M. Raspanti / J. Biomedical Science and Engineering 3 (2010) 1169-1174 1173
find its way into histology textbooks. We must take care
not to repeat the original sin of studying only tendons,
and then ascribe what we found here to all other tissues.
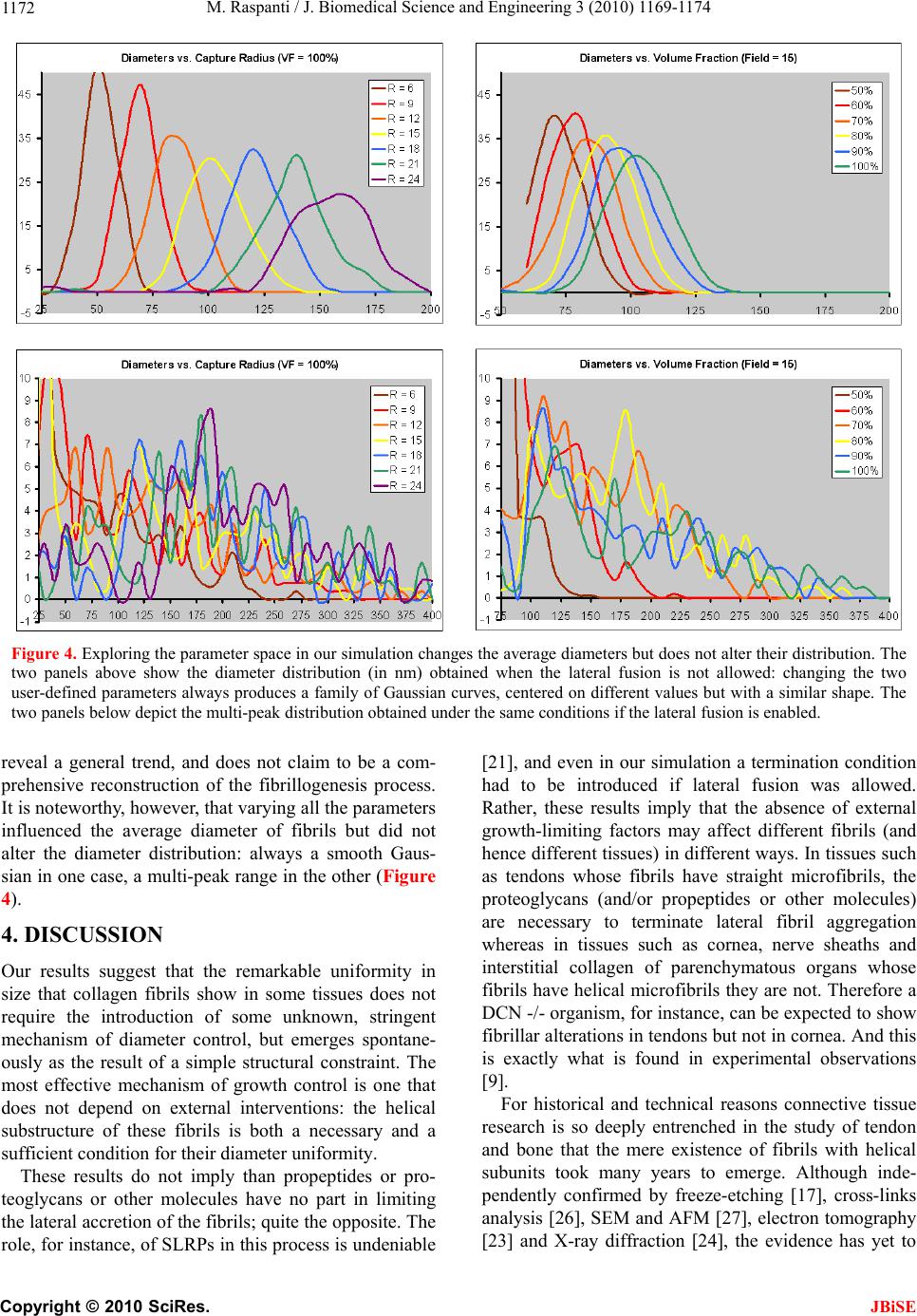
Fibrils with helical subfibrils are the most common
among our different tissues. Our data indicate that the
current knowledge on the fibrillogenesis process, all
gained on tendons, does not apply to these fibrils, and
that new research is required.
The next primary objective will be the identification
of the molecular switch controlling the initiation of one
or the other subfibrillar architecture, and which has so
far eluded detection. Fibrillogenesis is a delicate process
which can be influenced by myriad causes, some of
which do not even remain to make up the final product.
Perhaps the most promising candidate appears now
type V collagen. Almost since its discovery the type V
has associated with the control of fibril diameter [28],
and more recently it seems to be essential for fibril for-
mation in vivo: homozygote engineered organisms lack-
ing type V die at embryonic day 10 of cardiovascular
failure, while heterozygotes are viable but lack about
half their collagen fibrils [29].
It seems therefore not unreasonable to hypothesize
that type V collagen is involved in the formation of
slender, uniform fibrils in cornea, blood vessels and
sheaths, but obviously not in tendon and ligaments
where it appears only in traces. Further research is un-
derway.
REFERENCES
[1] Ottani, V., Martini, D., Franchi, M., Ruggeri, A. and
Raspanti, M. (2002) Hierarchical structures in fibrillar
collagens. Micron, 33, 587-596.
[2] Parry, D.A.D. and Craig, A.S. (1984) Growth and devel-
opment of collagen fibrils in connective tissue. In: Rug-
geri, A. and Motta, P.M., Eds., Ultrastructure of the
Connective Tissue Matrix, Martinus Nijhoff, Hague, tig
36-64.
[3] Ottani, V., Raspanti, M. and Ruggeri, A. (2001) Collagen
structure and functional implications. Micron, 32,
ons matures. Developmental Dynamics,
. Journal of Biological
orly
nces in
ent. Molecular and Cellular Biol-
and
don. Journal of
ive Oph-
gen. Journal of Biological
of collagen deposition. Nature
Journal of Biological
can.
enesis. Journal of Inves-
y. Journal of Ultra-
nce. Journal of Submi-
al Journal of
meter. European Journal of Cell Bi-
251–260.
[4] Birk, D.E., Zycband, E.I., Woodruff, S., Winkelmann,
D.A. and Trelstad, R.L. (1997) Collagen fibrillogenesis
in situ: fibril segments become long fibrils as the devel-
oping tend208, croscopic Cytology, 13, 135-143.
[19] Raspanti, M., Ottani, V. and Ruggeri, A. (1989) Different
architectures of the collagen fibril: morphological aspects
and functional implications. Internation
291-298.
[5] Chanut-Delalande, H., Fichard, A., Bernocco, S., Gar-
rone, R., Hulmes, D.J.S. and Ruggiero, F. (2001) Control
of heterotypic fibril formation by collagen V is deter-
mined by chain stoichiometry
Chemistry, 276, 24352-24359.
[6] Bornstein, P. (2002) The NH(2)-terminal propeptides of
fibrillar collagens: highly conserved domains with po
understood functions. Matrix Biology, 21, 217-226.
[7] Eyre, D.R., Weis, M.A. and Wu, J.J. (2008) “Adva
collagen cross-link analysis. Methods, 45, 65-74.
[8] Svensson, L., Aszodi, A., Heinegård, D., Hunziker, E.B.,
Reinholt, F.P., Fassler, R. and Oldberg, Å. (2002) Carti-
lage oligomeric matrix proteindeficient mice have nor-
mal skeletal developm
ogy, 22, 4366-4371.
[9] Danielson, K.G., Baribault, H., Holmes, D.F., Graham, H.,
Kadler, K.E. and Iozzo, R. (1997) Targeted disruption of
decorin leads to abnormal collagen fibril morphology
skin fragility. Journal of Cell Biology, 136, 729-743.
[10] Svensson, L., Aszodi, A., Reinholt, F.P., Fassler, R.,
Heinegård, D. and Oldberg, A. (1999) Fibromodulin-null
mice have abnormal collagen fibrils, tissue organization,
and altered lumican deposition in ten
Biological Chemistry, 274, 9636-9647.
[11] Chakravarti, S., Petroll, W.M., Hassell, J.R., Jester, J.V.,
Lass, J.H., Paul, J. and Birk, D.E. (2000) Corneal opacity
in lumican-null mice: defects in collagen fibril structure
and packing in the posterior stroma. Investigat
thalmology and Visual Science, 41, 3365-3373.
[12] Schonherr, E., Witsch-Prehm, P., Harrach, B., Robenek,
H., Rauterberg, J. and Kresse, H. (1995) Interaction of
biglycan with type I colla
Chemistry, 270, 2776-2783.
[13] Mao, J.R., Taylor, G., Dean, W.B., Wagner, D.R., Afzal,
V., Lotz, J.C., Rubin, E.M. and Bristow, J. (2002) Tenas-
cin-X deficiency mimics Ehlers-Danlos syndrome in
mice through alteration
Genetics, 30, 421-425.
[14] Kvist, A.J., Johnson, A.E., Mörgelin, M., Gustafsson, E.,
Bengtsson, E., Lindblom, K., Aszodi, A., Fassler, R., Sa-
saki, T. and Timpl, R. (2006) Chondroitin sulfate perle-
can enhances collagen fibril formation. Implications for
perlecan chondrodysplasias.
Chemistry, 281, 33127-33139.
[15] Wiberg, C., Klatt, A.R., Wagener, R., Paulsson, M.,
Bateman, J.F., Heinegård, D. and Mörgelin, M. (2003)
Complexes of matrilin-1 and biglycan or decorin connect
collagen VI microfibrils to both collagen II and aggre
Journal of Biological Chemistry, 278, 37698-37704.
[16] Bornstein, P., Kyriakides, T.R., Yang, Z., Armstrong, L.C.
and Birk, D.E. (2000) Thrombospondin 2 modulates col-
lagen fibrillogenesis and angiog
ative Dermatology, 5, 61-66.
[17] Ruggeri, A., Benazzo, F. and Reale, E. (1979) Collagen
fibrils with straight and helicoidal microfibrils: a
freeze-fracture and thin section stud
structure Research, 68, 101-108.
[18] Reale, E., Benazzo, F. and Ruggeri, A. (1989) Differ-
ences in the microfibrillar arrangement of collagen fibrils.
Distribution and possible significa
Biological Macromolecules, 11, 367-371.
[20] Birk, D.E. and Mayne, R. (1997) Localization of colla-
gen types I, III and V during tendon development.
Changes in collagen types I and III are correlated with
changes in fibril dia
ology, 72, 352-361.
[21] Raspanti, M., Viola, M., Sonaggere, M., Tira, M.E. and
Tenni, R. (2007) Collagen fibril structure is affected by
Copyright © 2010 SciRes. JBiSE