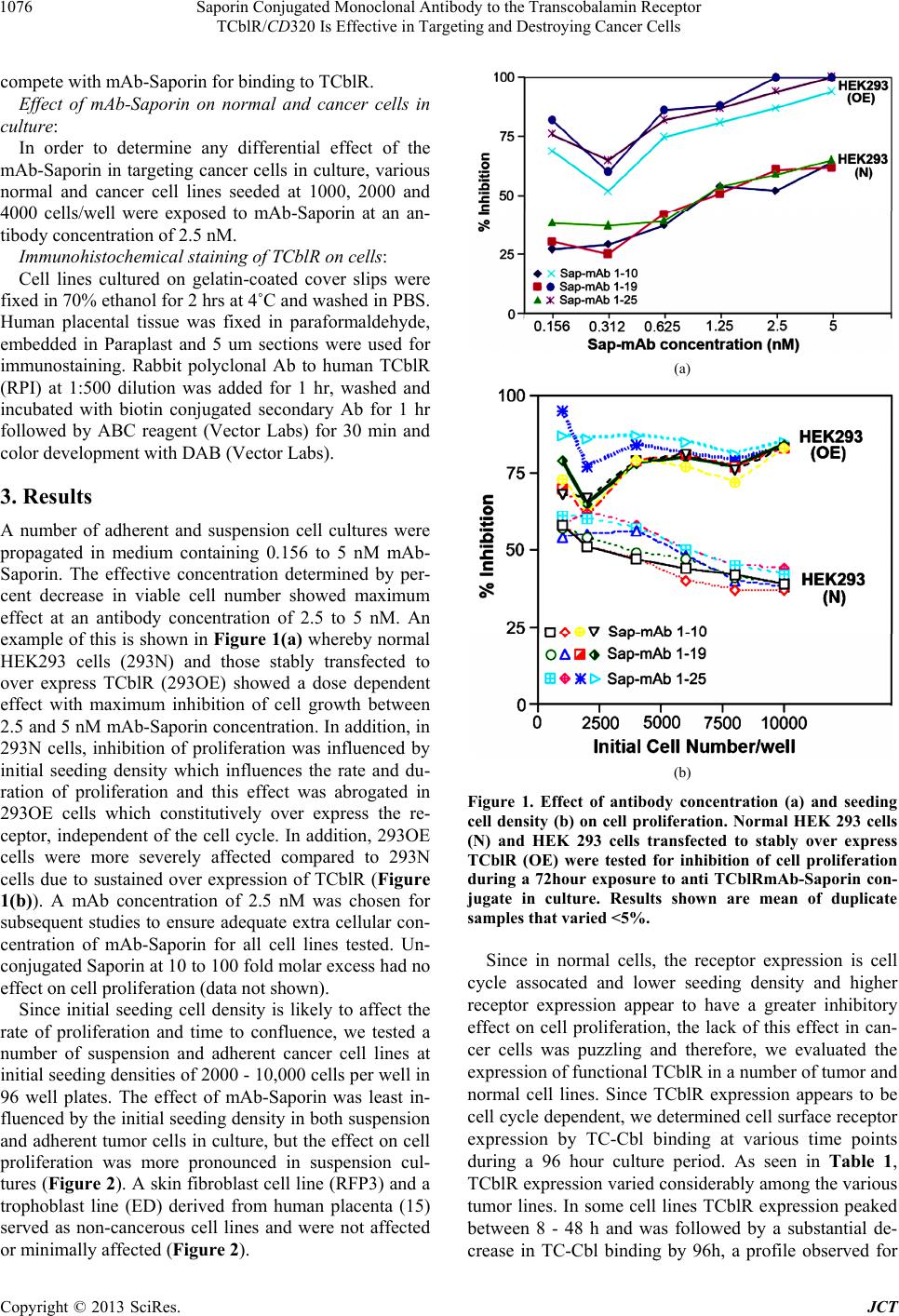
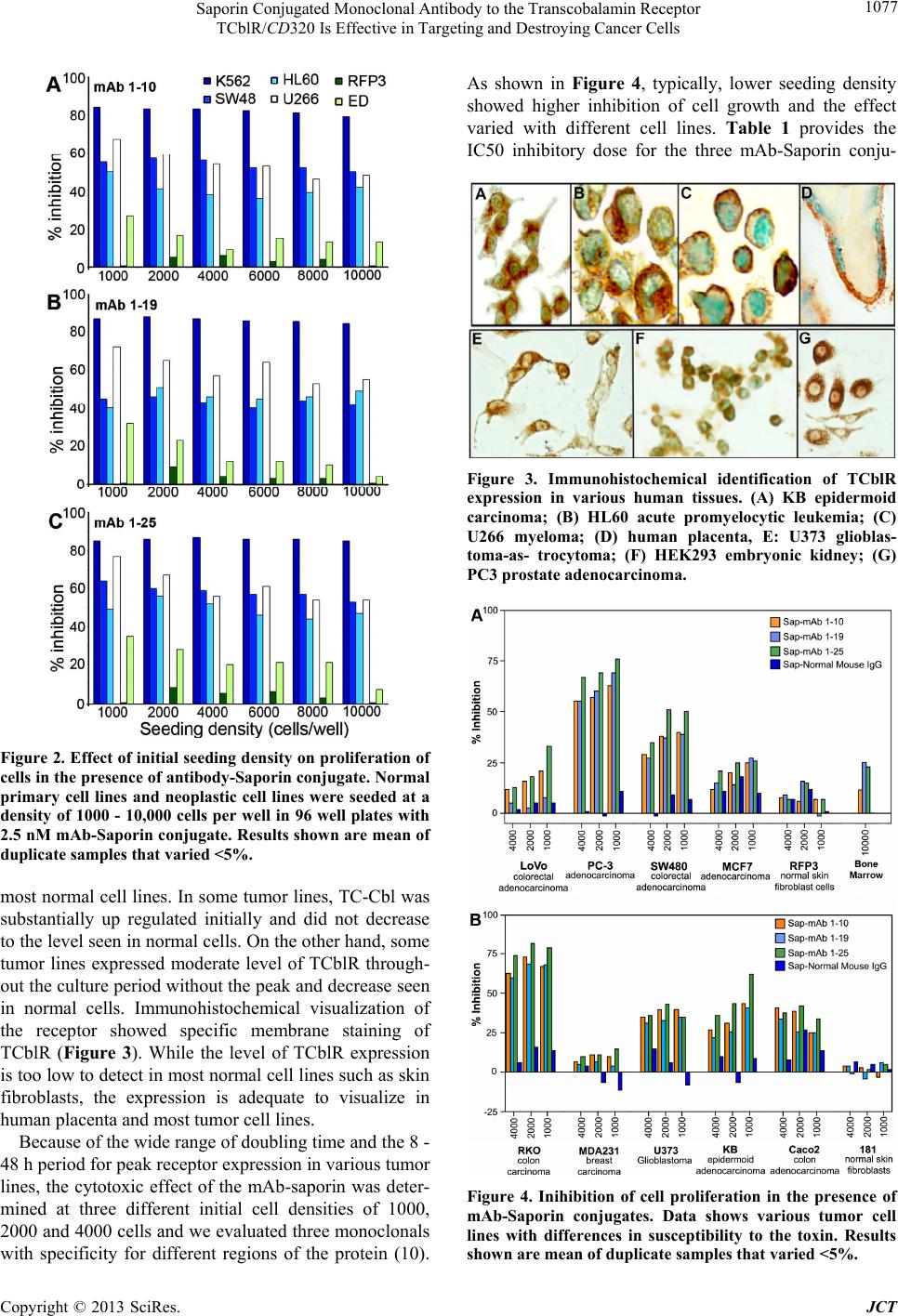
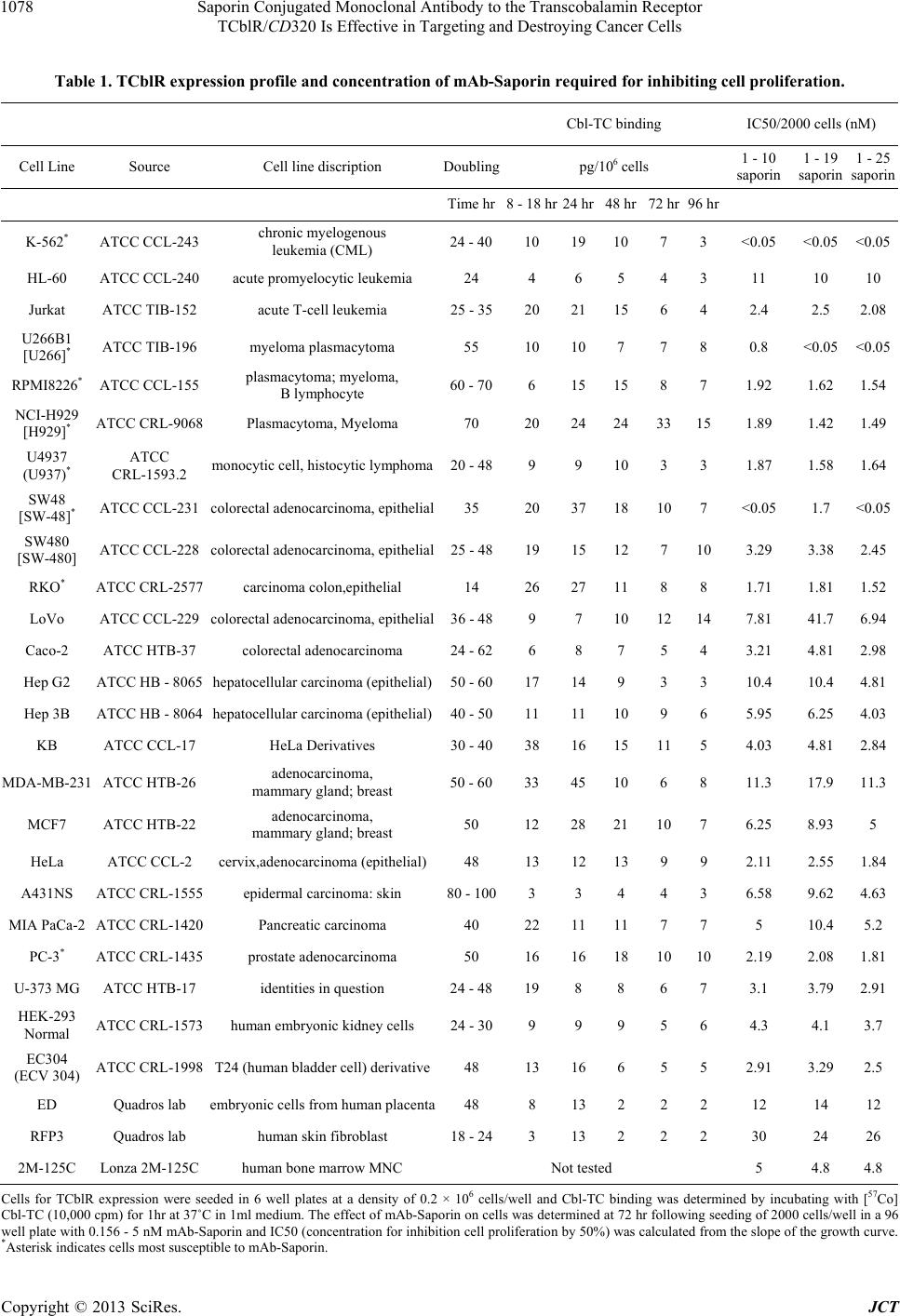
Saporin Conjugated Monoclonal Antibody to the Transcobalamin Receptor
TCblR/CD320 Is Effective in Targeting and Destroying Cancer Cells
1080
tional in recognizing the target antigen with high affinity,
an important requirement for mAbs to be used as carriers
of drugs and toxins. Even though a marginal increase in
internalization of mAb 1 - 10 and 1 - 19 is observed with
TC-Cbl in the culture medium, the binding of the native
ligand is not a pre requisite for antibody binding and in-
ternalization. Since in the absence of TC-Cbl, the apo re-
ceptor remains on the plasma membrane, TC-Cbl binding
appears to trigger the necessary response for internaliza-
tion. The effective binding and internalization of all an-
tibodies, including the ligand blocking antibody, mAb 1 -
25 is strong evidence that antibody binding triggers a
response similar to that of ligand TC-Cbl binding. Highly
potent toxins such as ricin, cholera toxin, gelonin and
saporin, drugs or radionuclides are very effective in de-
stroying cancer cells if a toxic dose can be delivered spe-
cifically to these cells [16,17]. This strategy requires a
tumor specific carrier to transport the toxin across the
plasma membrane into cells since these molecules cannot
cross the cell membrane by either specific or nonspecific
transport mechanisms. An ideal target protein would be a
receptor or cell surface protein that is expressed and in-
ternalized only in cancer cells. However, such proteins
are scarce and not easy to identify. Many proteins and
receptors are expressed in all cell types and some of
these are cell cycle associated or expressed only in ac-
tively dividing cells. Such proteins can be carriers for
drugs and toxins and may provide some degree of en-
hanced targeting to cancer cells. However selective tar-
geting to cancer cells and lack of toxicity to the normal
cell population will depend not only on the differential
expression but also on the density of the target protein in
the two cell types. For example, a protein with relatively
high expression such as the transferrin receptor [18],
even though differentially expressed in cancer and nor-
mal cells, would not be suitable for delivering a toxin
because the normal cells would internalize sufficient
toxin to kill the cell. The ideal target protein is one with
fairly low expression in normal cells and cannot inter-
nalize toxic amounts of drugs but is adequately over ex-
pressed in cancer cells to internalize cytotoxic amounts
of the drug. The TCblR is one such protein whose ex-
pression is sufficiently low to render any toxin internal-
ized in normal cells to be ineffective and is adequately
over expressed in some cancers to internalize sufficient
toxin to kill the cell. In addition, the cell cycle associated
expression of this protein makes highly proliferative
cancer cells with sustained expression, an ideal target for
this approach. Blocking Cbl uptake into cells with mono-
clonal antibody to TC can deplete cells of Cbl and ulti-
mately inhibit DNA synthesis leading to inhibition of cell
replication [19,20]. A specific antibody to TCblR that
blocks the binding of TC-Cbl could also have the same
effect [10]. This approach, even though less toxic, is
likely to be slow and many cancers require a faster effect
to destroy the malignant tissue before it metastasizes.
Inhibiting CD320 expression with siRNA also affects
proliferation by depleting intracellular Cbl [21]. The use
of potent drugs or toxins conjugated to antibody that can
deliver the payload to its target antigen is a highly effec-
tive strategy that can provide the specificity and speed of
action demanded in cancer therapy. The present data on
the use of mAb-Saporin conjugate to target TCblR ap-
pears to be specific for certain cancers and provides
proof of concept for utilizing this receptor for targeted
delivery of drugs and toxins and awaits confirmation of
in vivo targeting efficacy of this pathway.
5. Acknowledgements
This work was supported by NIH grant DK064732 to
EVQ and by KYTO Biopharma, Toronto, CA. EVQ and
JMS are inventors on patent applications US2011/
052154 and WO/2013/015821 by The Research Founda-
tion of SUNY. YN declared no potential conflicts.
REFERENCES
[1] E. V. Quadros, Y. Nakayama and J. M. Sequeira, “The
Protein and the Gene Encoding the Receptor for the Cel-
lular Uptake of Transcobalamin-Bound Cobalamin,” Blood,
Vol. 113, No. 1, 2009, pp. 186-192.
doi:10.1182/blood-2008-05-158949
[2] E. V. Quadros, S. P. Rothenberg and E. A. Jaffe, “Endo-
thelial Cells from Human Umbilical Vein Secrete Func-
tional Transcobalamin II,” American Journal of Physiol-
ogy, Vol. 256, No. 2, 1989, pp. C296-303.
[3] E. V. Quadros, Y. Nakayama and J. M. Sequeira, “The
Binding Properties of the Human Receptor for the Cellu-
lar Uptake of Vitamin B12,” Biochemical and Biophysi-
cal Research Communications, Vol. 327, No. 4, 2005, pp.
1006-1010. doi:10.1016/j.bbrc.2004.12.103
[4] K. Takahashi, M. Tavassoli and D. W. Jacobsen, “Rece-
ptor Binding and Internalization of Immobilized Transco-
balamin II by Mouse Leukaemia Cells,” Nature, Vol. 288,
No. 5792, 1980, pp. 713-715. doi:10.1038/288713a0
[5] T. Kishimoto, M. Tavassoli, R. Green and D. W. Jacob-
sen, “Receptors for Transferrin and Transcobalamin II
Display Segregated Distribution on Microvilli of Leuke-
mia L1210 Cells,” Biochemical and Biophysical Research
Communications, Vol. 146, No. 3, 1987, pp. 1102-1108.
doi:10.1016/0006-291X(87)90761-3
[6] E. V. Quadros, “Advances in the Understanding of Co-
balamin Assimilation and Metabolism,” British Journal of
Haematology, Vol. 148, No. 2, 2010, pp. 195-204.
doi:10.1111/j.1365-2141.2009.07937.x
[7] J. Lindemans, A. C. Kroes, J. van Geel, J. van Kapel, M.
Schoester, et al., “Uptake of Transcobalamin II-Bound
Cobalamin by HL-60 Cells: Effects of Differentiation In-
Copyright © 2013 SciRes. JCT