Open Journal of Ecology
Vol.2 No.3(2012), Article ID:21613,10 pages DOI:10.4236/oje.2012.23015
Factors associated with the spread of Chinese Tallow in East Texas forestlands
![]()
Forest and Wildlife Research Center, Mississippi State University, Starkville, USA; *Corresponding Author: zfan@cfr.msstate.edu
Received 11 May 2012; revised 14 June 2012; accepted 2 July 2012
Keywords: Chinese Tallow; Rate of Spread; CART; Logistic Regression; FIA
ABSTRACT
Chinese tallow tree (Triadica sebifera (L.) Small, Sapium sebiferum (L.) Roxb) is an invasive species that is replacing native ecosystems in areas of eastern Texas. It is imperative that the spatial pattern of the spread of this species be identified, as well as causal mechanisms. To that end, we seek to determine factors that contribute to the spread of Chinese tallow using classification and regression tree (CART) and logistic regression. We also attempt to identify current locations and spread rates across eastern Texas using Forest Inventory Analysis (FIA) data within major forest types. Distance to formerly infested plots and roads, slope, and disturbances (natural and anthropogenic) were identified as major (either facilitating or impeding) factors for the spread of Chinese tallow across the landscape. The highest probability of occurrence and spread rate of Chinese tallow were found in the oak/ gum/cypress forest type. Continued disturbance, from harvest events or natural disasters will allow the species to continue to spread throughout the region and could threaten overall forest productivity. We also discuss some implications of the continued spread of Chinese tallow on forest management. Forest managers could benefit from this analysis and use it as a guide for monitoring forest types with the highest risk of invasion.
1. INTRODUCTION
Chinese tallow tree (Triadica sebifera (L.) Small, Sapium sebiferum (L.) Roxb), native to Japan and central China, was introduced into the United States in the late 1700s as an ornamental and potential oil crop species [1]. It was introduced into the Gulf of Mexico coastal region by the USA Department of Agriculture for establishing local soap industries in the 1900s, and has continued to thrive along the Gulf Coast to present day [2]. Chinese tallow is an aggressive invader throughout forestlands along the Gulf Coast [3] and has become the fifth most common species in eastern Texas [4].
The tallow tree is in the Euphorbiaceae family and typically reaches maturity after three years. It is able to adapt to a variety of site conditions (shade to full sun) and soil types (acidic to alkaline, wet to dry) [5-6]. One plant can provide a considerable seed source (up to 100,000 seeds per year) that can be distributed by birds and water [7-8]. Seeds from Chinese tallow can germinate under stressful conditions and remain viable for many years; while it is unclear how long the trees remain viable for reproduction, it has been suggested it could approach 100 years [6,9]. After tallow trees are established in an area, their growth and reproduction are restricted by the local environmental conditions [10-12]. Competition with native species is an important factor to determine the invasibility of an area [13]. The competition between native species and Chinese tallow is for limited biological resources (e.g., light, nutrients, soil moisture) and physical space. Invasion of tallow trees is typical along forest edges and openings in coastal prairies and coastal plains [14]. In addition, the spread of tallow trees can be promoted in disturbed areas where ecosystem balance is dynamic and resources are available to create favorable habitat for colonization and establishment [15]. Along the Gulf Coast, land-falling hurricanes can have significant impacts on forested areas [16], exacerbating the invasion potential by Chinese tallow.
Tallow tree has become a major threat to coastal forests in Florida, Louisiana, Mississippi and Texas. Moreover, tallow tree is considered a transformer species [17] because it can change natural soil conditions and create a transformation of community structure from grassland to woodland [3,18]. However, tallow tree is largely confined to areas along the Gulf Coastal Plain. It has been suggested that tallow tree could survive in areas outside of its current range, as occurs in its native habitat ranges in China, and could expand up to 500 km from its current range [10]. Wang et al. [19] utilized Forest Inventory Analysis (FIA) data from the USA. Forest Service to simulate the expansion of Chinese tallow in Texas and Louisiana and estimate that over 1.5 million ha of forestlands could be occupied by the year 2023. Further, their study estimates that tallow tree could migrate over 300 km into areas as far north as 34˚N latitude within the next 115 years (100 years if temperatures increase by 2˚C) [19].
For invasive species research and management, it has been a challenge to understand the invasion process and associated driving factors, and design effective methods for controlling the spread of tallow tree and reducing ecological and economic loss. The objectives of this study are to 1) map the distribution of tallow tree in southeast Texas and 2) evaluate the impact of a suite of potential factors on the spread of tallow trees by analyzing FIA data using GIS and spatial statistics. The information will be useful to control and mitigate the spread of tallow trees in southern forested lands.
2. METHODS
2.1. Study Area and Inventory Data
East Texas occupies 45 counties and contains approximately 9.1 million ha, of which approximately 4.9 million ha are forested. Forests in east Texas are characterized by high productivity and species diversity [20]. Those forests which lie within the West Gulf Coastal Plain, cover six forest type groups: longleaf/slash pine, loblolly/shortleaf pine, oak/pine, oak/hickory, oak/gum/cypress, and Elm/ash/cottonwood (Figure 1). Loblolly/shortleaf pine forests occur most frequently in the study area, followed by oak/hickory and oak/gum/cypress forests. Oak/ hickory forests are mainly distributed in northern counties of east Texas, whereas oak/gum/cypress forests mostly cluster in southern counties close to the coast.
Beginning in 1999, the Forest Inventory and Analysis (FIA) program transitioned from a periodic inventory plan to an annual measurement scheme to monitor both annual and long-term forest change. All sample plots within a sampling/inventory unit are measured over a 5 - 7 years period with approximately 20% of the sample plots being measured each inventory year [21]. From 2001 to 2008, 2426 plots were measured in east Texas. Each FIA plot was spatially referenced by its latitude and longitude and identified with presence (1) or absence (0) of tallow tree. With infested FIA plots, the cover percentage of tallow tree was measured in each of the four subplots and was classified into four levels in this study: trace (<1%), low (1% ~ 10%), medium (10.1% ~ 50%) and high (>50%). In addition, stand and site factors associated with each FIA plot including forest type, stand origin (natural or artificial regeneration), ownership (private vs. public), elevation, slope, aspect, site productivity, physiographic class, stocking, and disturbance history (natural disturbances such as fire, flood, hurricane/tornado, human activities, etc.) were measured and/ or recorded based on the FIA protocols [21]. The distance of a FIA plot to the nearest highway, river, and infested plot was calculated by using GIS [22]. All variables
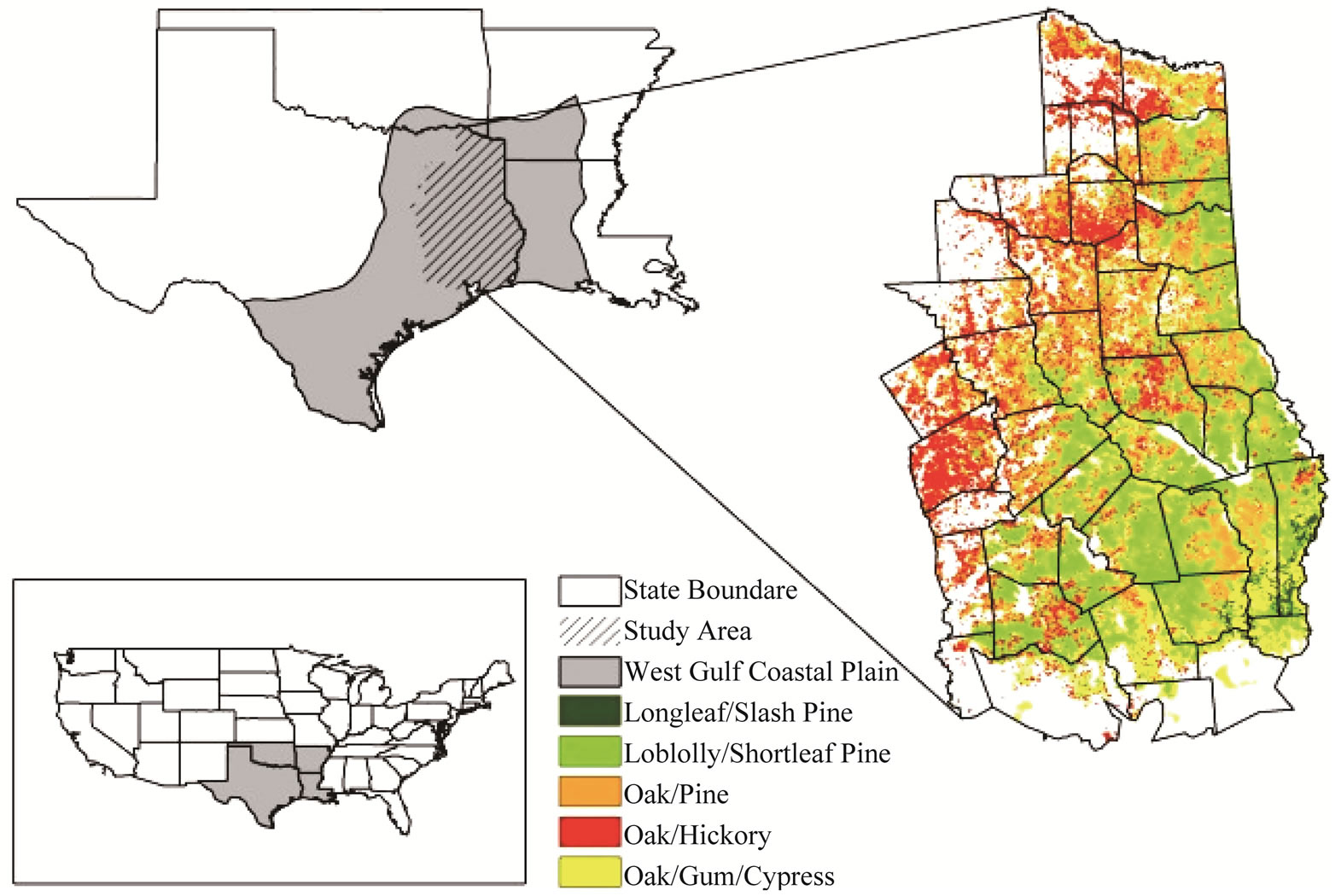
Figure 1. Map of study area, and associated forest types in Western Gulf Coastal Plain in East Texas.
collected for this study were grouped into three categories (i.e., disturbance-related, stand level and geographic variables) as defined in Table 1.
2.2. Regional Presence and Spread Patterns of Tallow Tree
From 2001 to 2008, 2426 FIA plots were measured in east Texas among which 454 plots were found to be infested by tallow tree with an overall probability of presence of 19%. Out of the 2426 FIA plots, 2224 plots were measured in both consecutive cycles (1999-2004, 2005- 2008), which were classified into four conditions (Table 2). To map the probability of presence of tallow tree in east Texas, all 2426 FIA plots were utilized through a Gaussian kernel density function. During 2004 through 2008, 190 out of 2224 FIA plots were found to be newly infested by tallow tree with an overall annual probability of spread of 1.8%. The same method (i.e., Gaussian kernel smoothing) was employed to map the probability of spread of tallow tree between 2004 and 2008 by using the 2224 repeat measured FIA plots. Fan et al. [23] described the details of using kernel smoothing to map the probability of presence and spread of invasive plants including the determination of best bandwidth of the Gaussian kernel density function. Meanwhile, both the newly infested and all infested FIA plots were extracted, separately by forest type group and the proportions of FIA plots falling within the four cover classes were calculated to reflect the presence and spread condition of tallow tree in different forest types/communities.
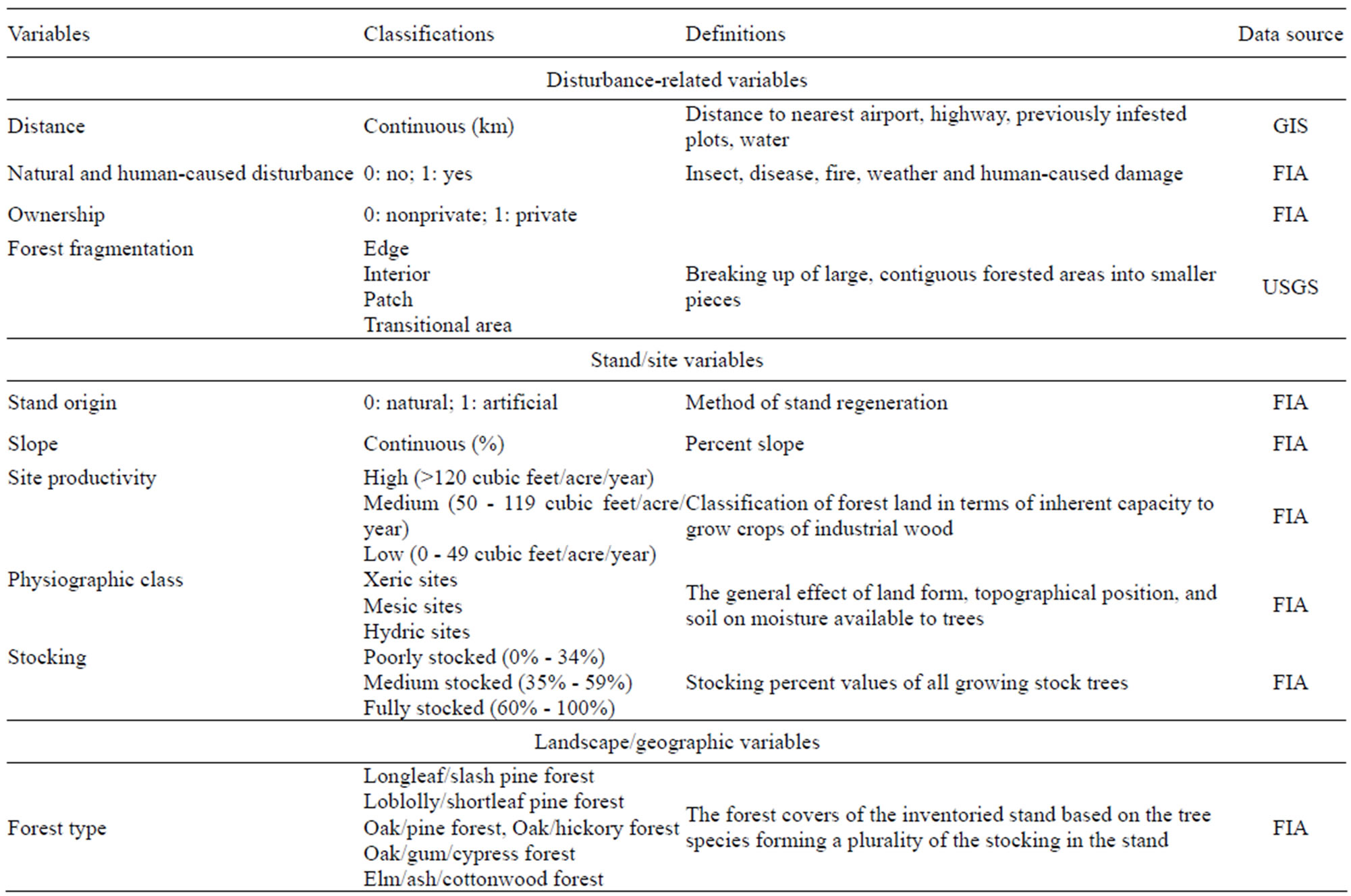
Table 1. Explanatory variables generated from FIA and USGS databases or calculated by using GIS for the CART and logistic regression model.

Table 2. Four conditions of tallow presence (1)/absence (0) in the remeasured plots.
2.3. Factors Associated with the Spread of Tallow Tree in Forested Land
Gan et al. [14] used logistic regression to link the probability of tallow tree occupation/presence to a set of factors which include landscape features, forest and site conditions, climatic condition, and natural and anthropogenic disturbances at the county level. Considering the non-stationary nature and spatial/temporal autocorrelation of the invasion process of invasive species, it is problematic to use a universal, non-spatial (i.e., spatial autocorrelation not being considered explicitly) regression model such as logistic regression to evaluate driving factor effect because spatial autocorrelation and factor effect may change dramatically over space [23]. Change in a driving factor over space is closely related to the scale or extent related to the dependent nature of the underlying processes it signifies. The distribution of tallow tree is spatially autocorrelated as it varies over space in a non-stationary manner. Furthermore, in recognition of presence/abundance of invasive species being the cumulative effect of natural and anthropogenic disturbances and landscape and site condition change over a long period of time, assessing the factors associated with the spread/infestation of Chinese tallow in forested land in east Texas will be accomplished by utilizing 1900 FIA plots (Out of which 137 were newly infested forest plots) within six forest type groups and measured in both inventory cycles. This allows for that any change occurred between the two inventory cycles to be accounted for through associated factors which should have a more direct relationship to this change. For this objective, a two step modeling scheme was implemented [24]. First we used the classification and regression tree (CART) to classify the study region into a set of homogenous, stationary sub-regions and identify factors which drive and/ or signify the large scale spread patterns of tallow tree. Then, a logistic regression (Eq.1) was conducted for each sub-region to analyze the relationship between the spread of tallow tree and associated factors at the subregional scale.
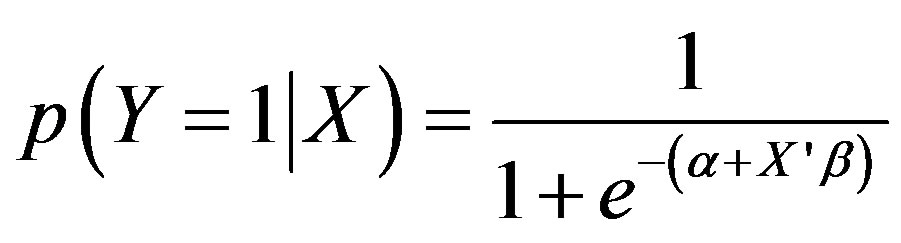 (1)
(1)
where 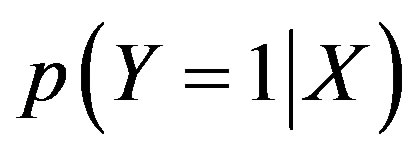 is the expected conditional probability for a remeasured plot to be infested (Y = 1; otherwise, Y = 0 if not infested) by tallow tree between the two consecutive cycles; α is the intercept of the model; X is the vector of associated factors (see Table 1); and β is the vector of estimated coefficients. The stepwise method with the significance level of 0.05 was chosen to select the significant factors associated with the infestation of tallow tree in a remeasured FIA plot. All statistical computation and analyses were conducted under the R statis tical environment by using the packages rpart and stats [25].
is the expected conditional probability for a remeasured plot to be infested (Y = 1; otherwise, Y = 0 if not infested) by tallow tree between the two consecutive cycles; α is the intercept of the model; X is the vector of associated factors (see Table 1); and β is the vector of estimated coefficients. The stepwise method with the significance level of 0.05 was chosen to select the significant factors associated with the infestation of tallow tree in a remeasured FIA plot. All statistical computation and analyses were conducted under the R statis tical environment by using the packages rpart and stats [25].
3. RESULTS
Out of 2426 FIA plots inventoried between 2001 and 2008, 454 plots were found to be infested by Chinese tallow with an overall presence probability of 19% in forestlands of east Texas. The presence probability of tallow tree by forest type group was 33 % in oak/gum/ cypress forests, 23% in longleaf/slash pine forests and elm/ash/cottonwood forests each, 20% in oak/pine forests, 18% in loblolly/shortleaf pine forests, and 8% in oak/hickory forests. All infested plots was dominated by the low (1% ~ 10%) cover class for all forest type group except for oak/gum/cypress where the medium (11% ~ 50%) cover class was dominant (Figure 2(a)).
Excluding formerly infested plots, there were 137 new plots out of the 1900 remeasured FIA plots were infested between 2005 and 2008 with an overall annual infestation probability of 1.8%. However, the annual infestation probability varied by forest type group and took the order of oak/gum/cypress (3.5%) > elm/ash/cottonwood (2.3%) > loblolly/shortleaf pine (1.8%) > oak/pine (1.3%) > oak/hickory and longleaf/slash pine (1%). The newly infested plots were dominated by the trace (<1%) cover class (Figure 2(b)).
Infested plots were primarily clustered in southern
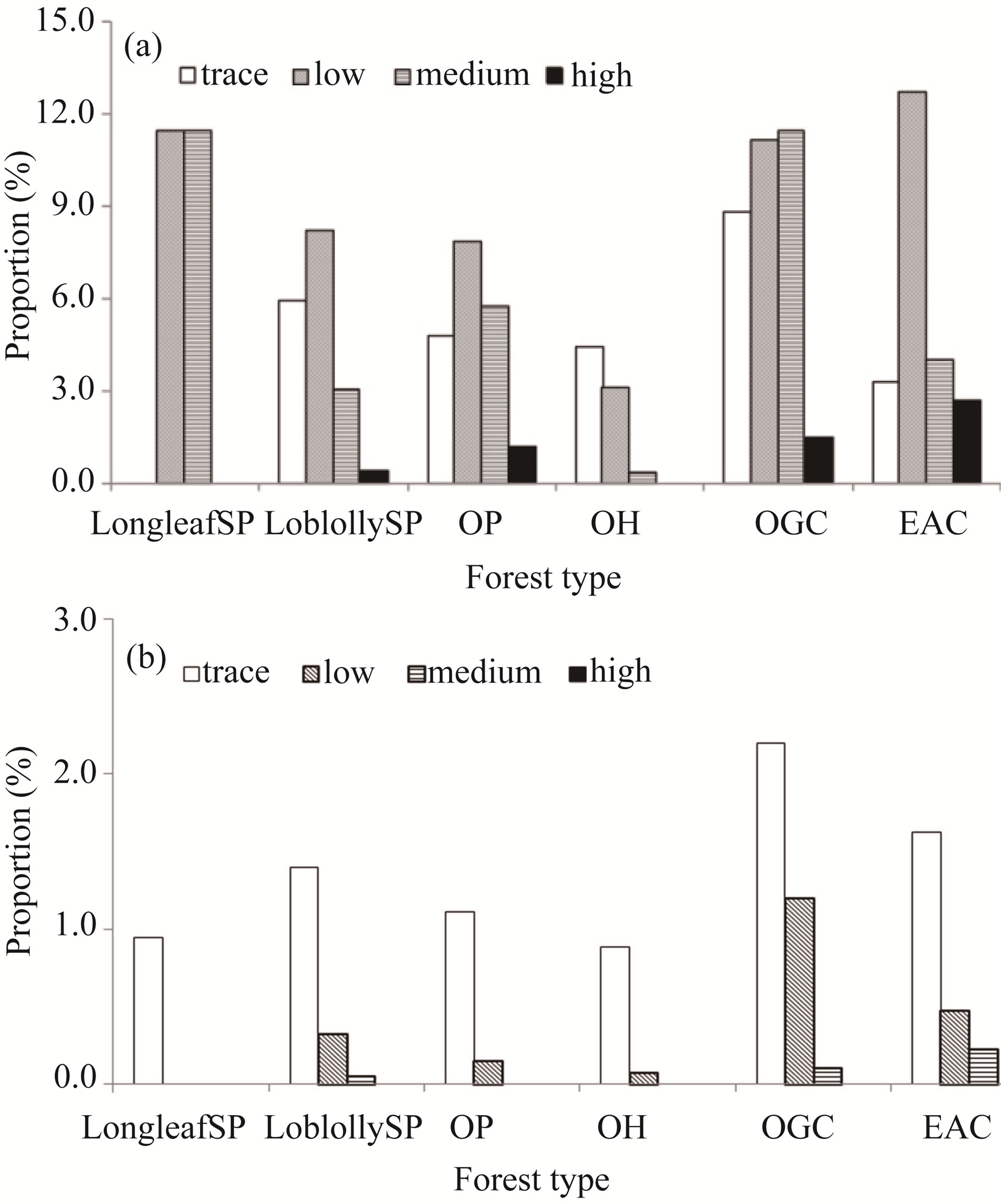
Figure 2. The proportion of plots infested by tallow tree by 2008 (a) and during 2004-2008 (b).
portions of the study region, scattered sporadically in the central portion and there were very few or no infested plots in the northern portion (Figure 3). The smoothed probability map of presence and spread/infestation showed a similar pattern with hot spots (most prevalent areas > 0.5 in probability of presence) mainly found in southern portions of the study region. Spread of Chinese tallow northward is much slower than along the coast where the annual infestation probability is generally >0.05 compared to 0.01 - 0.03 in the central and <0.01 in the north (Figure 4).
The CART model (Figure 5) showed that the annual infestation probability in the coastal area (node S1, latitude ≤ 30.4˚) averaged 9 percent, 7 times higher than that in the inland area (node S4, latitude > 30.4˚). Evidently, there were two hotspots of spread—the east (node S2, centering at the southeast border of Texas and Louisiana) and the west (node S3, the Houston metropolitan area) separated by the meridian of longitude of 94.9˚; the annual infestation probability reached as high as 13.2% in the eastern coastal area, around three times higher than
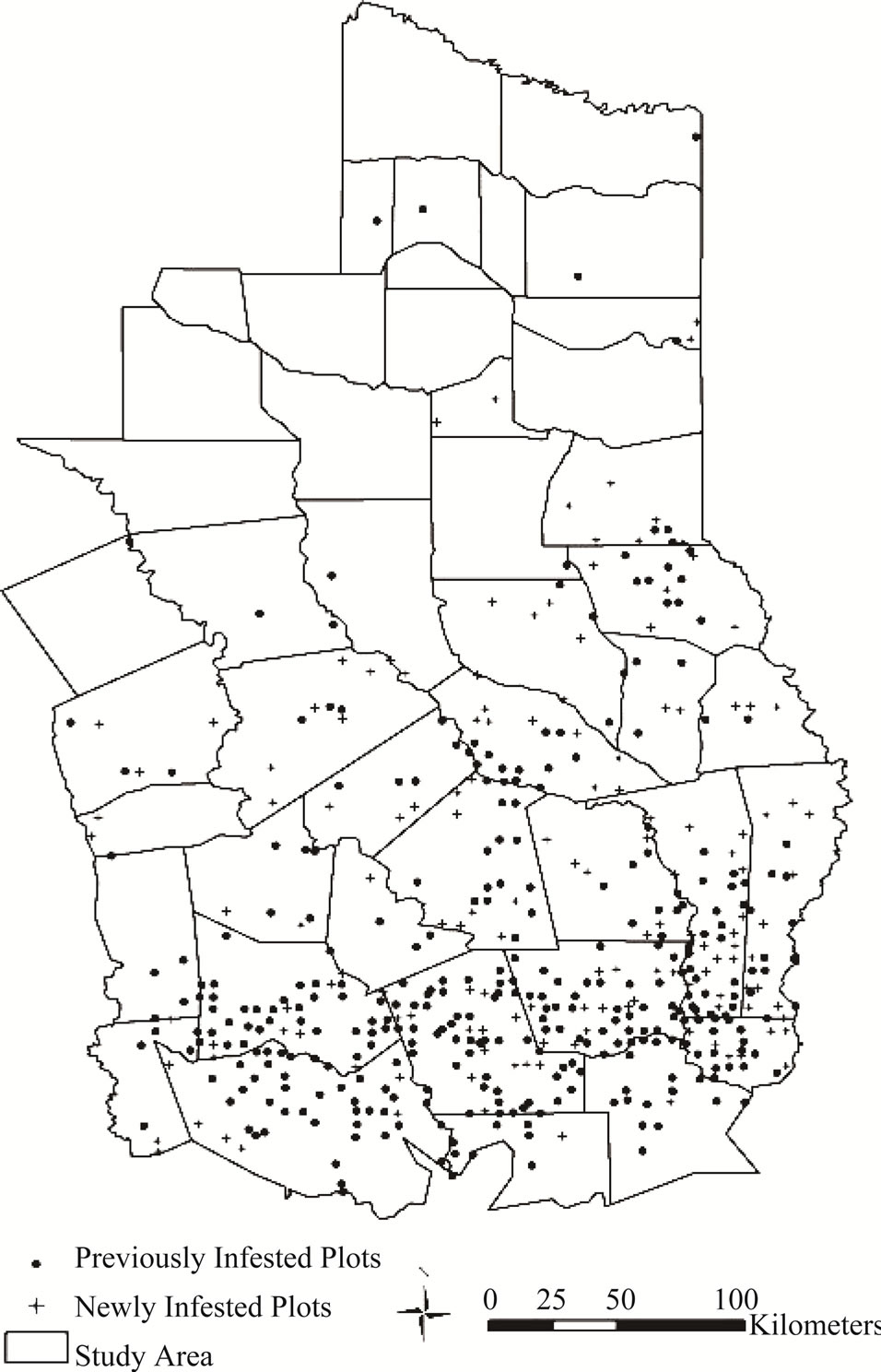
Figure 3. Map of plots previously and newly infested by Chinese Tallow.
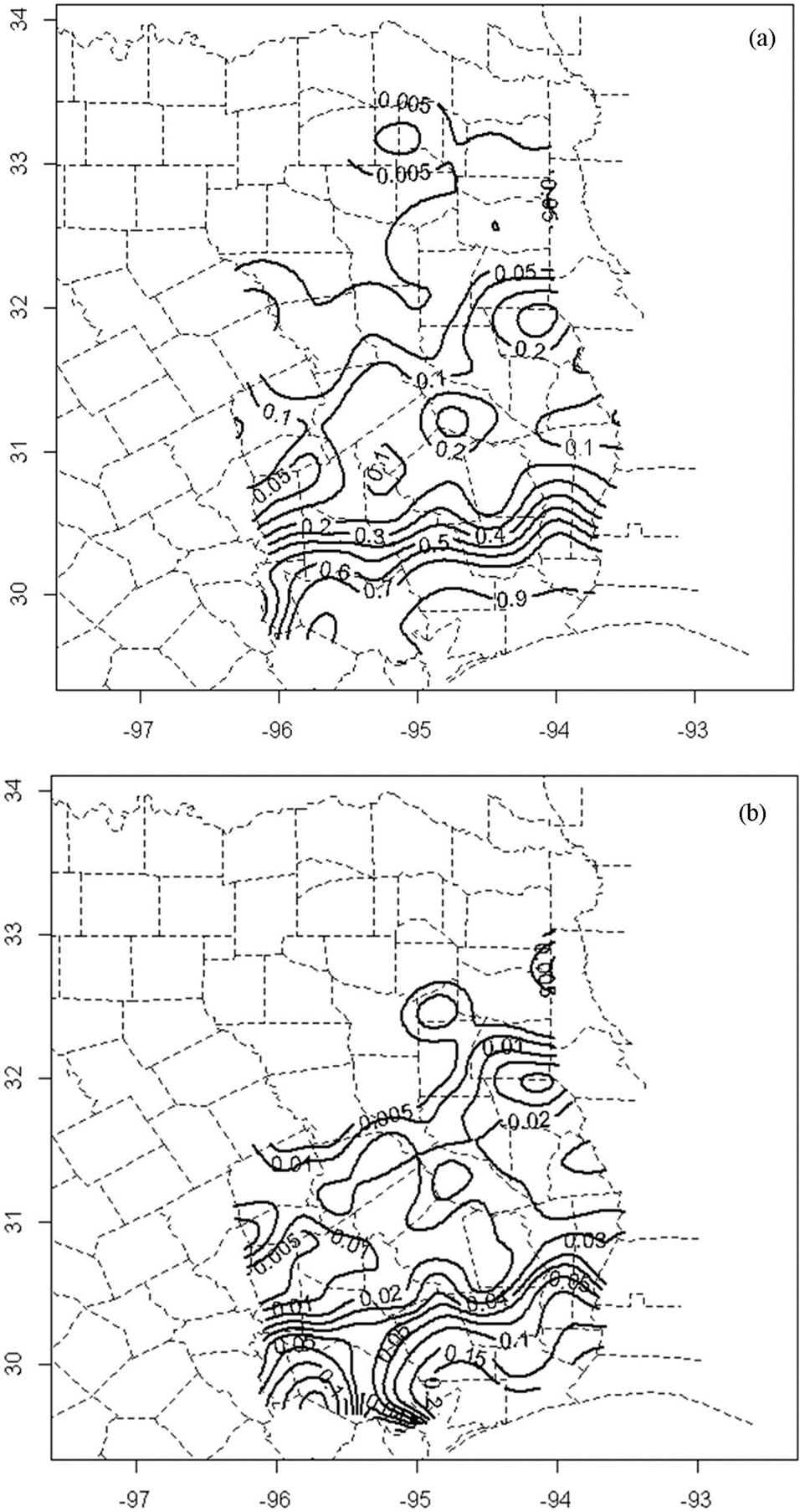
Figure 4. Smoothed probability maps depicting (a) the current condition and (b) the infestation rate of tallow tree in southeast Texas.
that (4.8%) in the western coastal area. Across inland areas, distance to formerly infested plots has proved to be the most significant factor to the spread of tallow, and sites which are within the distance of ≤27.8 km from infested plots (node S5) are highly susceptible to infestation (relative risk = 17.8) compared with sites >27.8 km away from infested plots (node S10). In these highly susceptible sites (node S5), slope appears to be the second most important factor to the spread of tallow; the relative risk for flat areas (node S6, slope ≤ 2%) to be infested by tallow is 3 times larger than in gently hilly areas (node S9, slope > 2%). Similar to the coastal area, a significant difference in annual infestation probability exists between the eastern and western inland, flat areas
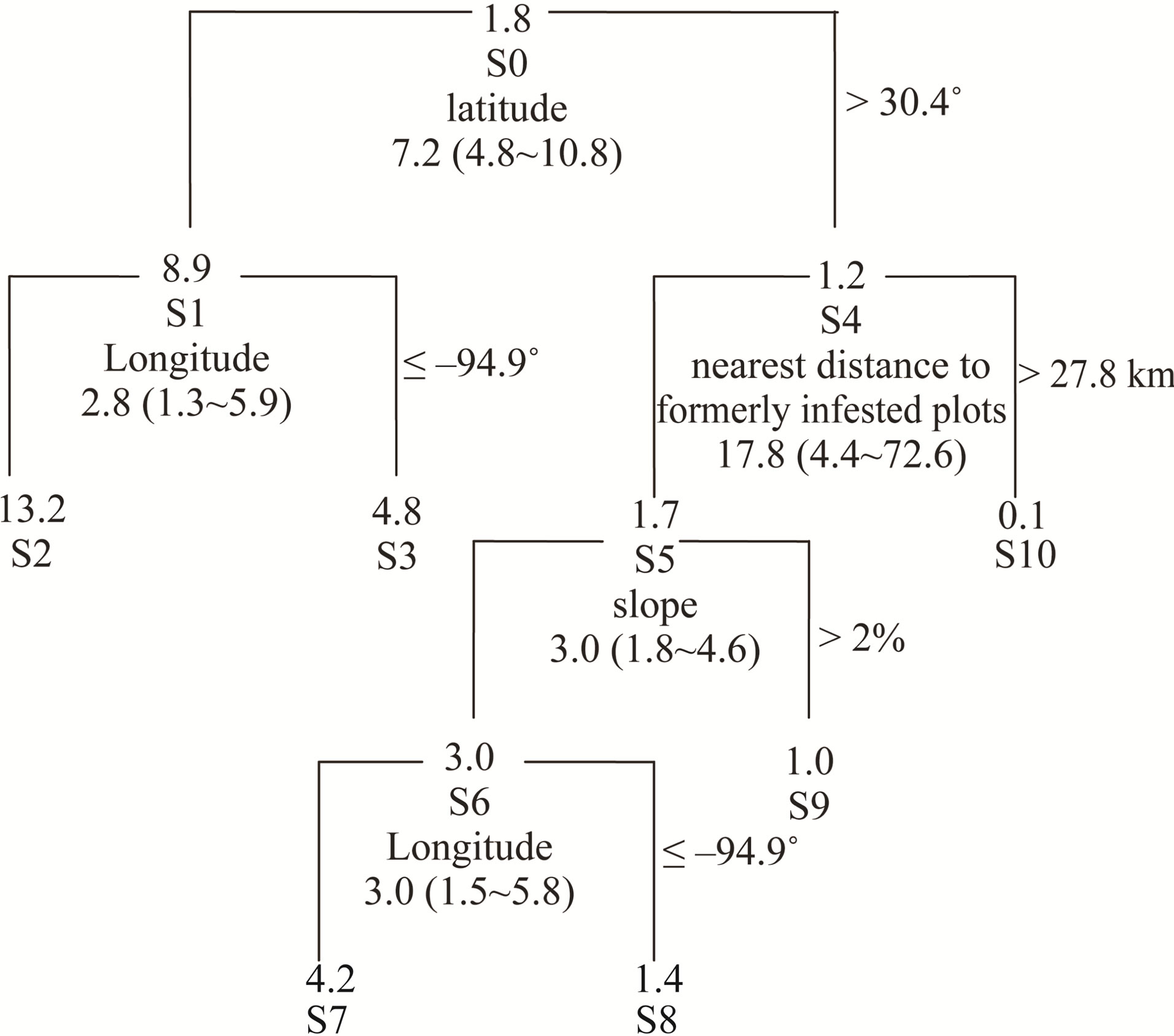
Figure 5. Classification tree model of annual infestation probability (%) by splitting factors based on 1900 repeatedly measured FIA plots between 2004 and 2008 in east Texas. S0 through S10 denote nodes. Beneath node number are the splitting factor, corresponding relative risk and bootstrapped 95% confidence interval (in parentheses).
(nodes S7 and S8) with the former being three times more susceptible to infestation than the latter (Figure 5).
The overall regional logistic regression model showed that distance to formerly infested plots and roads, slope, forest type group, and natural disturbances were significant factors for the spread of tallow, but it did not fit the data well based on the Hosmer and Lemeshow Chisquare goodness-of-fit test (Table 3). The sub-region (node) based logistic regression models indicated that contributing factors might change spatially. For certain sub-regions (represented by nodes S2, S3, S6, S8 and S10) identified by the CART model, none of the factors collected and incorporated in the model initially prove to be significant at the 0.05 significance level; but for those sub-regions with significant factors detected, all models fit the data well based on the Hosmer and Lemeshow Chi-square goodness-of-fit test. For instance, in the coastal area (node S1), only distance to formerly infested plots and roads appears to be significant factors, but for the inland area (node S4), more variables including stand stocking, human activity and forest type group in addition to distance to formerly infested plots are significantly correlated to the infestation of new FIA plots. Depending on the distance to formerly infested plots and slope, factors such as physiographic region and fragmentation status of a site may also affect the infestation probability locally (Table 3).
4. DISCUSSION
Since its initial introduction in Texas, Chinese tallow has continued to thrive, spreading into the marsh, grassland and forestland of southeastern Texas [26]. Effective control or mitigation of Chinese tallow depends on precise information regarding its distribution and spreading mechanisms. Gan et al. [14] and Wang et al. [19] used FIA data and simulation models to evaluate the factors associated with the occurrence of Chinese tallow in east Texas and Louisiana and used the occurrence-environment relationship to predict the potential expansion range. One of the limitations of one-time measurement (e.g., occurrence data) in studying spread and range expansion of invasive plants is that it cannot estimate the spread rate directly and, most often than not, will confound separate risk factors with land use history (e.g., invasion history) which uniquely determines the extent and condition of invasive plants [23,27]. In this study, the FIA plots measured in two consecutive inventory cycles were used to evaluate associated factors and estimate the spread rate of Chinese tallow in eastern Texas. Noticing the strong clustering patterns of tallow invasion, distance to formerly infested plots was calculated and incorporated with other factors in CART and logistical regression in a nested modeling framework to understand the spreading vectors and mechanisms of Chinese tallow.
Both the smoothed probability map and CART model show that the spread of tallow tree into inland areas has been greatly limited. The probability of being infested in the coastal area (node S1) is 7.2 (95% CI: 4.8 - 10.8) times larger than that in the inland area (node S4) (Figure 5). Studies suggest that low temperature may be a major environmental constraint on the spread of Chinese tallow northward [10,14,19]. Tallow tree is forecasted to increase its range over the next 100 years and could speed its invasion rate if average temperatures continue to increase [19]. Seed germination rate and seedling growth are consistently lower on northern and inland sites [28]. The strong clustering patterns and significantly positive association of formerly and newly infested plots (Figure 3) support the spread of Chinese tallow dispersal being limited at large (region or landscape) scales [28]. Other than low temperature which limited seed germination and seedling growth, current distribution of tallow tees (reflected by nearest distance to formerly infested plots) and slope also affect the spread of Chinese tallow through curbing seed dispersal (Figure 5). Change of the infestation probability with distance from formerly infested plots in the inland areas (node S4) indicated that Chinese tallow is inherently a local (short-distance) dispersal invader with an effective dispersal distance of 27.8 km. The average relative risk (ratio of the infestation probabilities) between sites ≤27.8 km and >27.8 km from infested plots reaches as high as 17.8 even though the infestation probability may change with other factors such as slope and longitude. Flat terrain (e.g., slope ≤ 2%) and high propagule pressure (clusters of infested plots) dramatically increase the likelihood of a site being infested
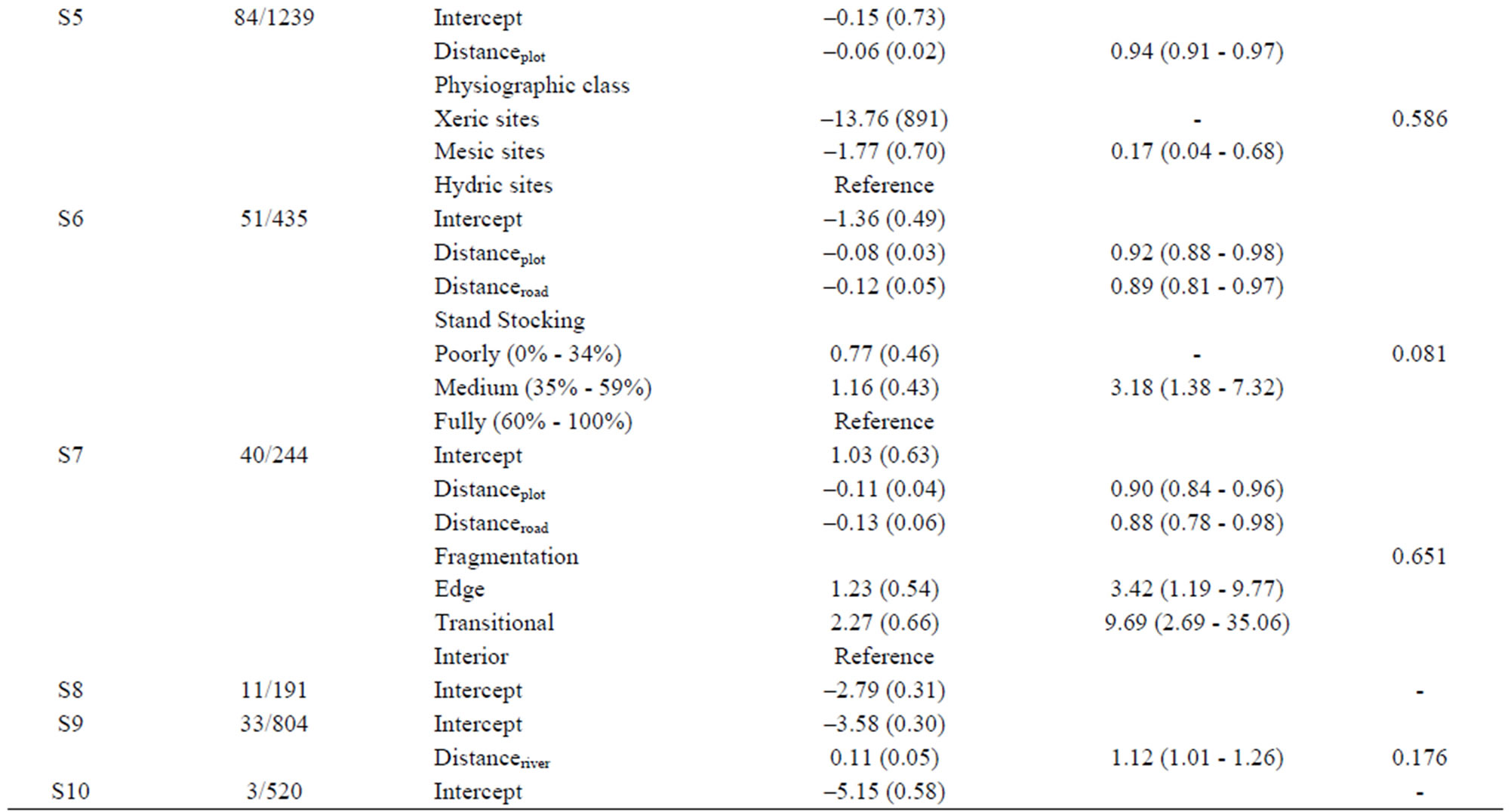
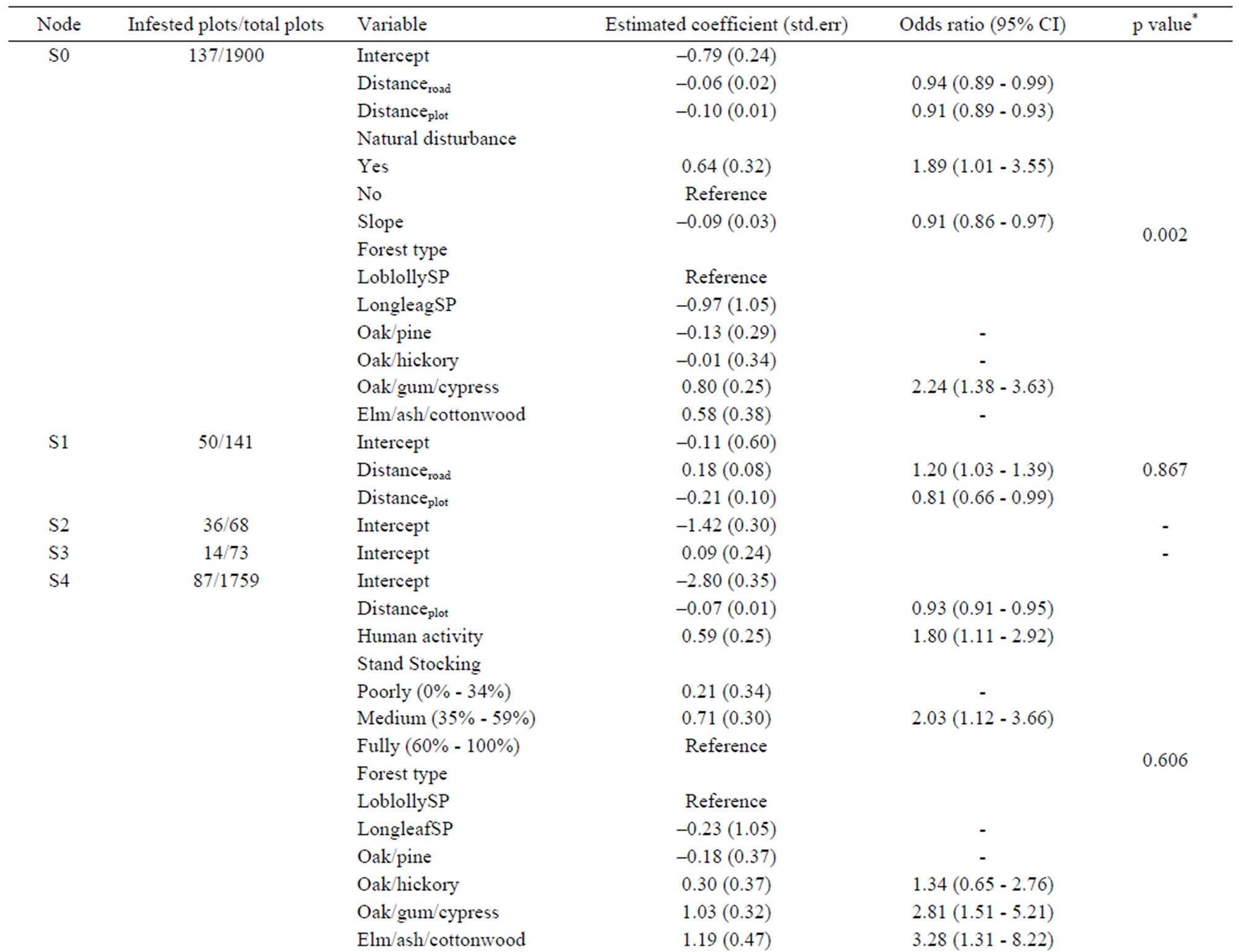
Table 3. Estimated parameters (standard errors) for the overall (node S0) and nested (nodes S1-S10) logistic regression models.
(Figure 4). Tallow seeds can be easily dispersed by birds or carried by rivers, streams, or storm water runoff to new destinations and encounter fewer barriers. Following its colonization, tallow tree often outcompetes native species such as loblolly pine (Pinus taeda) and water oak (Quercus nigra) by its rapid height growth which enables it to effectively control the environment following disturbances and results in a strong clustering pattern [1]. The localized invasion pattern may suggest water (current or storm runoff) as the major vector of seed dispersal even though tallow seeds are readily dispersed by birds [7,26]. This explains why tallow trees are more prevalent in the coastal prairies, flatwoods and bottomlands [3, 14,19].
We found that the invasion patterns of Chinese tallow and associated contributing factors varied across the region (Figures 4 and 5, Table 3). The CART model and a set of nested logistic regression models provide a flexible way to evaluate the effect of contributing factors on the spread rate of Chinese tallow over a large, heterogeneous region such as eastern Texas. Often, a single model may fail to adequately fit the data and result in bias estimation of factors of interest and subsequently inefficient prescriptions to control or mitigate biological invasion. For instance, wind disturbance, a significant determinant of tallow spread in the overall logistic model (node S0), was not shown in the separate, nested logistic models for the coastal (node S1) and inland areas (node S4). Instead, human activity was shown to be an important factor across inland areas (node S4). The chi-square test for partial association showed that natural disturbance was marginally significant (p = 0.08) in the coastal area. While human activity was significant in inland areas at the significance level of p = 0.05, it did not outweigh natural disturbance over the entire region due to, statistically, the sparse infested plots across inland areas. With regard to disturbances (natural and/or anthropogenic), a major driver or determinant of the dispersal of tallow seeds, one explanation of the logistic models in nodes S0, S1 and S4 is that seed dispersal in the coastal area (S1) was mainly related to natural disturbances, while human activities were a major dispersal agent within inland areas (S4). With the great prevalence of infested plots and high intensity and frequency of natural disturbances in the coastal area, eradication or control of tallow trees will be extremely difficult and costly. Grace [29] reported that prescribed burning could be used for limiting its invasion into coastal prairie ecosystems. In contrast, the nested models for the inland areas (node S4 through S10) suggest that the spread of tallow trees further inland might be impeded at current stage with less cost due to the relative sparseness of tallow trees. Regulating human activities and stand management practices which result in stand stocking reductions and forest fragmentation will be critical to curb the spread of tallow trees (Table 3).
Gan et al. [14] classified factors affecting the occupation/occurrence of Chinese tallow into four categories: landscape features (e.g., slope, elevation, adjacency to water bodies), site and forest conditions (e.g., stand age, site productivity, species diversity), natural and anthropogenic disturbances (e.g., timber management, distance to roads, wind, fire, and animal damage), and climatic conditions (e.g., mean minimum temperature in January). The potential effect of these identified factors on the occurrence of tallow tree was discussed in the logistic regression framework by using discriminated county level data which are generated from 2001-2005 FIA data. Basically, landscape features, spatial distribution of tallow trees and disturbances predominately affect the dispersal of seeds, while site and forest condition affect seed germination and seedling growth [28]. Considering the nature (binary presence vs. absence) of tallow tree data, factors identified by the CART and nested logistic regression models are mainly related to the dispersal of tallow trees. Among the six forest type groups, oak/gum/ cypress and elm/ash/cottonwood have the highest rate of spread and prevalence of tallow trees. These two forest type groups dominate in bottomlands and lowlands, respectively, best suited to seed dispersal, while other forest type groups are mainly found in uplands or in dry, coastal plain. Tallow tree was less likely to invade sloping lands than flat lands. Flat lands could better facilitate human and animal movement and enhance seed dispersal and alter the natural ecosystem. The oak/gum/cypress forest type group or hydric sites have been significantly impacted in the southeastern United States [30] and they could serve as dispersal vectors for tallow to invade into other areas. The higher proportions of the cover classes, with the exception of the “trace” class in oak/gum/cypress and elm/ash/cottonwood forest types, indicated that both are also suited to seed germination and growth of tallow trees [31], and have higher invasiveness than other forest types (Figure 2(b)).
To prevent tallow from spreading further inland, intensive survey or sampling of local population size and distribution of infested plots other than FIA is necessary since the infestation probability increases rapidly with the distance from infested plots (odds ratio = 0.93) (node S4 in Table 3). Considering cost restrictions, intensive invasive species surveys may be integrated into timber or other resources surveys. For instance, the Mississippi Institute for Forest Inventory (MIFI) started to implement a high density survey to aid in the assessment of Mississippi’s statewide forest resources [32]. The MIFI plots, which record all trees >2.5 cm in dbh, could be utilized to analyze tallow tree invasion from finer scales (e.g., county) and with higher accuracy (Dr. Tom Matney, Mississippi State University Department of Forestry, personal communication). In addition, a survey targeting susceptible or at-risk areas such as flator low-lands (e.g., slope ≤ 2%) (node S6 shown in the CART model ) or hydric sites along streams and rivers (node S5, as reflected by the logistic model) or sites in close proximity to roads (node S6) should be prioritized where resources (labor or time) are limited. Tallow can bear seeds as early as 3 years old, so proactive treatment is imperative to take full advantage of the short window from initial invasion to subsequent seed dispersal of infesting trees. Mechanically felling or removing trees or killing trees by chemicals or using prescribed burning are available in the southeast [26].
Future research should focus on more extensive sampling for Chinese tallow within various forest types. With a higher resolution dataset it may be possible to more accurately determine spread rates within various ecosystems. Also of importance is a more detailed study of disturbance events (e.g., land-use change, wind damage, fire, etc.) which could accelerate the spread of tallow tree in naturally regenerated ecosystems. It would also be useful to expand the study area across the southeastern United States and to point further inland with subsequent FIA cycles. This level of analysis could lead to more detailed information on the spread of Chinese tallow and allow for the development of more efficient methods for its mitigation.
5. CONCLUSIONS
We have identified the most significant factors regarding the probability of spread of Chinese tallow in southeastern Texas by utilizing a combination of causal factors and remeasured FIA plot data. Close proximity to previously infested plots and roads, flat areas (i.e., low slope), and natural/anthropogenic disturbance events (e.g., hurricanes, harvest) were all linked to the spread of Chinese tallow throughout the region. The highest presence probability and spread rate occurred in the oak/ gum/cypress forest type, characterized by moist, lowland areas. These areas are also in close proximity to coastal areas and are, therefore, more susceptible to damage from land-falling hurricanes; the landfall of Hurricane Rita in this area in 2005 could have exacerbated the spread of Chinese tallow throughout the coastal area.
Forest managers in areas with the presence of tallow tree should take actions to mitigate its spread throughout the ecosystem. Increasing anthropogenic impacts, via urban sprawl and road construction, and more frequent natural disturbances could further fragment forested areas and provide additional means of seed transport. Also, increasing temperatures throughout the region could allow for the spread of tallow tree further north of its current range. Prompt clearing of debris following a harvest or salvage of timber damaged from severe weather events, followed by manually replanting areas of forest loss could help minimize the continued spread of tallow tree in the region.
Future surveys should be conducted more intensively sample (spatially and temporally) for Chinese tallow. This would allow for a more thorough assessment of its presence and spread and allow for the development of more robust models of future changes within various forest types. This would allow for more explicit direction for forest managers/landowners in the region regarding understanding the spread of tallow tree and treatments to mitigate its spread.
REFERENCES
- Bruce, K.A. (1993) Factors affecting the biological invasion of the exotic Chinese tallow tree, Sapium sebiferum, in the Gulf Coast Prairie of Texas. Master Thesis, University of Houston, Houston.
- Miller, J.H. (2003) Nonnative invasive plants of Southern Forests: A field guide for identification and control. USDA Forest Service, Asheville.
- Bruce, K.A., Cameron, G.N. and Harcombe, P.A. (1995) Initiation of a new woodland type on the Texas coastal prairie by the Chinese tallow tree (Sapium sebiferum (L.) Roxb.) Bulletin of the Torrey Botanical Club, 122, 215- 225. doi:10.2307/2996086
- Oswalt, S.N. (2010) Chinese tallow (Triadica sebifer (L.) Small) population expansion in Louisiana, east Texas, and Mississippi. USDA Forest Service, Asheville.
- Bogler, D.J. (2000) Element Stewardship Abstract for Sapium sebiferum. The Nature Conservancy, Arlington.
- Jubinsky, G. and Anderson, L.C. (1996) The invasive potential of Chinese tallow-tree (Sapium sebiferum Roxb.) in the southeast. Castanea, 61, 226-231.
- Renne, I.J., Barrow, W.C., Johnson Randall, L.A. and Bridges, W.C. (2002) Generalized avian dispersal syndrome contributes to Chinese tallow tree (Sapium sebiferum, Euphorbiaceae) invasiveness. Diversity and Distributions, 8, 285-295. doi:10.1046/j.1472-4642.2002.00150.x
- Siemann, E. and Rogers, W.E. (2003) Changes in light and nitrogen availability under pioneer trees may indirectly facilitate tree invasions of grasslands. Journal of Ecology, 91, 923-931. doi:10.1046/j.1365-2745.2003.00822.x
- Zhang, K. and Lin, Y. (1994) Chinese tallow. China Forestry Press, Beijing.
- Pattison, R.R. and Mack, R.N. (2008) Potential distribution of the invasive tree Triadica sebifera (Euphorbiaceae) in the United States: evaluation CLIMEX predictions with field trials. Global Change Biology, 14, 813-826. doi:10.1111/j.1365-2486.2007.01528.x
- Chesson, P. (2000) Mechanisms of maintenance of species diversity. Annual Review of Ecology and Systematics, 31, 343-366. doi:10.1146/annurev.ecolsys.31.1.343
- Sakai, A.K., Allendorf, F.W., Holt, J.S., Lodge, D.M., Molofsky, J., With, K.A., Baughman, S., Cabin, R.J., Cohen, J.E., Ellstrand, N.C., McCauley, D.E., O’Neil, P., Parker, I.M., Thompson, J.N. and Weller, S.G. (2001) The population biology of invasive species. Annual Review of Ecology and Systematics, 32, 305-332. doi:10.1146/annurev.ecolsys.32.081501.114037
- Levine, J.M., Adler, P.B. and Yelenik, S.G. (2004) A meta-analysis of biotic resistance to exotic plant invasions. Ecology Letters, 7, 975-989. doi:10.1111/j.1461-0248.2004.00657.x
- Gan, J., Miller, J.H., Wang, H. and Taylor Jr., J.W. (2009) Invasion of tallow tree into southern US forests: influencing factors and implications for mitigation. Canadian Journal of Forest Research, 39, 1346-1356. doi:10.1139/X09-058
- Theoharides, K.A. and Dukes, J.S. (2007) Plant invasion across space and time: factors affecting nonindigenous species success during four stages of invasion. New Phytology, 176, 256-273. doi:10.1111/j.1469-8137.2007.02207.x
- Chapman, E.L., Chambers, J.Q., Ribbeck, K.F., Baker, D.B., Tobler, M.A., Zeng, H. and White, D.A. (2008) Hurricane Katrina impacts on forest trees of Louisiana’s Pearl River basin. Forest Ecology and Management, 256, 883-889. doi:10.1016/j.foreco.2008.05.057
- Pysek, P., Richardson, D.M., Rejmánek, M., Webster, G.L., Williamson, M. and Kirschner, J. (2004) Alien plants in checklists and floras: towards better communication between taxonomists and ecologists. Taxon, 53, 131-143. doi:10.2307/4135498
- Battaglia, L.L, Denslow, J.S., Inczauskis, J.R. and Baer, S.G. (2009) Effects of native vegetation on invasion success of Chinese tallow in a floating marsh ecosystem. Journal of Ecology, 97, 239-246. doi:10.1111/j.1365-2745.2008.01471.x
- Wang, H.H., Grant, W.E., Swannack, T.M., Gan, J., Rogers, W.E., Koralewski, T.E., Miller, J.H. and Taylor Jr., J.W. (2011) Predicted range expansion of Chinese tallow tree (Triadica sebifera) in forestlands of the southern United States. Diversity and Distributions, 17, 552-565. doi:10.1111/j.1472-4642.2011.00760.x
- Conner, R.C. and Hartsell, A.J. (2002) Forest area and conditions. USDA Forest Service, Asheville.
- Bechtold, W.A. and Patterson, P.L. (2005) The enhanced forest inventory and analysis program-national sampling design and estimation procedures. USDA Forest Service, Asheville.
- Moser, W.K., Hansen, M.H., Nelson, M.D. and Mc-Williams, W.H. (2009) Relationship of invasive groundcover plant presence to evidence of disturbance in the forests of the Upper Midwest of the United States. Chapter 3. In: Kohli, R.K., Jose, S., Singh, H.P. and Batish, D.R., eds., Invasive plants and forest ecosystems, CRC Press, Boca Raton, 29-58.
- Fan, Z., Moser, W.K., Hansen, M.H. and Nelson, M.D. (2012) Regional patterns of major non-native invasive plants and associated factors in Upper Midwest forests. Forest Science, 58.
- Fan, Z., Kabrick, J.M. and Shifley, S.R. (2006) Classification and regression tree based survival analysis in oakdominated forests of Missouri’s Ozark highlands. Canadian Journal of Forest Research, 36, 1740-1748. doi:10.1139/x06-068
- R Development Core Team (2011) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. http://www.R-project.org/
- Bruce, K.A., Cameron, G.N., Harcombe, P.A. and Jubinsky, G. (1997) Introduction, impact on native habitats, and the management of a woody invader, the Chinese Tallow Tree, Sapium sebiferum (L.) Roxb. Natural Areas Journal, 17, 255-260.
- Rejmánek, M. (1989) Invasibility of plant communities. In: Drake, J.A., Mooney, H.A., di Castri, F., Groves, R.H., Kruger, F.J., Rejmánek, M. and Williamson, M., eds., Biological Invasions. A Global Perspective, John Wiley & Sons, Chichester, 369-388.
- Pattison, R.R. and Mack, R.N. (2009) Environmental constraints on the invasion of Triadica sebifera in the eastern United States: an experimental field assessment. Oecologia, 158, 591-602. doi:10.1007/s00442-008-1187-7
- Grace, J.B. (1998) Can prescribed fire save the endangered coastal prairie ecosystem from Chinese tallow? Endangered Species Update, 15, 70-76.
- Tan, Y., Fan, Z. and Oswalt, C.W. (2010) Impact of ecological and socioeconomic determinants on the spread of tallow tree in southern forest lands. In: Merry, K., Bettinger, P., Fan, Z., Kushla, J., Litts, T., Siry, J., Hepinstall-Cymerman, J. and Song, B., eds., Proceedings of the 7th Southern Forestry and Natural Resources GIS Conference, Athens, 115-123.
- Conner, W.H. (1994) The effect of salinity and waterlogging on the growth and survival of baldcypress and Chinese tallow seedlings. Journal of Coastal Research, 10, 1045-1049.
- Parker, R.C., Glass, P.A., Londo, H.A., Evans, D.L., Belli, K.L., Matney, T.G. and Schultz, E.B. (2005) Mississippi’s forest inventory pilot program: use of computer and spatial technologies in large area inventories. Mississippi State University, Starkville.