were sequenced. Three PROKR2 mutations previously described in KS, and one new PROK2 mutation were found and investigation of the families showed incomplete penetrance of these muta- tions, which confirms complexity of GnRH neuron de- velopment in humans [81]. This challenges the proposi- tion by Balasubramaniam et al. (2011) that an hitherto unknown early neonatal population expressing PROKR2 may govern the migration of the GnRH neuron by virtue of their chemoattractive interaction with the developing OB which shows a high level of PROK2 expression [80]. 2) As compared to mice which develop a pure neuro- developmental phenotype i.e. a combination of olfactory and reproductive phenotype, humans with PROK2/PRO- KR2 mutations present with both KS as well as normos- mic IHH. This observation suggests that PROK2 path- way plays a key role in both neurodevelopmental and neuroendocrinefacets of GnRH ontogeny. However stu- dying the olfactory phenotypic spectrum in IHH patients Lewkowitz-Shpuntoff et al. 2012 found 31.5% patients were anosmic, 33.6% hyposmic and 34.9% normosmic out of 286 cases of IHH studied [82]. Although tradi- tionally it is believed that KS and nIHH were distinct clinical entities with KS representing a neurodevelop- mental phenotype with a primary defect in GnRH neu- ronal migration, whereas nIHH subjects represent a pure neuroendocrine defect in GnRH secretion/action. Most genes identified in subjects with KS have been shown to play a predominant GnRH migratory role (KAL1, NELF/ PROK2/PROKR2/FGF8/FGFR1) [83], whereas genes identified in nIHH subjects have been shown to primarily affect neuroendocrine regulation of GnRH (GnRH1, GnRHR, TAC3, TACR3, KISS1R) [84]. Thus 1/3 pa- tients of IHH displaying a hyposmic phenotype of which 39.5% harbored mutations in genes affecting neuronal migration like KAL1/PROK2/FGF signalling, suggest a pathophysiological overlap between KS and nIHH, while all PROKR2 variants were monoallelic and associated with anosmia/normosmia. 3) While in mice heterozygous gene deletions are re- portedly normal, in humans mostly clinical syndromes are found with the heterozygous state. The proposed hy- pothesis are a) an autosomal dominant mode of inheri- tance/haploinsufficiency state; b) a dominant negative effect of mutations; or c) oligogenic interactions with other genes/nongenic factors. Although an autosomal dominant state has not been supported [79], oligogenic interactions with mutations in other genes known to cause GnRH deficiency have been documented in some patients with heterozygous mutations in PROK2/PRO- KR2 [73,77,78]. However a dominant negative role for the mutations is still possible, requiring allelic dosing experiments in robust cellular model/organ system to confirm or refute the hypothesis. 4) Humans having identical PROK2/PROKR2 mutations show considerable variations in the expression and penetrance of both their olfactory and reproductive phenotypes. 5) The in vitro functional studies of human PROKR2 mutations show discordant effects on the various intracellular signaling pathways suggesting unique structure functional rela- tionship of the PROKR2 missense variants that have been systematically assessed, some mutations show sig- nificant impairment of receptor function (L173R, P290S, W178 S), while others (R85C, R248Q, V331M), prefer- entially affect either the intracellular calcium influx or the MAPK signaling cascade (R357W) [73,79]. The in- tracellular signaling effect of missense variants show diverse features. The discordant effects of PROKR2 mu- tations may indicate domain specific effects, with more detailed characterization will allow mapping of structure activity relationships and identify critical structural ele- ments of the PROKR2 receptors. Peng et al. (2011) identi- Copyright © 2013 SciRes. OPEN ACCESS  K. K. Kaur et al. / Open Journal of Genetics 3 (2013) 201-215 209 fied PROK2 dose dependently increased the cytoplasmic calcium in cells transfected with WT PROKR2 however R164 Q mutant (mutation in 2nd intracellular (IL2) loop) PROKR2 showed normal cell surface expression and ligand binding capacity, but lost the PROKR2 signalling. R164 Q mutation disrupted the interaction of IL2 loop to the Gαq, Gαi, And Gαi6 proteins [85]. A positive- charged aminoacid at this position is required for proper function, and the signaling efficacy and potency depends on the net amount of positive charges. They also showed that the interactive partner of Arg-164 may localize in the C terminal five residues of Gαq protein. A series of mutation analysis indicated that the basic amino acids at the C terminus of IL2 loop may function cooperatively in GPCR’s. Studying the variants of first intracellular loop (ICL 1) of PROKR2 (R80C, R85C and R85H) identified in patients with HH, Abreu et al. (2012) found that the R85C and R85H PROKR2 mutations, modestly inter- fered with receptor function, in contrast to R80C PROKR2 mutations which lead to marked reduction in receptor activity. Cotransfection of wild type and R80C mutant could exert a dominant negative effect on WT PROKR2 in vitro by interfering with WT receptor ex- pression, hence identifying importance of Arg 80 in ICL1 for PROKR2 expression and demonstration that R80C PROKR2 exerts a dominant effect on WT PROKR2 [86]. 6) Apotential dual effect of PROK2 mutations, as few male patients with mutations of PROK2 pathway display spermatogenic abnormalities, despite gonadotrop in treatment as shown by Sinisi et al. [75] oligozoospermia persisted suggesting a primary gonadal defect as well. As outlined in role in testicular function, these observations suggest a role of PROK2 pathway in regulating primary testicular function and spermatogenesis. While in con- trast to men, in women and female mice with PROK2 deficiency ovarian function gets restored with gonad- otropin replacement. 7) GnRH deficient patients with PROK2/PROKR2 mutations have been shown to be associated with nonre- productive feautures eg bikinesis and hearing loss has been seen in a minority of patients of PROK2/PROKR2 mutation, no cleft lip/renal agenesis has been associated with PROK2 mutations [87]. Also no disturbances in circadian rhythm/sleep disorders identified although in a case controlled Japanese study in patients with mood disorders (151 bipolar patients, 319 with major depres- sive disorders and 319 controls), Kishi et al. (2009) found a tagging SNP in PROKR2, associated with major depressive disorder [88]. Although in prok and prokr knockout mice increased neonatal mortality is reported it has only been reported in 1 family by Pitteloud 2007 with PROK2 mutations, but not in other families or pedigrees with PROK2 mutation [69]. 5.4. PROKR2 in Hypothalamic Amenorrhea Although functional hypothalamic amenorrhea is con- sidered a reversible form of GnRH deficiency, triggered by stressors like excessive exercise, nutritional defects, or psychological distress Caronia et al. (2011) analyz- ing 55 patients of HA found 6 heterozygous mutations in 7 of 55 patients, of which two were in the PROKR2 gene (R85H and L173 R), both of which were loss of function mutations, besides FGR1, GNRHR and KAL1 gene mu- tations suggesting that there is a genetic predisposition to HA in view of differing susceptibility in women to de- velop HA in response to stress [89]. 5.5. Role of PROK2 Pathway in Pituitary Development With the proposed role of PROK2 Pathway in angio- genesis and neuronal migration, Reynaud et al. 2012 reported two heterozygous PROKR2 mutations (p. Leu 173 Arg and pArg 85 His) which had been previously reported in isolated hypogonadotropic hypogonadism (IHH) and a novel PROKR2 variant (pAla 51 Thr) which in contrast to other mutations did not impair receptor signaling. While studying 72 index cases of hypopituita- rism with pituitary stalk interruption syndrome and thus proposed a potential role of PROK pathway in pituitary development and hypothesized that ectopic posterior pituitary may be a consequence of defective axonal pro- jections along the pituitary stalk or defective angiogene- sis of hypophyseal portal circulation [90]. Similarly McCabe et al. 2013 detected five PROKR2 variants in patients of congenital hypopotuitarism (CH), including septooptic dysplasia (SOD). Of 422 patients of complex forms of CH, they detected 5 PROKR2 variants in 11 patients with SOD/CH, novel p.G371R, and previously reported p.A51T, p.R85l, p.L173R, and p.R268C-the latter 3 being known to be functionally deleterious vari- ants [91]. Downregulation of PROK1 in pituitary ade- nomas except LH secreting adenomas suggests LH might be involved in PROK1 secretion [92]. 6. OTHER POTENTIAL ACTIONS In view of various physiological functions, it was pro- posed by Levit et al. (2011) to identify binding sites of known antagonists and additional potential binders to facilitate studying the novel PROKR’s with the view that blocking PROKR’s may serve as therapeutic tool for various diseases, including acute pain, inflammation and cancer. Potential human PROKR ligands with novel scaffolds identified by ligand based pharmacophore models derived from known antagonists and virtual Copyright © 2013 SciRes. OPEN ACCESS  K. K. Kaur et al. / Open Journal of Genetics 3 (2013) 201-215 210 screening performed on Drugbank dataset identified sev- eral HIV protease inhibitors for which endothelial cell dysfunction is a documented sideeffect. Their results suggest that the side effects might be due to inhibition of the PROKR signaling pathway. Docking of known bind- ers to a 3D homology model of PROKR1 is in agreement with the well established canonical TM-bundle binding site of family A GPCR’S. With the exception of a single loop residue that might be perused in the future for ob- taining subtype-specific regulation, their results suggest an identical TM-bundle binding site for PROKR1 and PROKR2, and variable regions may provide subtype specificity [93]. 7. CONCLUSIONS—THERAPEUTIC IMPLICATIONS AND FUTURE DIRECTIONS PROK1 has been found to be involved in the process of embryo implantation and initiation of parturition. It has a role in the pathogenesis of tubal pregnancy secondary to smoking, testicular leydig cell tumors, and other cancers. As PROK1 leads to the expression of various genes stimulated in endometrium and uNK cells including LIF, COX-2, IL-8 and IL-11, this helps in understanding the improved implantation despite knocking off VEGFR2 by cabergoline [94] in the prevention of OHSS attributable to role of PROK1 in early angiogenesis. This knowledge can be used to improve implantation and success rates in IVF and ART. Inhibiting PROK action may delay the onset of preterm labour by suppressing myometrial con- tractility and reducing the premature onset of inflamma- tory pathways known to be critical for induction of la- bour. Simultaneously this PROK signaling suppression may also prove beneficial to preterm neonatal outcome by reducing inflammation associated injury to the fetal brain and lungs. Astrategy for prevention of tubal preg- nancies in smokers can be achieved by understanding aetiopathogenesis of increased LIF expression in tubal mucosa and anticancer strategies in male and female cancers can be evolved. PROK2 signaling is a critical regulator of olfactory bulb morphogenesis and sexual maturation in mammals. Although PROK2 and PROKR2 have emerged as critical regulators of reproduction with PROK2 and PROKR2 mutations having a role in KS, the exact mode of in- heritance however remains controversial. The complex biology of GnRH neuronal development and function has not been fully understood. The recent discovery of muta- tions in the PROK2 pathway in human GnRH deficiency has provided some help, yet many challenges and ques- tions have been opened up. For e.g., in both murine and human PROK2/PROKR2 mutations in homozygous state have provided compelling evidence for the critical role played by the PROK pathway in embryonic migration of GnRH neurons. But the presence of PROK2/PROKR2 mutations in nIHH subjects and the reproductive abnor- malities found in prok knockout mice with partial olfac- tory bulb development is suggestive a potential role for PROK2 beyond GnRH neuronal migration. Further GnRH neurons don’t express PROK receptors which make the matter further perplexing. Also, although mice with heterozygous mutations do not show overt defects, humans with missense mutations present with clinical phenotypes. Although a monogenic recessive mode of inheritance has been clearly demonstrated, it has been done in very few cases. On top of that many of the het- erozygous mutations have also been identified in clini- cally unaffected individuals. Moreover a dominant nega- tive effect of the heterozygous mutations of PROKR2 was not demonstrated by in vitro studies, which argues against a monogenic dominant transmission. Hence po- tential digenic and oligogenic transmission has been suggested. Many of the heterozygous mutations of PROKR2 have also been identified in clinically unaf- fected individuals, which raises the question of actual contribution of PROK’S to the HH phenotype. Potential digenic and oligogenic transmission [95] is suggestive; but still further studies are necessary to confirm the ac- tual pathogenic role of heterozygous PROKR2 mutations with GnRH neurons not expressing PROK receptors suggests an intermediary pathway may mediate PROK2 system and the GnRH neuronal network which needs to be elucidated. The mechanism of PROK signaling is also ill understood with the interacting proteins, chaperones, transcription factors or if any 2nd messenger systems exist need to be unearthed. REFERENCES [1] Li, M., Bullock, C.M., Knauer, D.J., Ehlert, F.J. and Zhou, Q.Y. (2001) Identification of two prokineticin cDNAs: Recombinant proteins potently contract gastrointestinal smith muscle. Molecular Pharmacology, 59, 692-698. [2] Mollay, C., Wechselberger, C., Mignogna, G., Neegri, L., Melchiorri, P., Barra, D. and Kreil, G. (1999) Bv8, a small protein from frog skin and its homologue from snake venom induce hyperalgesia in rats. European Jour- nal of Pharmacology, 374, 189-196. doi:10.1016/S0014-2999(99)00229-0 [3] LeCouter, J., Kowalski, J., Foster, J., Hass, P., Zhang, Z., Dillard-Telm, L., Frantz, G., Rangell, L., Deguzman, L., Keller, G.A., Peale, F., Gurney, A., Hillan, K.J. and Ferrara, N. (2001) Identification of an angiogenic mito- gen selective for endothelial gland endothelium. Nature, 412, 877-884. doi:10.1038/35091000 [4] LeeCouter, J.J., Lin, R., Frantz, G., Zhang, Z., Hillan, K. and Ferrara, N. (2003) Mouse endocrine gland-derived vascular endothelial growth factor: A distinct expression pattern from its human ortholog suggests different roles as a regulator of organ specific angiogenesis. Endocri- Copyright © 2013 SciRes. OPEN ACCESS  K. K. Kaur et al. / Open Journal of Genetics 3 (2013) 201-215 211 nology, 144, 2606-2616. doi:10.1210/en.2002-0146 [5] Chen, J., Kuie, C., Sutton, S., Wilson, S., Yu, J.X., Kam- me, F., Mazur, C., Lovenberg, T. and Liu, C. (2005) Identification and pharmacological characterization of prokineticin 2β as a selective ligand for prokineticin re- ceptor 1. Molecular Pharmacology, 67, 2070-2076. doi:10.1124/mol.105.011619 [6] Urayama, K., Guilini, C., Messaddeq, N., Hu, K., Stteen- man, M., Kuurose, H., Ert, G. and Nebigil, C.G. (2007) The prokinecitin receptor-1 (GPR73) promotes cardio- myocyte survival and angiogenesis. The FASEB Journal, 21, 2980-2993. doi:10.1096/fj.07-8116com [7] Ng, K.L., Li, J.D., Cheng, M.Y., Leslie, F.M., Lee, A.G. and Zhou, Q.Y. (2005) Dependence of olfactory bulb neurogenesis on prokinecitin 2 signalling. Science, 308, 1923-1927. doi:10.1126/science.1112103 [8] Negri, L., Lattanzi, R., Giannini, E., Canestrelli, M., Nicotra, A. and Melchoorri, P. (2009) Chapter 11 Bv8/ prokineticins and their receptors: A new pronociceptive system. International Review of Neurobiology, 85, 145- 157. doi:10.1016/S0074-7742(09)85011-3 [9] Monnier, J. and Samson, M. (2008) Cytokine properties of prokineticins. The FEBS Journal, 275, 4014-4021. doi:10.1111/j.1742-4658.2008.06559.x [10] Maldonando-Perez, D., Evans, J., Denison, F., Millar, R.P. and Jabbour, H.N. (2007) Potential roles of the prokineticins in reproduction. Trends in Endocrinology & Metabolism, 18, 66-72. doi:10.1016/j.tem.2006.12.002 [11] Soga, T., Matsumoto, S., Oda, T., Saito, T., Hiyama, H., Takasaki, J., Kamohara, M., Ohishi, T., Matsushime, H. and Furuichi, K. (2002) Molecular cloning and charac- terization of prokineticin receptors. Biochemica et Bio- physica Acta, 1579, 173-179. [12] Kaser, A., Winklmayr, M., Lepperdinger, G. and Kreil, G. (2003) The AVIT family. Secreted cysteine rich verte- brate proteins with diverse functions. EMBO Reports, 4, 469-473. doi:10.1038/sj.embor.embor830 [13] Zhang, L., Yang, N., Conejo-Garcia, J.R., Katsaros, D., Mohamed-Hadley, A., Fracchioli, S., Schlienger, K., Toll, A., Levine, B., Rubin, S.C. and Coukos, G. (2003) Ex- pression of endocrine gland-vascular endothelial growth factor in ovarian carcinoma. Clinical Cancer Research, 9, 264-272. [14] Ferrara, N., Frantz, G., LeCouter, J., Dillard-Telm, L., Pham, T., Dreaksharapu, A., Giordano, T. and Peale, F. (2003) Differential expression of the angiogenic factor genes vascular endothelial growth factor (VEGF) and en- docrine gland-derived VEGF in normal and polycystic human ovaries. The American Journal of Pathology, 162, 1881-1893. doi:10.1016/S0002-9440(10)64322-2 [15] Fraser, H.M., Bell, J., Wilson, H., Taylor, P.D., Morgan, K., Anderson, R.A. and Duncan, W.C. (2005) Localiza- tion and quantification of cyclic changes in the expres- sion of endocrine gland vascular endothelial growth fac- tor in the human corpus luteum. The Journal of Clinical Endocrinology & Metabolism, 90, 427-434. [16] Lin, D.C., Bullock, C.M., Ehlert, F.J., Chen, J.L., Tian, H. and Zhou, Q.Y. (2002) Identification and molecular characterization of two closely related G protein-coupled receptors activated by prokineticins/endocrine gland vas- cular endothelial growth factor. The Journal of Biological Chemistry, 277, 19276-19280. doi:10.1074/jbc.M202139200 [17] Kisloiuk, T., Podlovni, H., Spanel-Borowski, K., Ovadia, O., Zhou, Q.Y. and Meidan, R. (2005) Prokineticins (en- docrine gland-derived vascular endothelial growth factor and Bv8) in the bovine ovary: Expression and role as mi- togens and survival factors for corpus luteum-derived endothelial cells. Endocrinology, 146, 3950-3958. doi:10.1210/en.2005-0297 [18] Kisliouk, T., Podlovni, H. and Meidan, R. (2005) Unique expression and regulatory mechanisms of EG-VEGF/ prokineticin1 and its receptors in the corpus luteum. An- nals of Anatomy, 187, 529-537. doi:10.1016/j.aanat.2005.07.005 [19] Kisliouk, T., Friedman, A., Klipper, E., Zhou, Q.Y., Schams, D., Alfraidy, N. and Meidan, R. (2007) Expres- sion pattern of prokineticin 1 and its receptors in bovine ovaries during the estrous cycle: Involvememt in corpus luteum regression and follicular atresia. Biology of Re- production, 76, 749-758. doi:10.1095/biolreprod.106.054734 [20] Plaks, V., Kachenko, V., Dekel, N. and Neeman, M. (2006) MRI analysis of angiogenesis during mouse em- bryo implantation. Magnetic Resonance in Medicine, 55, 1013-1022. [21] Plaks, V., Bimberg, T., Beerkutzki, T., Sela, S., Ben Ya- shar, A., Kalchenko, V., Mor, G., Keshet, D., Dekel, N., Neeman, M. and Jung, S. (2008) Uterine DC’s are crucial for decidua formation during embryo implantation. Jour- nal of Clinical Investigation, 118, 3954-3965. [22] Battersby, S., Critchley, H.O., Morgan, K., Millar, R.P. and Jabbour, H.N. (2004) Expression and regulation of the prokineticins (endocrine gland derived vascular en- dothelial growth factor and Bv8) and their recptors in the human endometrium across the menstrual cycle. The Journal of Clinical Endocrinology & Metabolism, 89, 2463-2469. doi:10.1210/jc.2003-032012 [23] Evans, J., Catalano, R.D., Morgan, K., Critchley, H.O., Millar, R.P. and Jabbour, H.N. (2008) Prokineticin 1 sig- nalling and gene regulation in early human pregnancy. Endocrinology, 149, 2877-2887. doi:10.1210/en.2007-1633 [24] Evans, J., Catalano, R.D., Brown, P., Sherwin, R., Crit- chley, H.O., Fazleabas, A.T. and Jabbour, H.N. (2009) Prokineticin 1 mediates fetal-maternal dialogue regarding endometrial leukemia inhibitory factor. The FASEB Jour- nal, 23, 2165-2175. doi:10.1096/fj.08-124495 [25] Maldonado-Perez, D., Evans, J., Denison, F., Millar, R.P., Thompson, E.A. and Jabbour, H.N. (2009) Prokineticin 1 modulates IL-8 expression via the calcineurin/NFAT signaling pathway. Biochimica et Biophysica Acta, 1793, 1315-1324. doi:10.1016/j.bbamcr.2009.03.008 [26] Ngan, E.S., Lee, K.Y., Yeung, W.S., Ngan, H.Y., Ng, E.H. and Ho, P.C. (2006) Endocrine gland-derived vas- cular endothelial growth factor is expressed in human periimplantation endometrium, bit not in endometrial carcinoma. Endocrinology, 147, 88-95. Copyright © 2013 SciRes. OPEN ACCESS  K. K. Kaur et al. / Open Journal of Genetics 3 (2013) 201-215 212 doi:10.1210/en.2005-0543 [27] Cook, I.H., Evans, J., Maldonaldo-Perez, D., Critchley, H.O., Sales, K.J. and Jabbour, H.N. (2010) Prokineticin 1 (PROK1) modulates interleukin (IL)-11 expression via prokineticin receptor 1 (PROKR1) and the calcineurin/ NFAT signaling pathway. Basic Science of Reproductive Medicine, 16, 158-169. doi:10.1093/molehr/gap084 [28] Dimitriadis, S., Robb, L. and Salamonsen, L.A. (2002) Interleukin 11 advances progesterone induced deciduali- zation of human endometrial stromal cells. Basic Science of Reproductive Medicine, 8, 636-643. doi:10.1093/molehr/8.7.636 [29] Dimitriadis, E., Stoikos, C., Baca, M., Fairlie, W.D., McCoubrie, J.E. and Salamonsen, L.A. (2005) Relaxin and prostaglandin E2 regulate interleukin 11 during hu- man endometrial stromal cell decidualization. The Jour- nal of Clinical Endocrinology & Metabolism, 90, 3458- 3465. doi:10.1210/jc.2004-1014 [30] Roh, C.R., Budhraja, V., Kim, H.S., Nelson, D.M. and Sadosky, Y. (2005) Microarrey based identification of differentially expressed genes in hypoxic term human tro- phoblasts and in placental villi of pregnancies with growth restricted fetuses. Placenta, 26, 319-328. doi:10.1016/j.placenta.2004.06.013 [31] Rimon, E., Chen, B., Shanks, A.L., Nelson, D.M. and Sadovsky, Y. (2008) Hypoxia in human trophoblasts stimulates the expression and secretion of connective tis- sue growth factor. Endocrinology, 149, 2852-2958. doi:10.1210/en.2007-1099 [32] Oh, S.Y., Song, S.E., Seo, E.S., Kim, K.H., Choi, S.J., Suh, Y.L., Sadovsky, Y. and Roh, C.R. (2009) The ex- pression of connective tissue growth factor in pregnancies complicated by severe preeclampsia or fetal growth re- striction. Placanta, 30, 981-987. doi:10.1016/j.placenta.2009.08.006 [33] Waddell, J.M., Evans, J., Jabbour, H.N. and Denison, F.C. (2011) CTGF expression is upregulated by PROK1 in early pregnancy and influence ahtr-8/Syneo Cell adhesion and network formation. Human Reproduction, 26, 67-75. [34] Macdonald, L.J., Sales, K.J., Grant, V., Brown, P., Jab- boir, H.N. and Catalano, R.D. (2011) Prokineticin 1 in- duces Diccopf1 expression and regulates cell prolifera- tion and decidualization in the human endometrium. Ba- sic Science of Reproductive Medicine, 17, 626-636. doi:10.1093/molehr/gar031 [35] Wallace, A.E., Catalano, R.D., Anderson, R.A. and Jab- bour, H.N. (2011) Chemokine (C-C) motif ligand 20 os regulated by PGF (2alpha)-F-prostanoid receptor signal- ing in endometrial adenocarcinoma and promote cell pro- liferation. Molecular and Cellular Endocrinology, 331, 129-135. doi:10.1016/j.mce.2010.08.018 [36] Tulac, S., Overgaard, M.T., Hamilton, A.E., Jumbe, N.L., Suchanek, E. and Giudice, L.C. (2006) Diccopf-1 an in- hibitor of Wnt signaling, is regulated by progesterone in human endometrial stromal cells. The Journal of Clinical Endocrinology & Metabolism, 91, 1453-1461. doi:10.1210/jc.2005-0769 [37] Kane, N., Jones, M., Brosens, J.J., Saunders, P.T., Kelly, R.W. and Critchley, H.O. (2008) Transforming growth factor-bera1 attenuates expression of both the progester- one receptor and dickkopf in differentiated human endo- metrial stromal cells. Molecular Endocrinology, 22, 716- 728. doi:10.1210/me.2007-0316 [38] Salker, M., Teklenburg, G., Molokhia, M., Lavery, S., Trew, G., Aojanepang, T., Mardon, H.J., Lokugamage, A.U., Rai, R., et al. (2010) Natural selection of human embryos: Impaired decidualization of endometrium dis- ables embryo-maternal interaction and causes recurrent pregnancy loss. PLoS One, 5, e10287. doi:10.1371/journal.pone.0010287 [39] Tiberi, F., Tropea, A., Romani, F., Apa, R., Marana, R. and Lanzone, A. (2010) Prokineticin 1, homeobox A10 and progesterone receptor messenger ribonucleic acid expression in primary cultures of endometrial stromal cells isolated from endometrium of healthy women and from eutopic endometrium of women with endometriosis. Fertility and Sterility, 94, 2558-2563. doi:10.1016/j.fertnstert.2010.03.006 [40] Su, M.T., Lin, S.H., Lee, I.W., Chen, Y.C., Hsu, C.C., Pan, H.A. and Kuo, P.L. (2010) Polymorphisms of endo- crine gland-derived vascular endothelial growth factor geneand its receptor genes are associated with recurrent pregnancy loss. Human Reproduction, 25, 2923-2930. doi:10.1093/humrep/deq256 [41] Haouzi, D., Mahmoud, K., Fourar, M., Bendhaou, K., Dechaud, H., Vos, J.D., Rerne, T., Dewailly, D. and Hamamah, S. (2009) Identification of new biomarkers of human endometrial receptivity in the natural cycle. Hu- man Reproduction, 24,198-205. doi:10.1093/humrep/den360 [42] Hoffmann, P., Feige, J.J. and Alfaidy, N. (2006) Expres- sion and oxygen regulation of endocrine gland derived vascular endothelial growth factor/prokineticin-1 and its recptors in human placenta during early pregnancy. En- docrinology, 147, 1675-1684. doi:10.1210/en.2005-0912 [43] Hoffmann, P., Feige, J.J. and Alfaidy, N. (2007) Placental expression of EG-VEGF and its receptors PKR1 (proki- neticin receptor-1) and PKR2 throughout mouse gesta- tion. Placenta, 28, 1049-1058. doi:10.1016/j.placenta.2007.03.008 [44] Hoffmann, P., Saoudi, Y., Benharouga, M., Graham, C.H., Schaal, J.P., Mazouni, C., Feige, J.J. and Alfaidy, N. (2009) Role of EG-VEGF in human placentation: Physiological and pathological implications. Journal of Cellular and Molecular Medicine, 13, 2224-2235. doi:10.1111/j.1582-4934.2008.00554.x [45] Dorsch, M., Qiu, Y., Soler, D., Frank, N., Duong, T., Gooearl, A., O’Neil, S., Lora, J. and Fraser, C.C. (2005) PK1/EG-VEGF-induces monocyte differentiation and ac- tivation. Journal of Leukocyte Biology, 78, 426-434. doi:10.1189/jlb.0205061 [46] Shojaei, F., Singh, M., Thompson, J.D. and Ferrara, N. (2008) Role of bv8 in neutrophil dependent angiogenesis in a transgenic model of cancer progression. Proceedings of the National Academy of Sciences of the United States of America, 105, 2675-2684. doi:10.1073/pnas.0712185105 Copyright © 2013 SciRes. OPEN ACCESS  K. K. Kaur et al. / Open Journal of Genetics 3 (2013) 201-215 213 [47] Zhong, C., Qu, X., Tan, M., Meng, Y.G. and Ferrara, N. (2009) Characterization and regulation of bv8 in human blood cells. Clinical Cancer Research, 15, 2675-2684. doi:10.1158/1078-0432.CCR-08-1954 [48] Denison, F.C., Battersby, S., King, A.E., Szuber, M. and Jabbour, H.N. (2008) Prokineticin-1 a novel mediator of of the inflammatory response in third trimester human placenta. Endocrinology, 149, 3470-3477. doi:10.1210/en.2007-1695 [49] Bollapragada, S., Youssef, R., Jordan, F., Greer, I., Nor- man, J. and Nelson, S. (2009) Term labour is associated with core inflammatory response in human fetal mem- branes, myometrium and cervix. American Journal of Obstetrics and Gynecology, 200, 104. e1-e11. [50] Gorowiec, M.R., Catalano, R.D., Norman, J.E., Denison, F.C. and Jabbour, H.N. (2011) Prokineticin 1 induces in- flammatory response in human myometrium: A Potential role in initiating term and preterm parturition. The American Journal of Pathology, 179, 2709-2719. doi:10.1016/j.ajpath.2011.08.029 [51] Catalano, R.D., Lannagan, T.R.M., Gorowiec, M., Deni- son, F.C., Norman, J.E. and Jabbour, H.N. (2010) Proki- neticins: Novel mediators of inflammatory and contractile pathways at parturition? Molecular Human Reproduction, 16, 311-319. doi:10.1093/molehr/gaq014 [52] Brouillet, S., Hoffmann, P., Benharouga, M., Salomon, A., Schaal, J.P., Feige, J.J. and Alfaidy, N. (2010) Mo- lecular characterization of EG-VEGF-mediated angio- genesis: Differential effects on microvascular and macrovascular endothelial cells. Molecular Biology of Cell, 21, 2832-2843. doi:10.1091/mbc.E10-01-0059 [53] Brouillet, S., Hoffmann, P., Chauvet, S., Salomon, A., Chamboredon, S., Sergent, F., Benharouga, M., Feige, J.J. and Alfaidy, M. (2012) Revisiting the role of hCG: New regulation of the angiogenic factor EG-VEGF and its re- ceptors. Cellular and Molecular Life Sciences, 69 , 1537- 1550. doi:10.1007/s00018-011-0889-x [54] Nugent, J.L., Wareing, M., Palin, V., Sibley, C.P., Baker, P.N., Ray, D.W., Farrow, S.N. and Jones, R.L. (2013) Chronic glucocorticoid exposure potentiates placental chorionic plate artey constriction: Implications for aber- rant fetoplacental vascular resistance in fetal growth re- striction. Endocrinology, 154, 876-887. doi:10.1210/en.2012-1927 [55] Feflea, S., Maria, A., Cimpean, A.M., Ceausu, R.A., Gaje, P. and Raica, M. (2012) Effects of antibodies to EG- VEGF on angiogenesis in the chick embryo chorioallan- toic membrane. In Vivo, 26, 793-797. [56] Jabbour, H.N., Kelly, R.W., Fraser, H.M. and Critchley, H.O. (2006) Endocrine regulation of menstruation. Endo- crine Reviews, 27, 17-46. [57] Shaw, J.L., Oliver, E., Lee, K.F., Entrican, G., Jabbour, H.N., Critchley, H.O. and Horne, A.W. (2010) Cotinine exposure increases fallopian tube PROKR1 expression via nicotinic AchR(alpha)-7: A potential mechanism ex- plaining the link between smoking and tubal ectopic pregnancy. The American Journal of Pathology, 177, 2509-2515. doi:10.2353/ajpath.2010.100243 [58] Shaw, J.L., Wills, G.S., Lee, K.E., Horner, P.J., McClure, M.O., Abrahams, V.M., Wheelhouse, N., Jabbour, H.N., Critchley, H.O., Entrican, G. and Horne, A.W. (2011) Chlamydia trachomatis infection increases fallopian tube PROKR2 via TLR2 and NFkappaB activation resulting in a microenvironment predisposed to ectopic pregnancy. The American Journal of Pathology, 178, 253-260. doi:10.1016/j.ajpath.2010.11.019 [59] Ji, Y.F., Chen, L.Y., Xu, K.H., Yao, J.F. and Shi, Y.F. (2009) Locally elevated leukemia inhibitory factor in the inflamed fallopian tube resembles that found in tubal pregnancy. Fertility and Sterility, 91, 2308-2314. doi:10.1016/j.fertnstert.2008.01.110 [60] LeCouter, J., Lin, R., Tejada, M., Frantz, G., Peale, F., Hillan, K.J. and Ferrara, N. (2003) The endocrine-gland- derived VEGF homologue bv8 promotes angiogenesis in the testis: Localization of bv8 receptors to endothelial cells. Proceedings of the National Academy of Sciences of the United States of America, 100, 2685-2690. doi:10.1073/pnas.0337667100 [61] Wechselnberger, C., Puglisi, R., Engel, E., Lepperdinger, G., Boitani, C. and Kreil, G. (1999) The mammalian homologues of frog bv8 are mainly expressed in sper- matocytes. FEBS Letters, 462, 177-181. doi:10.1016/S0014-5793(99)01473-8 [62] Sampson, M., Peale Jr., F.V., Frantz, G., Rioux-Leclercq, N., Rajpert De Meyts, E. and Ferrara, N. (2004) Human endocrine-gland derived vascular endothelial growth fac- tor: Expression early in development and in leydig cell tumors suggests roles in normal and pathological testis angiogenesis. The Journal of Clinical Endocrinology & Metabolism, 89, 4078-4088. doi:10.1210/jc.2003-032024 [63] Lin, R., LeCouter, J., Kowalski, J. and Ferrara, N. (2002) Characterization of endocrine gland derived vascular en- dothelial growth factor signaling in adrenal cortex capil- lary endothelial cells. The Journal of Biological Chemis- try, 277, 8724-8729. doi:10.1074/jbc.M110594200 [64] Aston, K.L. and Carrell, D.I. (2009) Genome-wide study of single-nucleotide polymorphisms associated with azo- ospermia and severe oligozoospermia. Journal of An- drology, 30, 711-725. doi:10.2164/jandrol.109.007971 [65] Tu, L.H., Yu, L.L., Xiong, C.L. and Zhang, H.P. (2012) Potential role of prokineticin 2 in experimental varico- coele-induced rat testes. Urology, 80, 952.e15-e19. [66] Pasquali, D., Rossi, V., Staibano, S., DeRosa, G., Chieffi, P., Prezioso, D., Mirone, V., Mascolo, M., Tramantano, D., Bellastella, A. and Sinisi, A.A. (2006) The endocrine gland-derived vascular endothelial growth factor (EG- VEGF/prokineticin 1 and 2 and receptor expression in human prostate: Upregulation of EG-VEGF/prokineticin 1 with malignancy. Endocrinology, 147, 4245-4251. doi:10.1210/en.2006-0614 [67] De la Iglesia, H.O. and Schwaerz, W.J. (2006) Minire- view: Timely ovulation: Circadian regulation if the fe- malehypothalamo-pituitary-gonadalaxis. Endocrinology, 2147, 1148-1153. [68] Ward, D.R., Dear, F.M., Ward, J.A., Anderson, S.I., Spergel, D.J., Smith, P.A. and Ebling, F.J. (2009) Inner- vation of gonadotropin-releasing hormones neurons by peptidergic neurons conveying circadian or energy bal- Copyright © 2013 SciRes. OPEN ACCESS  K. K. Kaur et al. / Open Journal of Genetics 3 (2013) 201-215 214 ance information in the mouse. PLoS One, 4, e5.322. [69] Pitteloud, N., Zhang, C., Pognatelli, D., Li, J.D., Raivio, T., Cole, L.W., Plumer, L.W., Jacobson-Dickman, E.E., Mellon, P.L., Zhou, Q.Y. and Crowley Jr., W.F. (2007) Loss of function mutation in the prokineticin 2 gene causes Kallmann syndrome and normosmic idiopathic hypogonadotropic hypogonadism. Proceedings of the Na- tional Academy of Sciences of the United States of Amer- ica, 104, 17447-17452. doi:10.1073/pnas.0707173104 [70] Matsumoto, S., Yamazaki, C., Masumoto, K.H., Nagano, M., Naito, M., Soga, T., Hiyama, H., Matsumoto, M., Tkasaki, J., Kamohara, M., Matsuo, A., Ishii, H., Kabori, M., et al. (2006) Abnormal development of the olfactory bulb and reproductive system in mice lacking proki- neticin receptor PKR2. Proceedings of the National Academy of Sciences of the United States of America, 103, 4140-4145. doi:10.1073/pnas.0508881103 [71] Martin, C., Balasubramanium, R., Dwyer, A.A., Au, M.G., Sidis, Y., Kaiser, U.B., Seminara, S.B., Pitteloud, N., Zhou, Q.Y. and Crowley Jr., W.F. (2011) The role of the prokineticin 2 pathway in human reproduction: Evi- dence from the study of human and murine mutations. Endocrine Reviews, 32, 225-246. doi:10.1210/er.2010-0007 [72] Dode, C., Teixeira, L., Levvilliers, J., Fouveaut, C., Bouchard, P., Kottler, M.L., Lespinasse, J., Lienhart- Roussie, A., Mathew, M., Mierman, A., et al. (2006) Kalmann syndrome: Mutations in the genes encoding prokineticin 2 and prokineticin receptor 2. PLoS Gene, 2, e175. [73] Cole, L.W., Sidis, Y., Zhang, C., Quinton, R., Plummer, L., Pignarelli, D., Hughes, V.A., Dwyer, A.A., Raivio, T., Hayes, F.J., Seminara, S.B., Huot, C., Alos, N., Speiser, P., Takeshita, A., et al. (2008) Mutations in prokineticin2 and prokineticin receptor genes in human gonadotro- phin-releasing hormone deficiency: Molecular genetics and clinical spectrum. The Journal of Clinical Endocri- nology & Metabolism, 93, 3551-3559. doi:10.1210/jc.2007-2654 [74] Leroy, C., Fouveaut, C., Leclercq, S., Jacquemont, S., Boullay, H.D., Lespinasse, J., Delpech, M., Dupont, J.M., Hardelin, J.P. and Dode, C. (2008) Biallelic mutations in the prokineticin gene in two sporadic case of Kallmann syndrome. European Journal of Human Genetics, 16, 865-868. doi:10.1038/ejhg.2008.15 [75] Sinisi, A.A., Asci, R., Bellastella, G., Maione, L., Espo- sito, D., Elefante, A., DeBellis, A., Bellastella, A. and Io- lascon, A. (2008) Homozygous mutation in the proki- neticin receptor 2 gene (Val 274Asp) presenting as re- versible Kallmann syndrome and persistent oligozoo- spermia: Case report. Human Reproduction, 23, 2380- 2384. doi:10.1093/humrep/den247 [76] Abreu, A.P., Trarbach, E.B., de Castro, M., Frade-Costa, E.M., Versiani, B., Matias-Baptista, M.T., Garmes, H.M., Mendonca, B.B. and Latronico, A.C. (2008) Loss of function mutations in the genes encoding prokineticin 2 or prokineticin receptor 2 cause autosomal recessive Kallmann syndrome. The Journal of Clinical Endocri- nology & Metabolism, 93, 4113-4118. doi:10.1210/jc.2008-0958 [77] Canto, P.M., Munguia, P., Soderlund, D., Castro, J.J. and Mendez, J.P. (2009) Genetic analysis in patients with Kallmann syndrome: Coexisrence of mutations proki- neticin receptor 2 and KAL1. Journal of Andrology, 30, 41-45. doi:10.2164/jandrol.108.005314 [78] Sarfati, J., Guiochon-Mantel, A., Rondard, P., Arnulf, L., Garcio-Pinero, A., Wolczynski, S., Brailly-Tabard, S., Bidet, M., Ramos-Arroyo, M., Mathieu, M., Lienhardt- Roussie, A., Morgan, G., et al. (2010) A comparative phenotypic study of Kallmann syndrome patients carry- ing monoallelic and biallelic mutations in the proki- neticin 2 or prokineticin receptor 2 genes. The Journal of Clinical Endocrinology & Metabolism, 85, 659-669. doi:10.1210/jc.2009-0843 [79] Monnier, C., Dode, C., Fabre, L., Teixeira, L., Labesse, G., Pin, J.P., Hardelin, J.P. and Rondard, P. (2009) PROKR2 missense mutations associated with Kallmann syndrome impair receptor signaling activity. Human Mo- lecular Genetics, 18, 75-81. doi:10.1093/hmg/ddn318 [80] Balasubramaniam, R., Plummer, L., Sidis, Y., Pitteloud, N., Cecila, N., Zhou, Q.Y. and Crowley Jr., W.F. (2011) The puzzles of the prokineticin 2 pathway in human re- production. Molecular and Cellular Endocrinology, 346, 44-50. doi:10.1016/j.mce.2011.05.040 [81] Plana, A.M., Villanueva, C., Laccourreye, O., Bonfils, P. and de Roux, N. (2013) PROKR2 and PROK2 mutations cause isolated congenital anosmia without gonadotropic deficiency. European Journal of Endocrinology, 168, 31- 37. doi:10.1530/EJE-12-0578 [82] Lewkowitz-Shupntoff, H.M., Hughes, V.A., Plummer, L., Au, M.G., Doty, R.L., Seminara, S.B., Chan, Y.M., Pot- teloud, N., Crowley Jr., W.F. and Balasubramaniam, R. (2012) Olfactory phenotypic spectrums in idiopathic hy- pogonadotropic hypogonadism: Pathophysiological and genetic implications. The Journal of Clinical Endocri- nology & Metabolism, 97, E136-E144. doi:10.1210/jc.2011-2041 [83] Hardelin, J.P. and Dode, C. (2008) The complex genetics of Kallmann syndrome: KAL1, FGFR1, FGF8, PROKR2, PROK2, et al. Sexual Development, 2, 181-193. doi:10.1159/000152034 [84] Chan, Y.M., De Guillebon, A., Lang-Muritano, M., Plummer, L., Cerrato, F., Tsiaras, S., Gaspert, A., Lavoie, H.B., Crowley Jr., W.F., Amory, K., Pitelloud, N. and Seminara, S.B. (2009) GNRH1 mutations in patients with idiopathic hypogonadotropic hypogonadism. Proceedings of the National Academy of Sciences of the United States of America, 106, 11703-11708. doi:10.1073/pnas.0903449106 [85] Peng, Z., Tang, Y., Luo, H., Jiang, F., Sun, L. and Li, J.D. (2011) Disease causing mutation in PKR2 receptor re- veals a critical role of positive charges in the second in- tracellular loop for G-protein coupling and receptor traf- ficking. The Journal of Biological Chemistry, 286, 16615-16622. doi:10.1074/jbc.M111.223784 [86] Abreu, A.P., Noel, S.D., Xu, S., Carroll, R.S., Ltronico, A.C. and Kaiser, U.B. (2012) Evidence of the first intra- cellular loop of prokineticin receptor 2 in receptor func- tion. Molecular Endocrinology, 26, 1417-1427. doi:10.1210/me.2012-1102 Copyright © 2013 SciRes. OPEN ACCESS  K. K. Kaur et al. / Open Journal of Genetics 3 (2013) 201-215 Copyright © 2013 SciRes. 215 OPEN ACCESS [87] Abreu, A.P., Kaiser, U.B. and Latronico, A.C. (2010) The role of prokineticins in the pathogenesis of hypogonado tropic hypogonadism. Neuroendocrinology, 91, 81-90. doi:10.1159/000308880 [88] Kishi, T., Kitajima, T., Tsunoka, T., Okumura, T., Ikeda, M., Okochi, T., Kinoshita, Y., Kawashima, K., Yama- nouchi, Y., Ozaki, N. and Iwara, N. (2009) Possible asso- ciation of prokineticin receptor 2 gene (PROKR2) with mood disorders in the Japanese population. Neuromo- lecular Medicine, 11, 114-122. [89] Caronia, L.M., Martin, C., Welt, K.C., Sykiotis, G.P., Quinton, R., Thambundit, A., Avbelj, M., Dhruvakumar, S., Plummer-Hughes, V.A., Seminara, S.B., et al. (2011) A genetic basis for functional hypothalamic amenorrhea. The New England Journal of Medicine, 364, 215-225. doi:10.1056/NEJMoa0911064 [90] Reynaud, R., Jayakody, S.A., Monnier, C., Saveanu, A., Bouligand, J., Guedj, A.M., Simonin, G., Lecomte, M., Barlier, A., Rondard, P., Martinez-Barbera, J.P., Guio- chon-Mantel, A. and Brue, T. (2012) PROKR2 variants in multiple hypopituitarism with pituitary stalk interrupt- tion. The Journal of Clinical Endocirnology & Metabo- lism, 97, E1068-E1073. doi:10.1210/jc.2011-3056 [91] McCabe, M.J., Gaston-Massuet, C., Gregory, L.C., Alat- zoglu, K.S., Tziaferi, V., Sbai, O., Rondard, P., Masu- moto, K.H., Nagano, M., Shigeyoshi, Y., Pfiefer, M., Hulse, T., Buchanan, C.R., Potteloud, N., Martinez-Bar- bera, J.P. and Dattani, M.T. (2013) Variations in PROKR2 but not PROK2, are associated with hypopitui- tarism and septo-optic dysplasia. The Journal of Clinical Endocirnology & Metabolism, 98, E547-E557. [92] Raica, M., Coculescu, M., Cimpean, M. and Ribatti, D. (2010) Endocrine gland derived-VEGF is downregulated in human pituitary adenoma. Anticancer Research, 30, 3981-3986. [93] Levit, A., Yarnitzky, T., Wiener, A., Meidan, R. and Niv, M.Y. (2011) Modeling of human prokineticin receptors: Interactions with novel small molecules binders and po- tential off-target drugs. PLoS One, 6, e27990.1-17. [94] Alvarez, C., Alonso-Muriel, I., Garcia, G., Crespo, J., Bellever, J., Simon, C. and Pellicer, A. (2007) Implanta- tion is apparently unaffected by the dopamine agonist Cabergoline when administered to prevent ovarian hyper- stimulation syndrome in women undergoing assisted re- production treatment: A pilot study. Human Reproduction, 22, 3210-3214. doi:10.1093/humrep/dem315 [95] Sykiotis, G.P., Plummer, L., Hughes, V.A., Au, M., Dur- rani, S., Young, S.N., Dwyer, A.A., Quinton, R., Hall, J.E., Gusella, J.F., Seminara, S.B., Crowley Jr., W.F. and Pitteloud, N. (2010) Oligogenic basis of isolated gonado- tropin-releasing hormone deficiency. Proceedings of the National Academy of Sciences of the United States of America, 107, 15140-15144. doi:10.1073/pnas.1009622107
|