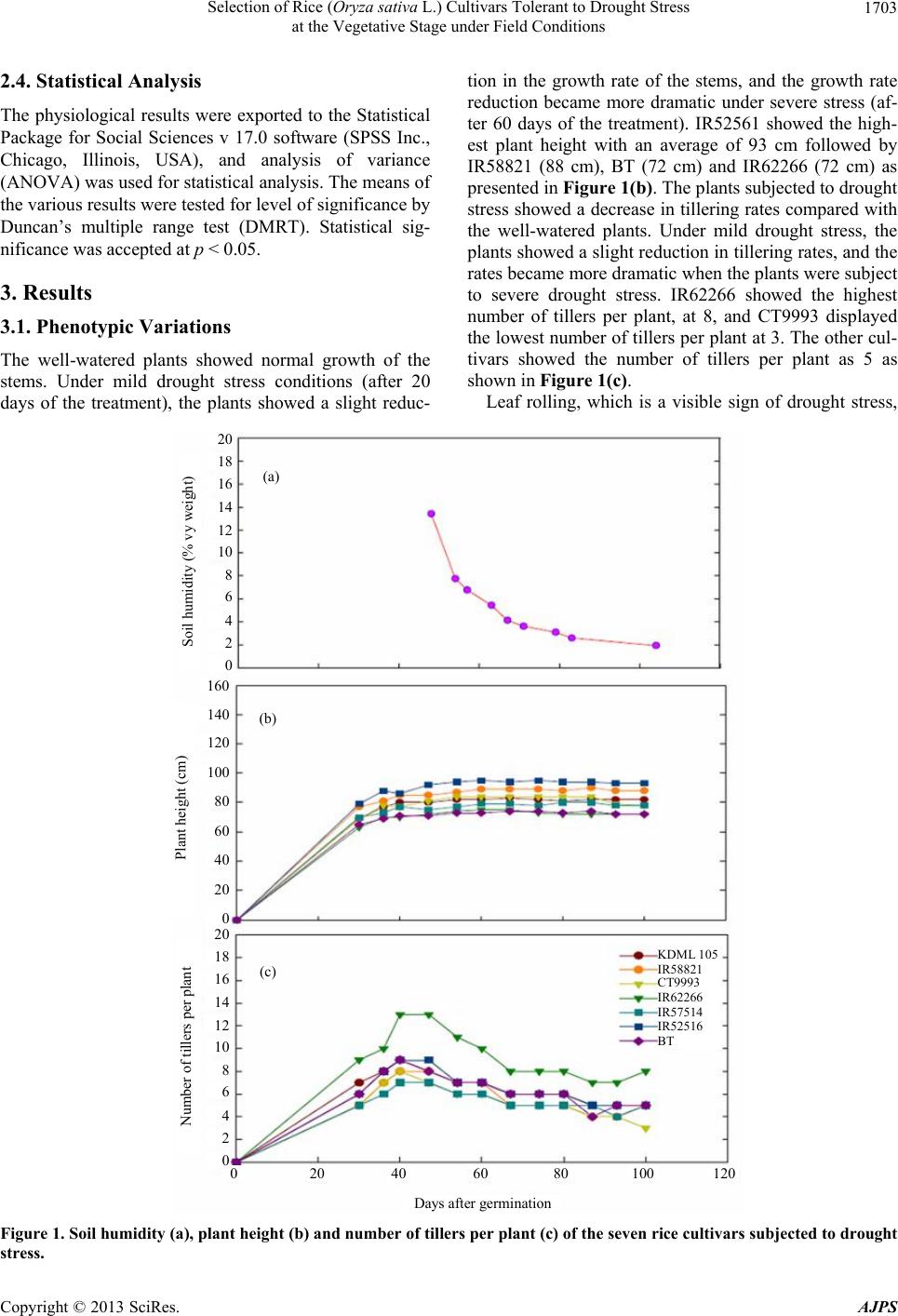
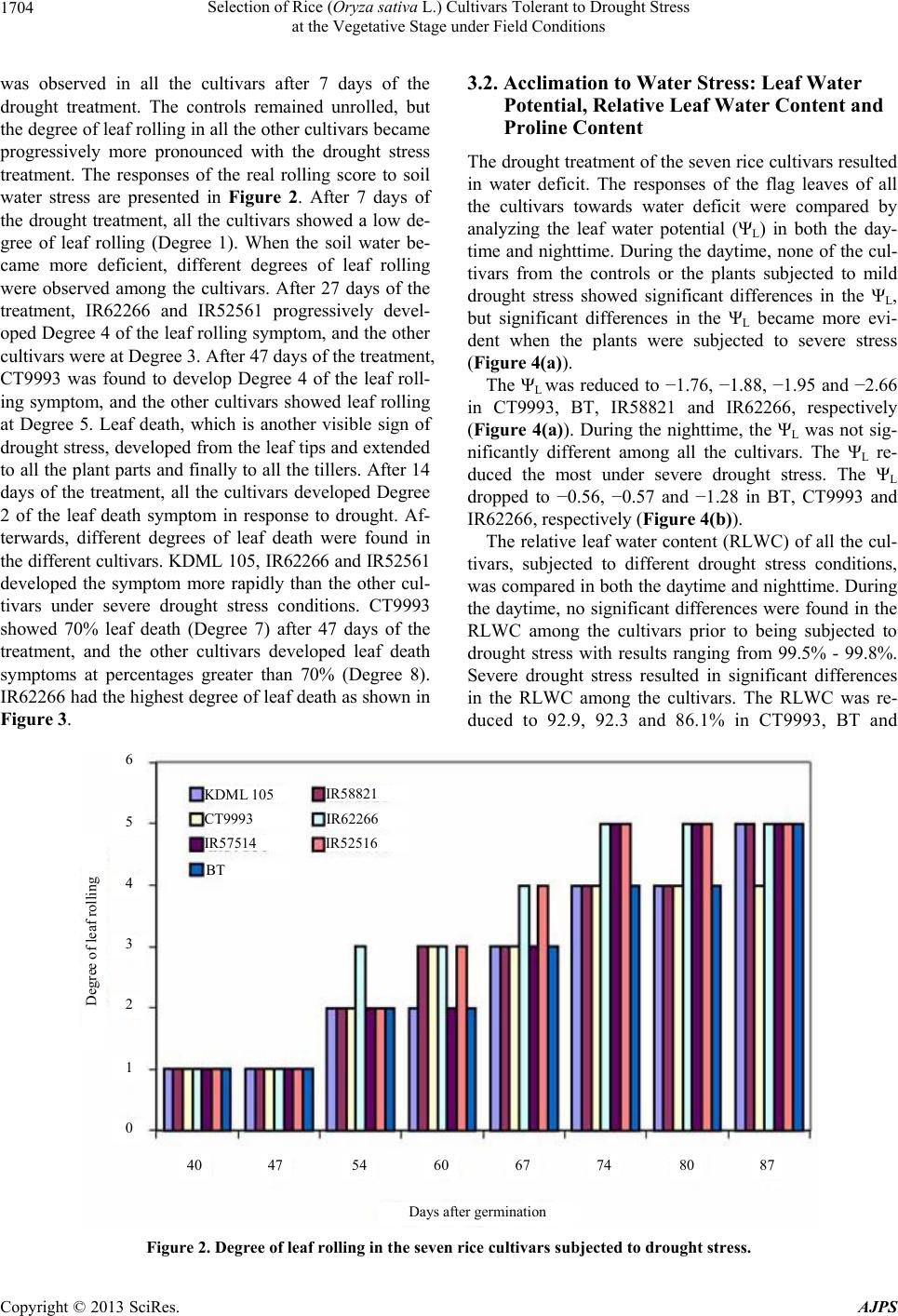
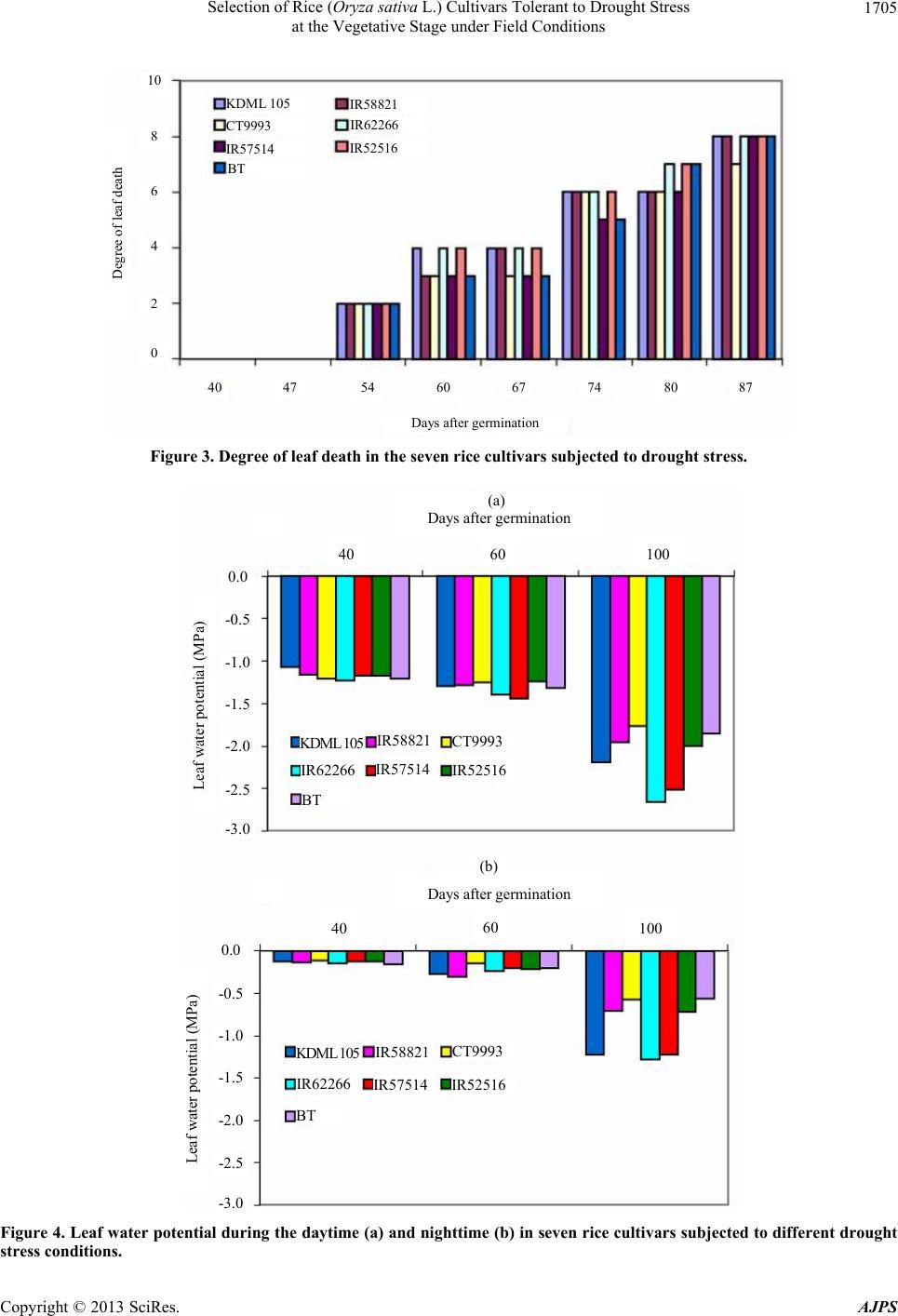
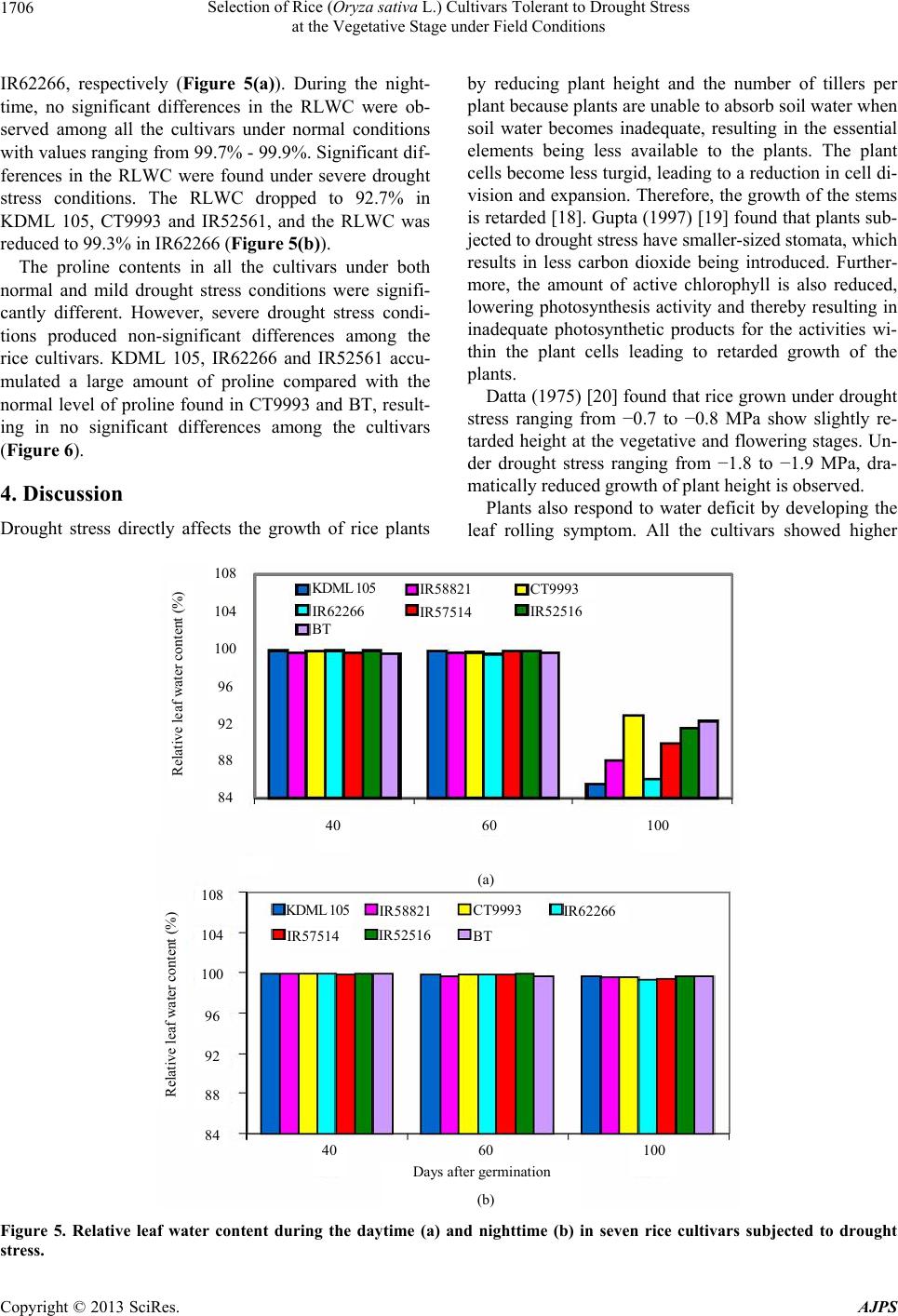
Selection of Rice (Oryza sativa L.) Cultivars Tolerant to Drought Stress
at the Vegetative Stage under Field Conditions
1708
[2] R. N. Roy and R. V. Misra, “Economic and Environ-
mental Impact of Improved Nitrogen Management in
Asian Rice-Farming Systems,” Proceedings of the 20th
Session of the International Rice Commission, 2002, pp.
23-26.
[3] OAE (Office of Agricultural Economics), “Report on
Survey of Rice Growing Areas in Thailand Crop Year
2012/2013,” Center for Agricultural Information, Office
of Agricultural Economics, Ministry of Agriculture and
Cooperation, Bangkok, 2013.
[4] B. Jongdee, “Designing a National Breeding Program for
Developing Drought-Tolerant Rainfed Lowland Varieties:
The Thailand Experience,” In: K. S. Fischer, R. Lafitte, S.
Fukai, G. Atlin and B. Hardy, Eds., Breeding Rice for
Drought-Prone Environments 64-69, International Rice
Research Institute, Los Banõs, The Philippines, 2003.
[5] L. J. Wade, C. G. McLaren, L. Quintana, D. Harnpichit-
vitaya, S. Rajatasereekul, A. K. Sarawgi, A. Kuma, H. U.
Ahmed, A. K. Sarwoto, R. Singh, J. Rodriguez, Sio-
pongco and S. Sarkarung, “Genotype by Environment In-
teractions across Diverse Rainfed Lowland Rice Envi-
ronments,” Field Crops Research, Vol. 64, No. 1, 1999,
pp. 35-50. doi:10.1016/S0378-4290(99)00049-0
[6] P. J. Kramer and J. S. Boyer, “Water Relations of Plant
and Soil,” Academic Press, San Diego, 1995.
[7] L. D. Nooden, “The Phenomena of Senescence and Ag-
ing,” In: L. D. Nooden and A. C. Leopald, Eds., Senes-
cence and Aging in Plants, Academic Press, San Diego,
1988, pp. 1-50.
[8] J. M. Hansen, Y. M. Go and D. P. Jones, “Nuclear and
Mitochondrial Compartmentation of Oxidative Stress and
Redox Signaling,” Annual Review of Pharmacology and
Toxicology, Vol. 46, 2006, pp. 215-34.
doi:10.1146/annurev.pharmtox.46.120604.141122
[9] B. Jongdee, “New Rice Breeding Methods for Rainfed
Lowlands of North and Northeast Thailand,” Proceedings
of an International Workshop: Increased Lowland Rice
Production in the Mekong Region, Vientiane, 30 Octo-
ber-2 November 2000, pp. 221-228.
[10] G. Pantuwan, S. Fukai, M. Cooper, S. Rajatasereekul and
J. C. O’Toole, “Yield Response of Rice (Oryza sativa L.)
Genotypes to Drought under Rainfed Lowlands 2. Selec-
tion of Drought Resistant Genotypes,” Field Crops Re-
search, Vol. 73, No. 2-3, 2002, pp. 169-180.
doi:10.1016/S0378-4290(01)00195-2
[11] A. Price and B. Courtois, “Mapping QTLs Associated
with Drought Resistance in Rice: Progress, Problems and
Prospects,” Plant Growth Regulation, Vol. 29, 1999, pp.
123-133. doi:10.1023/A:1006255832479
[12] B. Jongdee, “Designing a National Breeding Program for
Developing Drought Tolerant Rainfed Lowland Varieties:
The Thailand Experiences,” In: K. S. Fischer, R. Lafitte,
S. Fukai, G. Atlin and B. Hardy, Eds., Breeding Rice for
Drought-Prone Environments, International Rice Research
Institute, Los Banõs, 2004. pp. 64-69.
[13] G. Pantuwan, S. Fukai, M. Cooper, S. Rajatasereekul and
J. C. O’Toole, “Yield Response of Rice (Oryza sativa L.)
Genotypes to Drought under Rainfed Lowlands. 1. Grain
Yield and Yield Components,” Field Crop Research, Vol.
73, 2001, pp. 153-168.
doi:10.1016/S0378-4290(01)00187-3
[14] J. C. O’Toole and R. Cruz, “Response of Leaf Water
Potential, Stomatal Resistance and Leaf Rolling to Water
Stress,” Plant Physiology, Vol. 65, No. 3, 1980, pp. 428-
432. doi:10.1104/pp.65.3.428
[15] IRRI, “Annual Report for 1975,” International Rice Re-
search Institute, Los Banõs, 1976, p. 418.
[16] N. C. Turner, “Techniques and Experimental Approaches
for the Measurement of Plant Water Status,” Plant Soil,
Vol. 58, No. 1, 1981, pp. 339-366.
doi:10.1007/BF02180062
[17] L. S. Bates, R. P. Waldren and I. D. Teare, “Rapid De-
termination of Free Proline for Water Stress Studies,”
Plant and Soil, Vol. 39, No. 1, 1973, pp. 205-208.
doi:10.1007/BF00018060
[18] T. C. Hsiao, J. C. O’Toole, E. B. Yambao and N. C.
Turner, “Influence of Osmotic Adjustment on Leaf Roll-
ing and Tissue Death in Rice (Oryza sativa L.),” Plant
Physiology, Vol. 75, No. 1, 1984, pp. 338-341.
doi:10.1104/pp.75.2.338
[19] U. S. Gupta, “Crop Improvement: Stress Tolerance,” Sci-
ence Publishers, Inc., New Hamphire, 1997.
[20] S. K. Datta, “Drought Tolerance in Rice in Relation to
Growth Stages,” IRRI, Los Banõs, 1975.
[21] M. Dingkuhn, R. T. Cruz, C. O. O’Toole, N. C. Turner
and K. Doerffling, “Responses of Seven Diverse Rice
Cultivars to Water Deficits. III. Accumulation of Abscisic
Acid and Proline in Relation to Leaf Water Potential and
Osmotic Adjustment,” Field Crops Research, Vol. 27, No.
1-2, 1991, pp. 103-117.
doi:10.1016/0378-4290(91)90025-Q
[22] A. Sigari, L. J. Wadel and A. Yamauch, “Response of
Roots Traits of Rainfed Lowland Rice to Drought and
Rewatering,” In: J. Abe and S. Morita, Eds., Root System
Management that Leads to Maximize Rice Yields, JSRR,
Tokyo, 1997.
[23] R. Munns, “Comparative Physiology of Salt and Water
Stress,” Plant, Cell and Environment, Vol. 25, No. 2,
2002, pp. 239-250.
doi:10.1046/j.0016-8025.2001.00808.x
[24] H. T. Nguyen, R. C. Babu and A Blum, “Breeding for
Drought Resistance in Rice: Physiology and Molecular
Genetics Considerations,” Crop Science, Vol. 37, No. 5,
1997, pp. 1426-1434.
doi:10.2135/cropsci1997.0011183X003700050002x
Copyright © 2013 SciRes. AJPS