G. Strippoli et al. / Agricultural Sciences 4 (2013) 376-381
380
ences were observed between Coratina and Arbequina
for growth, nodes number and internodes mean length of
sylleptic shoots (4.0 cm, 3.1 - 3.6, 12.9 - 11.5 mm, re-
spectively).
As shoot development was the result of the two com-
plementary phases, known as organogenesis and disten-
sion [29], consequently in the climatic and cultural con-
ditions under study these phases should be in general
concentrated and sped up in spring, particularly for the cv.
Arbequina.
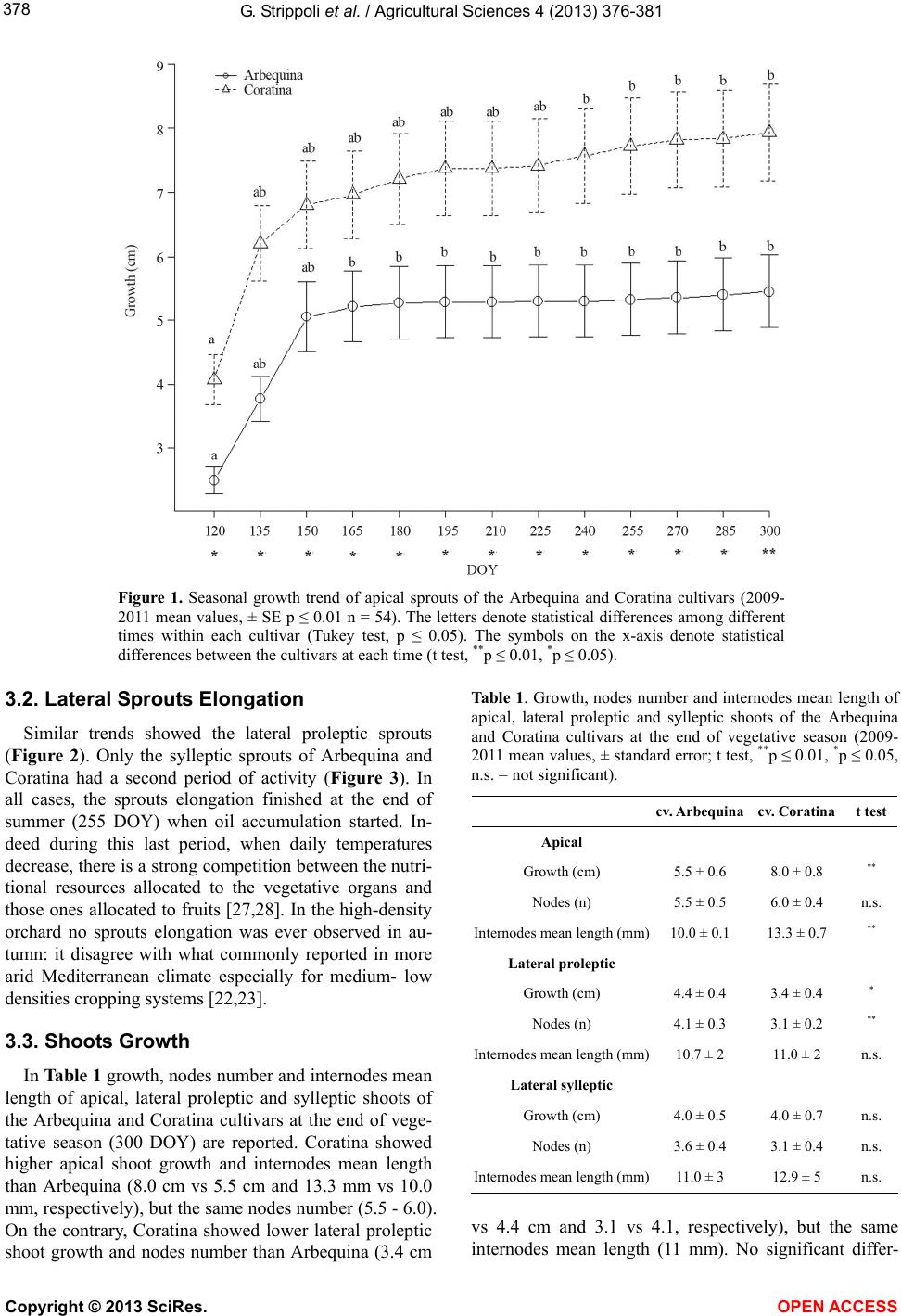
At all measurement times, Coratina showed ever sig-
nificant higher apical sprouts growth values with respect
to those ones of Arbequina (Figure 1). On the contrary,
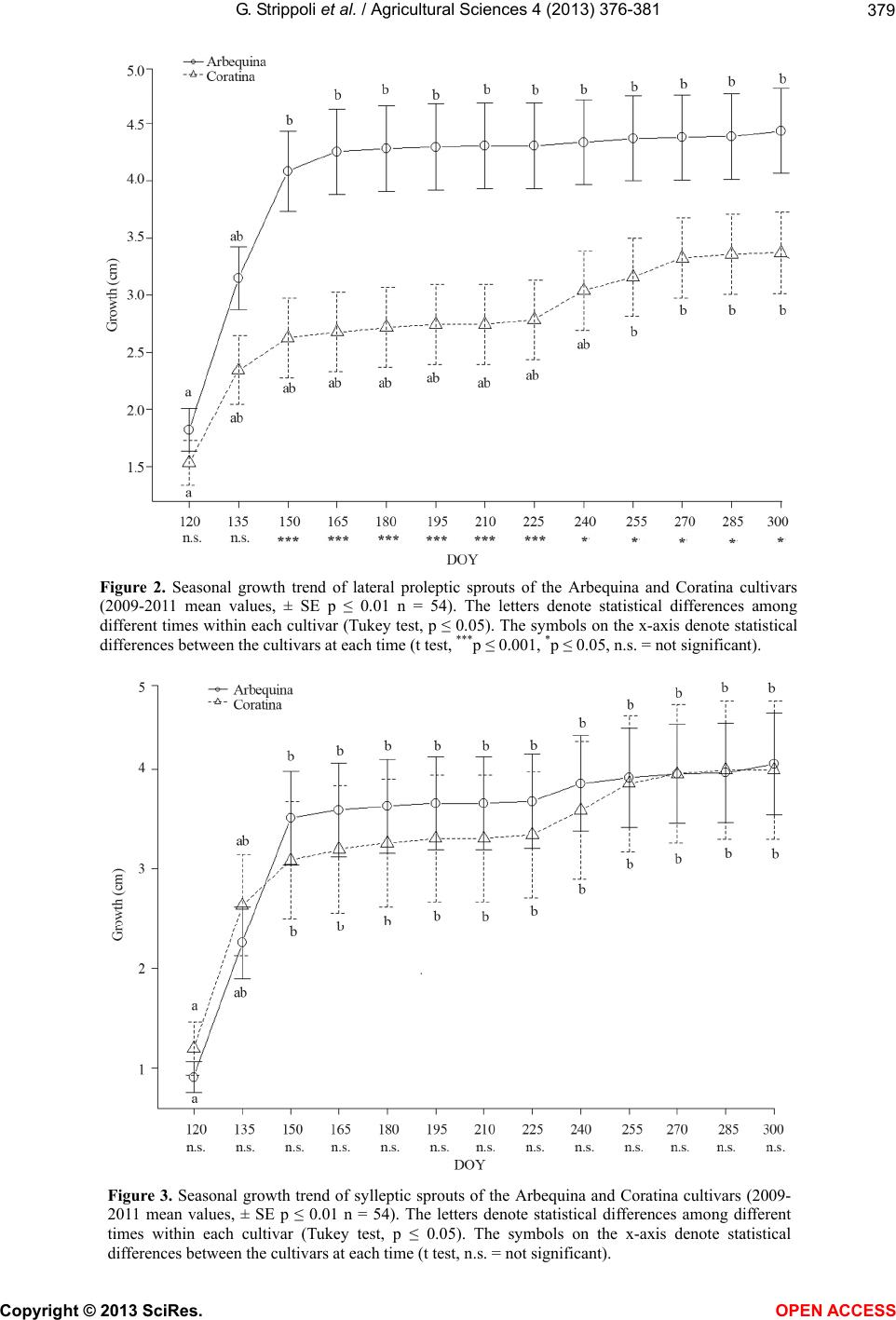
Arbequina showed significant higher lateral proleptic
sprouts elongations with respect to those ones of Corat-
ina, except for the first two times (Figure 2). Finally, no
different sylleptic sprouts growth values were observed
between the cultivars at all measurement times (Figure
3).
The differences observed between the two cultivars
could be explained considering their different vigour,
already stated in the literature: Coratina is a cultivated
variety with a medium vigour, while Arbequina is a low-
vigour genotype [1,2,4].
4. CONCLUSION
These are the first data on seasonal elongation of all
types of sprouts (apical, lateral proleptic and sylleptic) on
an olive shoot under high-density irrigated conditions.
The introduction of this innovative cropping system al-
lows a considerable reduction of production costs. In
traditional olive groves most of the production costs are
definitely linked to the farming operations carried out
manually. These operations are fully removed in this
context as performed by continuous machines response-
ble for the management of the olive. The result is a con-
siderable increase in the economic performance of the
olive grove and a consequent reduction in the unit cost
for kg of oil [30]. These data are very useful first of all
for varietal choice and field management, such as fertili-
zation and pruning not only in high-density orchards, and
then for new olive breeding programs.
5. AKNOWLEDGEMENTS
This research had the financial support of the National Research
Program (PRIN-MIUR, Project “Biological processes and environmen-
tal factors involved in the vegetative growth, fruiting and oil quality
control in superintensive olive (Olea europaea L.) plantings”.
REFERENCES
[1] Camposeo, S., Ferrara, G., Palasciano, M. and Godini, A.
(2008) Varietal behaviour according to the superintensive
olive culture training system. Acta Horticulturae, 791,
171-274.
[2] Godini, A., Vivaldi, G.A. and Camposeo, S. (2011) Olive
cultivars field-tested in super high-density system in
southern Italy. California Agriculture, 65, 39-40.
doi:10.3733/ca.v065n01p39
[3] Contò, F. (2004) Optimizing the use of water and envi-
ronmental resources fora model of sustainable develop-
ment of the “Basso Tavoliere” Area. Edizione L’Aquilone,
Potenza.
[4] Camposeo, S. and Godini, A. (2010) Preliminary obser-
vations the performance of 13 varieties according to the
super high density oliveculture training system in Apulia
(southern Italy). Advanced Horticultural Science, 24,
16-20.
[5] Connor, D.J. and Gómez-del-Campo, M. (2013) Simula-
tion of oil productivity and quality of N-S oriented olive
hedgerow orchards in response to structure and intercep-
tion of radiation. Scientia Horticulturae, 150, 90-99.
doi:10.1016/j.scienta.2012.09.032
[6] Camposeo, S. and Vivaldi, G.A. (2011) Short-term effects
of de-oiled olive pomace mulching application on a
young super high-density olive orchard. Scientia Horti-
culturae, 129, 613-621.
doi:10.1016/j.scienta.2011.04.034
[7] Gómez-del-Campo, M. (2013) Summer deficit-irrigation
strategies in a hedgerow olive orchard cv. “Arbequina”:
Effect on fruit characteristics and yield. Irrigation Sci-
ence, 31, 259-269. doi:10.1007/s00271-011-0299-8
[8] Vivaldi, G.A., Strippoli, G. and Camposeo, S. (2013)
Ecophysiological response to irrigation of two olive cul-
tivars grown in a high-density orchard. Agricultural Sci-
ences (in press).
[9] Camposeo, S., Vivaldi, G.A. and Gattullo, C.E. (2013)
Ripening indices and harvesting times of different olive
cultivars for continuous harvest. Scientia Horticulturae,
151, 1-10. doi:10.1016/j.scienta.2012.12.019
[10] Tombesi, A., Proietti, P., Iacovelli, G., Tombesi, S. and
Farinelli, D. (2011) Vegetative and productive behaviour
of four olive Italian cultivars and “Arbequina” according
to super intensive olive training system in central Italy.
Acta Horticulturae, 924, 211-218.
[11] Goethe, J.W. (1790) Versuch die metamorphose der
pflanzen zu erklären. Modern Edition, La Metamorphose
des Plantes (1975), Triades, Paris.
[12] Goebel, K. (1900) Organography of plants. Part I. Gen-
eral organography (translated by IB Balfour). The Clar-
endon Press, Oxford.
[13] Oldeman, R.A.A. (1974) L’architecture de la fôret guy-
anaise. O.R.S.T.O.M., Paris.
[14] Hallé, F. and Oldeman, R.A.A. (1970) Essai sur l’archi-
tecture et la dynamique de croissance des arbres tropicaux.
Masson, Paris.
[15] Edelin, C., (1981) Quelques aspects de l’architecture
vegetative des Conifères. Bulletin de la Sociètè Botanique
de France, Lettres Botanique de France. Lettres
Botaniques, 128, 177-188.
Copyright © 2013 SciRes. OPEN ACCESS