Open Journal of Genetics
Vol.1 No.3(2011), Article ID:8889,10 pages DOI:10.4236/ojgen.2011.13008
Effect of temperature on crossing over in the mus309 mutant, deficient in DNA double-strand break repair, of Drosophila melanogaster suggests a mechanism for crossover interference
Laboratory of Genetics, Department of Biology, University of Turku, Turku, Finland.
Email: petter.portin@utu.fi
Received 16 September 2011; revised 27 October 2011; accepted 19 November 2011.
Keywords: Chiasma; Chromosome; Map Length; Meiosis
ABSTRACT
Crossing-over frequencies, crossover interference, recombination frequencies and map distances were compared in the cv-v-f region of the X chromosome of Drosophila melanogaster in females bearing either wild type 3rd chromosomes (control) or having the DNA double-strand break repair deficient mus309D2/mus309D3 mutant constitution in the 3rd chromosomes (experiment), and raised in three different temperatures viz. 18˚C, 25˚C and 29˚C. In addition, the fecundity of the females was also measured. In the control crosses none of the mean values of the parameters measured was dependent on the temperature, whereas in the experimental crosses all the parameters, except for the frequency of true single crossovers in the cv-v interval, the recombination frequency of the v and f markers, and the coefficient of coincidence, changed due to the effect of temperature. When comparing the genotypes studied, a significant difference between them was observed in all the parameters measured, apart from the frequency of the true single crossovers in the cv-v interval. These results support the counting number model of the mechanism of interference based on the genetic distance, but are in contradiction with the models based on physical distance.
1. INTRODUCTION
1.1. General Introduction
Meiosis is a fundamental process in all sexually reproducing eukaryotes, consisting of two successive divisions of the meiocyte nucleus but only one cycle of replication of the chromosomes. Meiosis involves pairing of the homologous chromosomes and, exchange by crossing over of genetic material between them resulting in the formation of chiasmata. A chiasma is a sufficient condition for the segregation of homologous chromosomes, which leads to the reduction of the chromosome number from diploid to haploid. The alternation of meiosis and fertilization leads to the recombination of genetic material.
An important phenomenon associated with crossing over is crossover interference, i.e. the fact that multiple crossovers are less frequent than would be expected on the basis of random coincidence of single crossovers [1-3]. Crossover interference has recently garnered considerable attention because it may be responsible for the occurrence crossovers called obligate crossovers, and thus for the formation of obligate chiasmata.
The term “obligate crossover” refers to the fact that, in most species, it is rare to find chromosomes that do not undergo crossing over. For example, in Drosophila, there is usually one chiasma per chromosome arm. The feature of the obligate chiasma is biologically sensible because it ensures the disjunction of homologous chromosomes.
1.2. Models of Crossover Interference and the Aim of the Present Study
Models of crossover interference can, in principle, be divided into two different categories. The first category of models, called genetic models [4], assumes that interference is dependent on the genetic (i.e. linkage map) distance (Morgans) between adjacent crossovers. To my knowledge, currently only one model, called the ‘counting model’, falls into this category [4,5].
The second category of models, which may be called physical models, hypothesize that crossover interference is dependent on the physical distance (microns or base pairs) between the adjacent crossovers. In general, these models, which are many, suggest that some kind of physical signal travels along the bivalent and determines the distribution of crossovers.
The aim of the present study was to distinguish between the explanatory values of these models. Crossingover frequencies, crossover interference, recombination frequencies and map distances were compared in the cv-v-f region of the X chromosome of Drosophila melanogaster in females bearing either wild type 3rd chromosomes (control) or having the DNA doublestrand break repair deficient mus309D2/mus309D3 mutant constitution in the 3rd chromosomes (experiment), and raised in three different temperatures viz. 18˚C, 25˚C and 29˚C. In addition, the fecundity of the females was also measured.
The combined effect of temperature and the mus309 mutation on crossing over and crossover interference was chosen as the experimental design in the first place because temperature is easy to control. The second, and more important reason, however, was that following the arguments of several earlier authors [6-8], and as explained in Discussion, by doing so one can tell something about the mechanism of crossing over and crossover interference in particular.
It was observed that in the control crosses, none of the mean values of the parameters measured was dependent on the temperature, whereas in the experimental crosses all the parameters except for the frequency of true single crossovers in the cv-v interval, the recombination frequency of the v and f markers, and the coefficient of coincidence, changed due to the effect of temperature. When comparing the genotypes studied, a significant difference between them was observed in all the parameters studied, except for the frequency of the true single crossovers in the cv-v interval. These results support the counting number model of the mechanism of interference based on the genetic distance, but are in contradiction with the models based on physical distance.
1.3. Molecular Models of Crossing Over and the mus309 Gene
Present molecular models of meiotic crossing over and gene conversion suggest that crossing over is initiated by the formation of DNA double-strand breaks (DSBs), catalyzed most likely in all eukaryotes by the topoisomerase-like Spo11 protein, which in Drosophila is encoded by the mei-W68 gene [9], in co-operation with other enzymes [10-13]. The occurrence of DSBs is followed by formation of heteroduplex DNA and rejoining of the ends created in the breakage involving a singleend-invasion intermediate (SEI). Following this, a physical structure called a displacement loop, or D-loop, is formed. Subsequent DNA synthesis and second end capture form a structure known as a double Holliday junction (dHJ), which is then resolved to form either crossovers or noncrossovers [14,15].
Two alternative pathways for the repair of the DSBs are known: the synthesis-dependent strand annealing (SDSA) pathway and the double-strand-break repair (DSBR) pathway. The former pathway leads exclusively to non-crossover products and the latter to both crossover and non-crossover products [16,17].
In Drosophila melanogaster, the mus309 locus on the right arm of chromosome three (86F4) encodes, in a manner similar to its orthologues in other organisms, the mammalian BLM locus included, a RecQ helicase [18-21] and, accordingly, is involved in DSB repair [16,17,22]. In particular, it is known that the product of the mus309 locus is involved in the SDSA pathway of the repair of the DSBs [23,24]. More specifically, it is also known that in mus309 mutants the SDSA pathway is blocked, while the DSBR pathway remains functional [25]. Thus, the mus309 gene seems to control the choice made by the oocyte between the two alternative pathways of DSB repair. The same is also true for the mus309 orthologue, the Sgs1 locus, in yeast [26]. Consequently, if in mus309 mutants more DSBs are repaired as crossovers by the DSBR pathway, a change in the crossover/non-crossover ratio can be expected, since fewer non-crossovers are produced.
2. MATERIALS AND METHODS
2.1. Experimental Procedures
The effect of temperature on crossing over frequency and crossover interference in the X chromosome in the regions between the crossveinless (cv, 1 - 13.7), vermilion (v, 1 - 33.0) and forked (f, 1 - 56.7) markers was compared in the experimental and control crosses indicated below. In the control fly crosses, cv v f/+ + +; +/+ females derived from the preliminary cross in which homozygous cv v f females were crossed with + + +/Y males, were crossed with their cv v f/Y brothers. In the experimental crosses, cv v f/+ + +; mus309D2/mus309D3 females were crossed with cv v f/Y males. The experimental females were derived from the following preliminary cross: cv v f; mus309D3/TM6, Tb females crossed with + + +/Y; mus309D2/TM6, Tb males (Tb; Tubby 3 - 90.6) and identified on the basis of their non-Tubby phenotype. Both the preliminary and the actual crosses were made at three different temperatures, viz. 18˚C ± 1˚C, 25˚C ± 1˚C and 29˚C ± 0.5˚C.
Both the mus309 alleles used carry mutational changes that could potentially impair or abolish at least the helicase function of the MUS309 protein. In mus309D2, there is a stop codon between the sequence motifs encoding the third and fourth helicase motif of the protein. mus309D3, for its part, has a glutamic acid to lysine substitution in the conserved helicase II motif, in addition to another amino acid substitution close to the C terminus [27]. It has been demonstrated that the genotype mus309D2/ mus309D3 is semi-sterile [27-29].
Because of the semi-sterility of the females, the experimental crosses were carried out in cultures in which 3 females were mated with 3 - 5 males, whereas the control crosses were single-female cultures. The same number (20) of crosses was made in both the control and the experimental series. The progeny were raised on a standard Drosophila medium consisting of semolina, syrup, agar-agar and both dried and fresh yeast. The parental females were allowed to lay eggs in the culture bottles for four days and were transferred thereafter together with the males into fresh culture bottles for two more days of egg-laying. Thus, progeny was the result of a total of six days. Because of accidental death of the females, progeny was not derived from every culture bottle. The numbers of bottles which gave progeny in the control crosses were as follows: In 18˚C 19 bottles, in 25˚C 19 bottles, and in 29˚C 18 bottles. The respective numbers in the experimental crosses were as follows: In 18˚C 19 bottles, in 25˚C 20 bottles, and in 29˚C 20 bottles.
2.2. Calculation of the Frequency of the True Single Crossovers
Some of the observed single crossovers in the cv – v and v – f intervals actually result from meioses that have two exchanges, one in each interval. Assuming no chromatid interference, the three classes of double-exchange tetrads, 2-, 3- and 4-strand doubles, occur in a 1:2:1 ratio [30]. Therefore, the true frequency of single crossovers, i.e. the number of single crossovers that resulted from meioses with only one exchange in the cv-v-f region, was calculated by subtracting the observed frequency of double crossovers from that of each of the single crossover classes.
2.3. Measurement of Interference
The coefficient of coincidence, C, was calculated according to the following formula [31], which is a maximum likelihood equation
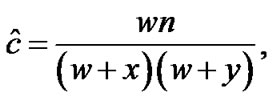 (1)
(1)
where w is the number of flies which were double crossovers, x and y are the numbers of flies which were single crossovers for cv and v, and v and f, respectively, and n is the total number of flies.
The variance of C was calculated according to the following formula [31]
 (2)
(2)
where a and b are the recombination frequencies of cv and v, and v and f, respectively. This is also a maximum likelihood equation.
2.4. Statistical Methods
In the statistical analyses of the results, one-way analysis of variance, Student’s t-test and the binomial t-test were employed. The results of the analyses of variance as well as those of the student’s t-test including their signifycances are always given in the figure legend of the relevant figure. Similarly, the results of the binomial t-tests are given in the appropriate table (Table 3).
3. RESULTS
The distribution of the progeny into different phenotypic classes in each temperature as well as their total numbers in the control crosses is given in (Table 1), and the respecve distribution in the experimental crosses in (Table 2).
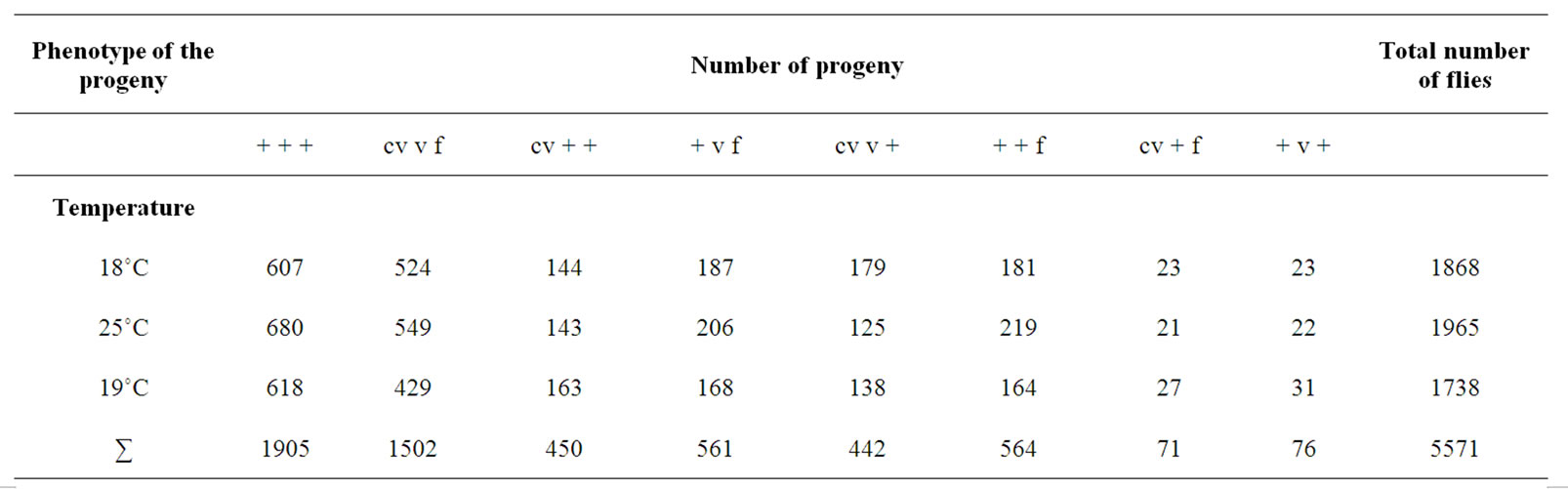
Table 1. The results of the control crosses; distribution of progeny raised at three different temperatures (18˚C, 25˚C and 29˚C) of cv v f/+ + +; +/+ females crossed with cv v f/Y; +/+ males.
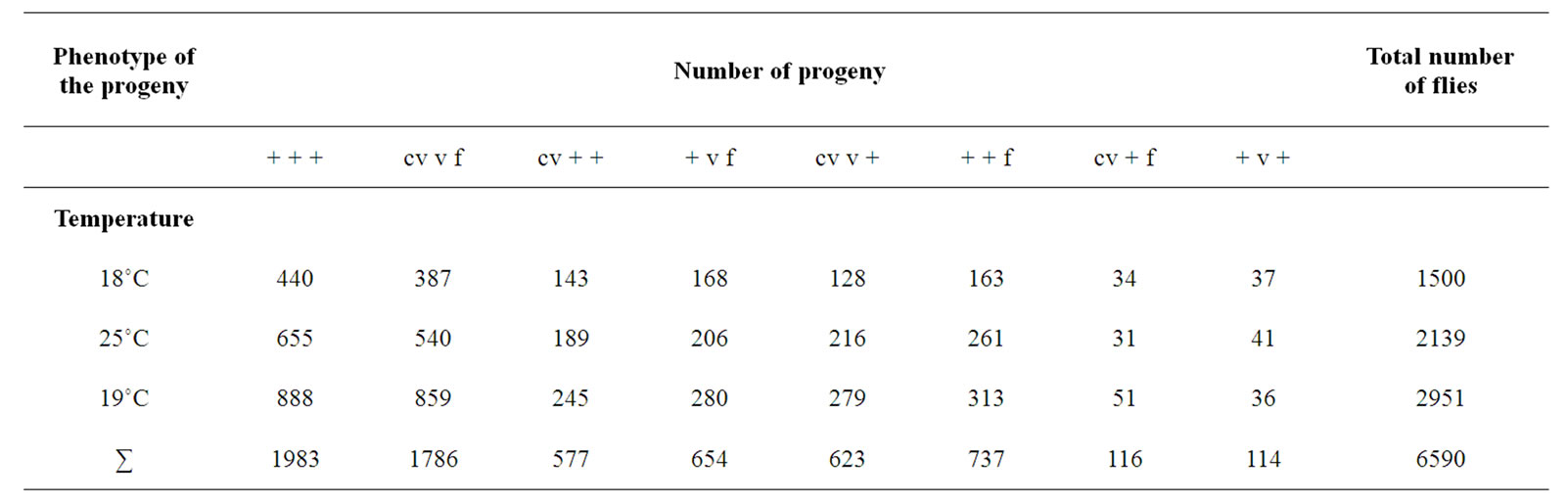
Table 2. The results of the experimental crosses; distribution of progeny raised at three different temperatures (18˚C, 25˚C and 29˚C) of cv v f/+ + +; mus309D2/mus309D3 females crossed with cv v f/Y; +/+ males.
The number of progeny per female, i.e. the fecundities of the females, in each temperature for the control and experimental crosses is given in (Figure 1). As appears from the figure, the fecundity of the control females was not dependent on the temperature, whereas that of the experimental females was. It appears also that the mean value of the fecundity of the experimental females was significantly lower than that of the control females (P < 0.0001).
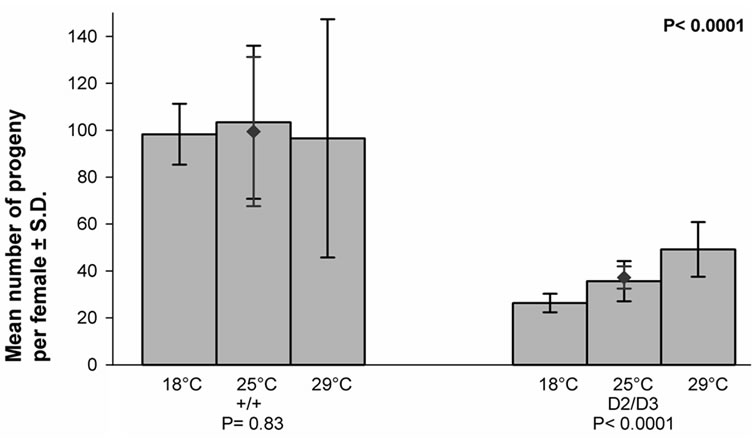
Figure 1. The effect of temperature on the fecundity of the mus309+/mus309+ (+/+) and mus309D2/mus309D3 (D2/D3) females kept in three different temperatures, viz. 18˚C, 25˚C and 29˚C. Columns represent the mean numbers of progenies per female in each temperature in the two genotypes and the vertical lines their standard deviations. The mean number together with its standard deviation for both genotypes is also given and indicated by the red diamond (♦). Testing of the heterogeneity of the results inside each mus309 genotype was carried out using the one-way analysis of variance. The results were as follows: genotype +/+: F = 0.19, df1 = 2, df2 = 53, P = 0.83; genotype D2/D3: F = 34.0, df1 = 2, df2 = 56, P < 0.0001. The significance of the difference of the mean values mentioned was calculated according to the Student’s t-test, the result of which was as follows: t = 14.85, df = 115 and P < 0.0001. In addition, the P-values for the test of heterogeneity are given below the group of columns for the respective genotype, and that for the test of the difference between the genotypes in the upper right corner of the figure.
The frequencies of the true single crossovers in the cv-v interval in each temperature for the control and experimental females are given in (Figure 2). The frequency was temperature sensitive in neither group of females. Nor was there any difference between the control and experimental females (P = 0.9215).
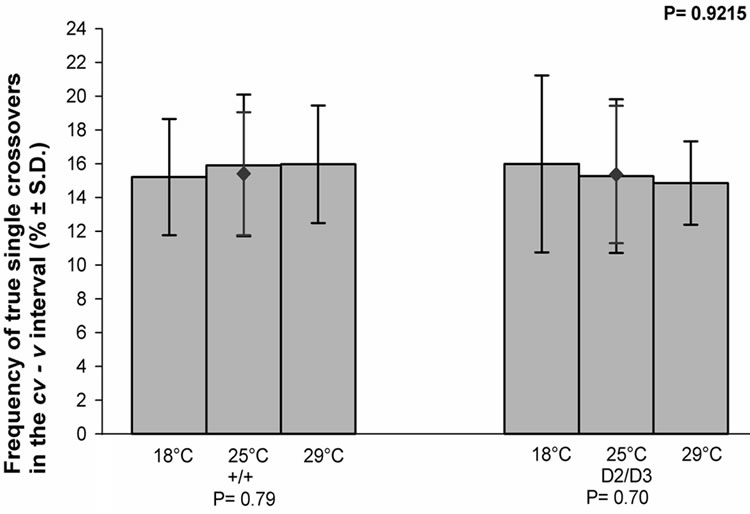
Figure 2. The effect of temperature on the frequency of the true single crossovers between the cv and v markers of the X chromosome in the progeny of the mus309+/mus309+ (+/+) and mus309D2/mus309D3 (D2/D3) females raised at three different temperatures, viz. 18˚C, 25˚C and 29˚C. The columns represent the mean frequencies of crossovers in percentages per culture bottle and the vertical lines their standard deviations. The mean frequency together with its standard deviation for both genotypes is also given and indicated by the red diamond (♦).Testing of the heterogeneity of the results inside each mus309 genotype was carried out using the one-way analysis of variance. The results were as follows: genotype +/+: F = 0.24, df1 = 2, df2 = 53, P = 0.79; genotype D2/D3: F = 0.36, df1 = 2, df2 = 56, P = 0.70. The significance of the difference of the mean values mentioned was calculated according to the Student’s t-test, the result of which was as follows: t = 0.0987, df = 114 and P = 0.9215. In addition, the P-values for the test of heterogeneity are given below the group of columns for the respective genotype, and that for the test of the difference between the genotypes in the upper right corner of the figure.
The frequencies of the true single crossovers in the v -f interval in each temperature for the control and experimental crosses are given in (Figure 3). As shown in the figure, the frequency in the control crosses was not dependent on the temperature, whereas that of the experimental crosses was. It also appears that the mean value of the frequency in question was significantly higher in the experimental than in the control crosses (P = 0.0007).
The frequencies of the double crossovers in the cv-v-f region of the X chromosome in each temperature for the control and experimental crosses are given in (Figure 4). As can be seen in the figure, the frequency in the control crosses was not dependent on the temperature, whereas that of the experimental crosses was. It also appears that the mean value of the frequency in question was significantly higher in the experimental than in the control crosses (P = 0.0039).
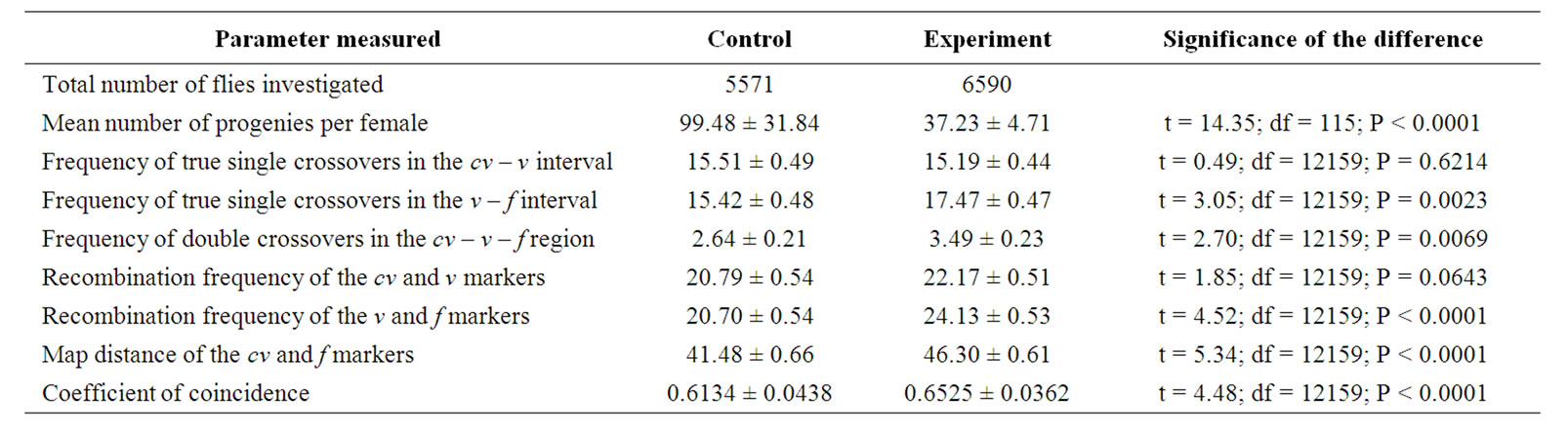
Table 3. Comparison of different parameters calculated on the basis of the total number of progeny of cv v f/+ + +; +/+ (control) and cv v f/+ + +; mus309D2/mus309D3 (experimental) females crossed with cv v f/Y; +/+ males, and raised at three different temperatures viz. 18˚C, 25˚C and 29˚C.

Figure 3. The effect of temperature on the frequency of the true single crossovers between the v and f markers of the X chromosome in the progeny of the mus309+/mus309+ (+/+) and mus309D2/mus309D3 (D2/D3) females raised in three different temperatures, viz. 18˚C, 25˚C and 29˚C. The columns represent the mean frequencies of crossovers in percentages per culture bottle and the vertical lines their standard deviations. The mean frequency together with its standard deviation for both genotypes is also given and indicated by the red diamond (♦).Testing of the heterogeneity of the results inside each mus309 genotype was carried out using the one-way analysis of variance. The results were as follows: genotype +/+: F = 2.04, df1 = 2, df2 = 53, P = 0.14; genotype D2/D3: F = 3.67, df1 = 2, df2 = 56, P = 0.032. The significance of the difference of the mean values mentioned was calculated according to the Student’s t-test, the result of which was as follows: t = 3.49, df = 114 and P = 0.0007. In addition, the P-values for the test of heterogeneity are given below the group of columns for the respective genotype, and that for the test of the difference between the genotypes in the upper right corner of the figure.
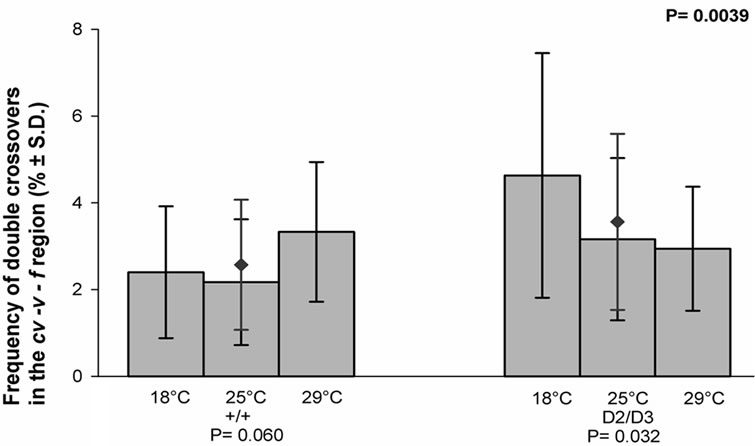
Figure 4. The effect of temperature on the frequency of the double crossovers in the cv-v-f region of the X chromosome in the progeny of the mus309+/mus309+ (+/+) and mus309D2/ mus309D3 (D2/D3) females raised in three different temperatures, viz. 18˚C, 25˚C and 29˚C. The columns represent the mean frequencies of crossovers in percentages per culture bottle and the vertical lines their standard deviations. The mean frequency together with its standard deviation for both genotypes is also given and indicated by the red diamond (♦). Testing of the heterogeneity of the results inside each mus309 genotype was carried out using the one-way analysis of variance. The results were as follows: genotype +/+: F = 2.97, df1 = 2, df2 = 53, P = 0.060; genotype D2/D3: F = 3.67, df1 = 2, df2 =56, P = 0.032. The significance of the difference of the mean values mentioned was calculated according to the Student’s t-test, the result of which was as follows: t = 3.00, df = 115 and P = 0.0039. In addition, the P-values for the test of heterogeneity are given below the group of columns for the respective genotype, and that for the test of the difference between the genotypes in the upper right corner of the figure.
The recombination frequencies of the cv and v markers, i.e. their map distances, in each temperature for the control and experimental crosses are given in (Figure 5). As illustrated in the figure, the frequency in the control crosses was not dependent on the temperature, whereas that of the experimental crosses was. It also appears that the mean value of the frequency in question was signifycantly lower in the experimental than in the control crosses (P = 0.0002).
The recombination frequencies of the v and f markers, i.e. their map distances, in each temperature for the control and experimental crosses are given in (Figure 6). The frequency was temperature sensitive in neither group of crosses. However, the mean value of the map distance in the experimental crosses was significantly higher than in the control crosses (P < 0.0001).
The map distances of the cv and f markers in each temperature for the control and experimental crosses are given in (Figure 7). As appears from the figure, the map distance in the control crosses was not dependent on the temperature, whereas that in the experimental crosses was. It also appears that the mean value of the map distance in question was significantly higher in the experimental than in the control crosses (P < 0.0001).
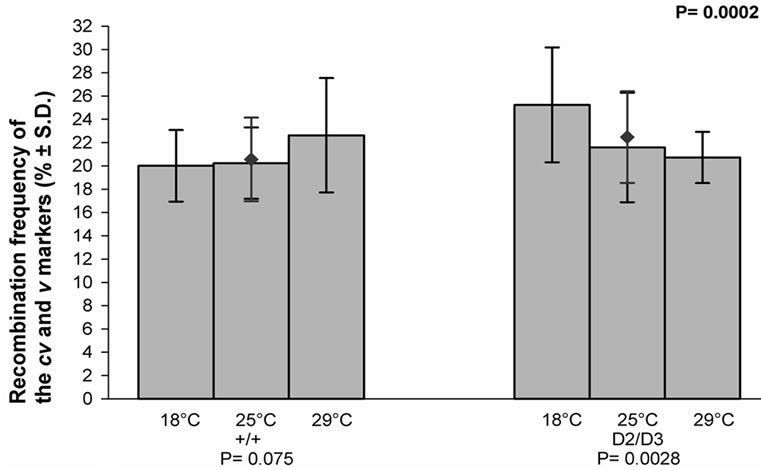
Figure 5. The effect of temperature on the recombination frequency of the cv and v markers of the X chromosome in the progeny of the mus309+/mus309+ (+/+) and mus309D2/mus309D3 (D2/D3) females raised in three different temperatures, viz. 18˚C, 25˚C and 29˚C. The columns represent the mean frequencies of recombinant progenies in percentages per culture bottle and the vertical lines their standard deviations. The mean frequency together with its standard deviation for both genotypes is also given and indicated by the red diamond (♦). Testing of the heterogeneity of the results inside each mus309 genotype was carried using the one-way analysis of variance. The results were as follows: genotype +/+: F = 2.72, df1 = 2, df2 = 53, P = 0.075; genotype D2/D3: F = 6.56, df1 = 2, df2 = 56, P = 0.0028. The significance of the difference of the mean values mentioned was calculated according to the Student’s t-test, the result of which was as follows: t = 3.86, df = 114 and P = 0.0002. In addition, the P-values for the test of heterogeneity are given below the group of columns for the respective genotype, and that for the test of the difference between the genotypes in the upper right corner of the figure.
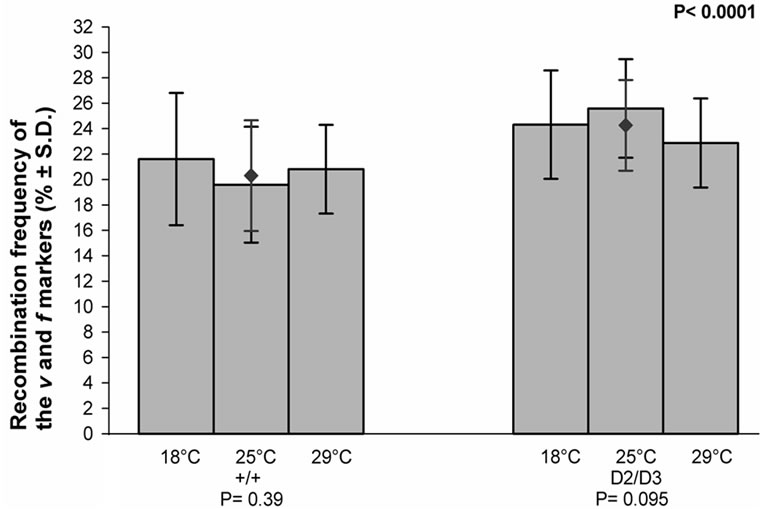
Figure 6. The effect of temperature on the recombination frequency of the v and f markers of the X chromosome in the progeny of the mus309+/mus309+ (+/+) and mus309D2/mus309D3 (D2/D3) females raised in three different temperatures, viz. 18˚C, 25˚C and 29˚C. The columns represent the mean frequencies of recombinant of progenies per culture bottle and the vertical lines their standard deviations. The mean frequency together with its standard deviation for both genotypes is also given and indicated by the red diamond (♦). Testing of the heterogeneity of the results inside each mus309 genotype was carried out using the one-way analysis of variance. The results were as follows: genotype +/+: F = 0.97, df1 = 2, df2 = 53, P = 0.39; genotype D2/D3: F = 2.45, df1 = 2, df2 = 56, P = 0.095. The significance of the difference of the mean values mentioned was calculated according to the Student’s t-test, the result of which was as follows: t = 5.19, df = 115 and P < 0.0001. In addition, the P-values for the test of heterogeneity are given below the group of columns for the respective genotype, and that for the test of the difference between the genotypes in the upper right corner of the figure.
The coefficients of coincidence in each temperature for the control and experimental crosses are given in (Figure 8). The coefficient was not dependent on the temperature in either group of crosses. However, its mean value in the experimental crosses was significantly higher than in the control crosses (P < 0.0001).
The values of all the parameters measured calculated directly on the basis of the total numbers of progeny in the control and experimental crosses and their comparisons are given in (Table 3). The significances of the differences are the same as those calculated on the basis of the mean values per female or per culture bottle, given in the figures, except that the comparison of the recombination frequencies of the cv and v markers gave no significant difference. However, essentially, the two methods of the comparison of the mus309 genotypes studied, gave similar results.
In general, the main features of the results are as follows: None of the parameters measured was dependent on the temperature in the control crosses involving nonmutant mus309 females. In the experimental crosses involving mutant mus309 females all the parameters,

Figure 7. The effect of temperature on the map distance between the cv and f markers of the X chromosome in the progeny of the mus309+/mus309+ (+/+) and mus309D2/mus309D3 (D2/D3) females raised in three different temperatures, viz. 18˚C, 25˚C and 29˚C. The columns represent the mean map distances in centimorgans (cM) per culture bottle and the vertical lines their standard deviations. The mean map distance together with its standard deviation for both genotypes is also given and indicated by the red diamond (♦). Testing of the heterogeneity of the results inside each mus309 genotype was carried out using the one-way analysis of variance. The results were as follows: genotype +/+: F = 1.63, df1 = 2, df2 = 53, P = 0.21; genotype D2/D3: F = 6.16, df1 = 2, df2 = 56, P = 0.0038. The significance of the difference of the mean values mentioned was calculated according to the Student’s t-test, the result of which was as follows: t = 7.97, df = 114 and P < 0.0001. In addition, the P-values for the test of heterogeneity are given below the group of columns for the respective genotype, and that for the test of the difference between the genotypes in the upper right corner of the figure.
except the frequency of true single crossovers in the cv-v interval, the recombination frequency of the v and f markers, and the coefficient of coincidence, were dependent on the temperature. When comparing the genotypes studied, a significant difference between them was observed in all the parameters studied, except for the frequency of the true single crossovers in the cv-v interval. As explained in the following discussion, these results support the counting number model of the mechanism of interference based on the genetic distance, but are in contradiction with the models based on physical distance.
4. DISCUSSION
4.1. The Effect of Temperature and the mus309 Mutation on Fecundity
The decrease in and temperature sensitivity of the fecundity of the experimental females, as compared to the control females, is interesting. It is logical to assume that the formation of the DSBs and their subsequent repair is temperature-dependent in the mus309 mutants, which are deficient in such repair. It is therefore suggested that the decrease in the fertility observed is due to a complete

Figure 8. The effect of temperature on the crossover interference in the cv-v-f region of the X chromosome in the progeny of the mus309+/mus309+ (+/+) and mus309D2/mus309D3 (D2/D3) females raised in three different temperatures, viz. 18˚C, 25˚C and 29˚C. The columns represent the mean coefficients of coincidence (C) per culture bottle and the vertical lines their standard deviations. The mean value of C calculated on the basis of the total number of progeny of the females of the genotype given together with its standard deviation, which was calculated according to the formula given in material and methods, for both genotypes is also given and indicated by the red diamond (♦). Testing of the heterogeneity of the results inside each mus309 genotype was carried out using the one-way analysis of variance. The results were as follows: genotype +/+: F = 1.29, df1 = 2, df2 = 53, P = 0.28; genotype D2/D3: F = 1.65, df1 = 2, df2 = 56, P = 0.20. The significance of the difference of the mean values mentioned was calculated according to the Student’s t-test, the result of which was as follows: t = 8.58, df = 12160 and P < 0.0001. In addition, the P-values for the test of heterogeneity are given below the group of columns for the respective genotype, and that for the test of the difference between the genotypes in the upper right corner of the figure.
failure of the repair of the DSBs, thus leading to sterility of the female or lethality of its progeny [13]. It is, in fact, known that a defect in DSB repair activates a signaling pathway that leads to defects later in the oocyte development [32-35]. Moreover, it is also logical to assume that failure of the repair is the more complete the higher the temperature, as the results suggest (Figure 1). These ideas are also consistent with the suggestions which will be presented below on the effect of the temperature on the crossing over frequencies.
4.2. The Effect of the Temperature and the mus309 Gene on Crossing Over and Crossover Interference
It is convincingly established that those meiotic mutants of Drosophila melanogaster affecting crossing over which also affect interference involve preconditions of crossing over, whereas those mutants that affect crossing over without affecting interference involve the crossing over event itself [36]. Consequently, the genes involved are called precondition genes and exchange genes, respectively.
This was theoretically shown as follows: Let a be the probability of the fulfillment of preconditions of crossing over in one region and only in that region in a three-point crossing-over experiment. Let b be the probability of fulfillment of the same in another region and only in that region. Let d be the probability of the fulfillment of the preconditions in both regions at the same time, and x the probability of exchange, given the preconditions. Then the coefficient of coincidence, C, is
 (3)
(3)
Since C is independent of x, if a mutant that acts on crossing over also affects interference, it must influence the preconditions of crossing over. If, however, interfereence remains unaltered, the target of the effect is the exchange itself [37].
What in this respect is true for meiotic mutants is, of course, also true for other factors that affect crossing over, such as temperature in the present study.
As has been shown earlier [25,38], and was also observed in this study, the mus309 mutations affect both crossing over and crossover interference and, accordingly, involve some precondition of crossing over belonging to the class that has been referred to as the “precondition mutants”, meaning that they act prior to the time when crossovers are actually generated [39].
As indicated in the introduction, the precondition of crossing over, which the mus309 gene product affects, is the repair of DSBs—a necessary condition for crossing over. In particular, it is known that the MUS309 protein is involved in the SDSA pathway of the repair of the DSBs. More specifically, it is also clear that in the mus309 mutants the SDSA pathway is blocked, while the DSBR pathway remains functional.
In the experimental crosses temperature affected crossing-over frequencies, the double crossovers included, but it did not affect interference. In the control crosses, however, neither crossing over frequencies nor interfereence was affected by the temperature. This indicates that temperature affects the event of crossing over itself, but only in combination with the mus309 mutation. This suggests that temperature affects the formation of DNA double-strand breaks.
4.3. Test of the Models of Crossover Interference
As mentioned in the introduction, models of crossover interference can, in principle, be divided into two different categories. The first category of models, called genetic models [4], assumes that interference is dependent on genetic (i.e. linkage map) distance (Morgans) between adjacent crossovers. To my knowledge, currently only one model, called the “counting model” [4,5], falls into this category.
The central feature of the counting model is that recombinational intermediates (C’s) have two fates – they can be resolved with crossing over (Cx) or without (Co). The C’s are distributed at random with respect to each other, and interference results from constraints on the resolution of C’s. The basic constraint is that each pair of neighboring Cx’s must have a certain number, m, of Co’s between them, as if the meiocyte was able to ‘count’ recombination events.
The second category of models, which may be called physical models, hypothesizes that crossover interfereence is dependent on physical distance (microns or base pairs) between the adjacent crossovers. In general, these models suggest that some kind of physical signal travels along the bivalent and determines the distribution of crossovers. One of the models belonging to this category, the reaction-diffusion model [40], is quantitative while the other models are qualitative.
According to the reaction-diffusion model, a “random walking” precursor becomes immobilized and matures into a crossover point. The interference is caused by a pair-annihilation of the random walkers, called the A particles, due to their collision together, or by annihilation of a random walker due to its collision with an immobilized point. This model has two parameters—the initial density of the random walkers, α, and the rate, h, of their processing into crossover points. It is logical to conclude that interference decreases if the α value increases and/or h decreases [40].
It is also quite logical to assume that if the mus309 mutations affect the balance by which the double Holliday junctions will be resolved as crossovers instead of non-crossovers the m value of the counting model should decrease in mus309 mutants [25]. The results of the present study are consistent with this idea.
In contrast, however, the results of the present study are not compatible with the reaction-diffusion model. According to this model, interference depends on two factors only, viz. the initial density of crossover precursors, i.e. DSBs, and the rate of their processing into crossovers. Therefore, it is hard to conceive, in terms of the reaction-diffusion model, how the number of crossovers, i.e. the map distances, would change due to the effect of temperature but their distances, i.e. interference, would not, as the initial density of DSBs does not change. This seems, however, to be the case in the results of the present study.
The results are also in contradiction with any model of crossover interference based on physical distance on the following grounds: the map distances in the experimental and control females are different, and react differently to temperature, the map distances in the experimental crosses being temperature sensitive while the distances in the control crosses are not. However, the crossover interference is independent of the temperature in both series of crosses. As explained above, this observation supports the models of interference based on genetic distance. On the other hand, the results are in contradiction with the models based on physical distance. In fact, if interference was dependent on physical distance, how could it remain unchanged despite the temperature when the genetic map distances change but the physical distances do not?
5. ACKNOWLEDGEMENTS
Thanks are given to Professor Janos Szabad (Szeged, Hungary) for introducing me to the mus309 gene, and the generous donation of the mutant stocks which, however, are readily available in stock centers. Skilful technical assistance by Mirja Rantanen, M.Sc. is gratefully acknowledged. Docent Kai Ruohomäki, PhD helped me by conducting the statistical analyses, and Kurt Ståhle, the technician, by drawing the figures. To all of them I am very grateful. Special thanks are given to Maaria Tringham, M.Sc., and Damon Tringham, M.Phil., for checking the language.
REFERENCES
- Sturtevant, A.H. (1915) The behavior of the chromosomes as studied through linkage. Zeitschrift für Inductive Abstammuns und Vererbungslehre, 13, 234-287.
- Muller, H.J. (1916) The mechanism of crossing over. American Naturalist, 50, 193-221. doi:10.1086/279534
- Hillers, K.J. (2004) Crossover interference. Current Biology, 14, R1036-R1037. doi:10.1016/j.cub.2004.11.038
- Foss, E., Lande, R., Stahl, F.W. and Steinberg, C.M. (1993) Chiasma interference as a function of genetic distance. Genetics, 133, 681-691.
- Mortimer, R.K. and Fogel, S. (1974) Genetical interference and gene conversion. In: Grell, R.F., Ed., Mechanims in Recombination, Plenum Press, New York, 263-275.
- Bridges, C.B. (1915) A linkage variation in Drosophila. Journal of Experimental Zoology, 19, 1-21. doi:10.1002/jez.1400190102
- Sandler, L. and Lindsley, D.L. (1974) Some observations on the study of the genetic control of meiosis in Drosophila melanogaster. Genetics, 78, 289-297.
- Lifschytz, E. (1975) Differential sensitivities and the target of heat-induced recombination at the base of the X chromosome of Drosophila melanogaster. Genetics, 79, 283-294.
- McKim, K.S. and Hayashi-Hagihara, A. (1998) Mei-W68 in Drosophila melanogaster encodes a Spo11 homolog: evidence that the mechanism for initiating meiotic recombination is conserved. Genes and Development, 12, 2932-2942. doi:10.1101/gad.12.18.2932
- Keeney, S., Giroux, C.N. and Kleckner, N. (1997) Meiosis-specific DNA double-strand breaks are catalyzed by Spo11, a member of a widely conserved protein family. Cell, 88, 375-384. doi:10.1016/S0092-8674(00)81876-0
- Keeney, S. (2001) Mechanism and control of meiotic recombination. Current Topics in Developmental Biology, 52, 1-53. doi:10.1016/S0070-2153(01)52008-6
- Keeney, S. and Neale, M.J. (2006) Initiation of meiotic recombination by formation of DNA double-strand breaks: Mechanism and regulation. Biochemical Society Transactions, 34, 523-525. doi:10.1042/BST0340523
- Joyce, E.F. and McKim, K.S. (2009) Drosophila PCH2 is required for a pachytene checkpoint that monitors double-strand-break-independent events leading to meiotic crossover formation. Genetics, 181, 39-51. doi:10.1534/genetics.108.093112
- Olsen-Krogh, B. and Symington, L.S. (2004) Recombination proteins in yeast. Annual Reviews of Genetics, 38, 233-271. doi:10.1146/annurev.genet.38.072902.091500
- Lorenz, A. and Whitby, M.C. (2006) Crossover promotion and prevention. Biochemical Society Transactions, 34, 537-541. doi:10.1042/BST0340537
- Heyer W.-D., Ehmsen, K.T. and Solinger, J.A. (2003) Holliday junctions in eukaryotic nucleus: Resolution in sight? Trends in Biochemical Sciences, 28, 548-557. doi:10.1016/j.tibs.2003.08.011
- Heyer, W.-D. (2004) Recombination: Holliday junction resolution and crossover formation. Current Biology, 14, R56-R58. doi:10.1016/j.cub.2003.12.043
- Ellis, N.A., Groden, J., Ye, T-Z., Staughen, J., Lennon, D.J., Ciocci, S., Proytcheva, M. and German, J. (1995) The bloom’s syndrome gene-product is homologous to RecQ helicases. Cell, 83, 655-666. doi:10.1016/0092-8674(95)90105-1
- Karow, J.K., Chakraverty, R.K. and Hickson, J.D. (1997) The bloom’s syndrome gene product is a 3’- 5’ DNA helicase. Journal of Biological Chemistry, 272, 30611-30614. doi:10.1074/jbc.272.49.30611
- Mohaghegh, P., Karow, J.K., Brosh, R.M. Jr., Bohr, V.A. and Hickson, I.D. (2001) The bloom’s and Werner’s syndrome proteins are DNA structure-specific homologues. Nucleic Acids Research, 29, 2843-2849. doi:10.1093/nar/29.13.2843
- Wu, L., Davies, S.L., Levitt, N.C. and Hickson, I.D. (2001) Potential role for the BLM helicase in recombinetional repair via a conserved interaction with RAD51. Journal of Biological Chemistry, 276, 19375-19381. doi:10.1074/jbc.M009471200
- Brabant, A.J., van Stan, R. and Ellis, N.A. (2000) DNA helicases, genome instability, and human genetic disease. Annual Reviews of Genomics and Human Genetics, 1, 409-459.
- Adams, M.D., McVey, M. and Sekelsky, J.J. (2003) Drosophila BLM in double-strand break repair by synthesis-dependent strand annealing. Science, 299, 265-267. doi:10.1126/science.1077198
- Laurencon, A., Orme, C.M., Peters, H.K., Boulton, C.L., Vladar, E.K., Langley, S.A., Bakis, E.P., Harris, D.T., Harris, N.J., Wayson, S.M., Hawley, R.S. and Burtis, K.C. (2004) A large-scale screen for mutagen sensitive loci in Drosophila. Genetics, 167, 217-231. doi:10.1534/genetics.167.1.217
- Portin, P. (2005) mus309 mutation, defective in DNA double-strand break repair, affects intergenic but not intragenic meiotic recombination in Drosophila melanogaster. Genetical Research, 86, 185-191. doi:10.1017/S0016672305007883
- Rockmill, B., Fung, J.C., Branda, S.S. and Roeder, G.S. (2003) The Sgs1 helicase regulates chromosome synapsis and meiotic crossing over. Current Biology, 13, 1954- 1962. doi:10.1016/j.cub.2003.10.059
- Kusano, K., Johnson-Schlitz, D.M. and Engels, W.R. (2001) Sterility of Drosophila with mutations in the Bloom syndrome gene complementation by Ku70. Science, 291, 2600-2602. doi:10.1126/science.291.5513.2600
- Boyd, J.B., Golino, M.D., Shaw, K.E.S., Osgood, C.J. and Green, M.M. (1981) Third-chromosome mutagensensitive mutants of Drosophila melanogaster. Genetics, 97, 607-623.
- Beal, E.L. and Rio, D.C. (1996) Drosophila IRBP / Ku p70 corresponds to the mutagen-sensitive mus309 gene and is involved in P-element excision in vivo. Genes and Development, 10, 921-933. doi:10.1101/gad.10.8.921
- Weinstein, A. (1936) The theory of multiple-strand crossing over. Genetics, 21, 155-199.
- Stevens, W.L. (1936) The analysis of interference. Journal of Genetics, 32, 51-64. doi:10.1007/BF02982501
- Ghabrial, A. and Schüpbach, T. (1999) Activation of meiotic checkpoint regulates translation of Gurken during Drosophila oogenesis. Nature Cell Biology, 1, 354- 357. doi:10.1038/14046
- Ghabrial, A., Ray, R.P. and Schüpbach, T. (1998) Okra and spindle-B encode components of the RAD52 DNA repair pathway and affect meiosis and patterning in Drosophila oogenesis. Genes and Development, 12, 2711- 2713. doi:10.1101/gad.12.17.2711
- Abdu, U., Gonzalez-Reyes, A., Ghabrial, A. and Schüpbach, T. (2003) The Drosophila spn-D gene encodes a RAD51C-like protein that is required exclusively during meiosis. Genetics, 165, 197-204.
- Staeva-Vieira, E., Yoo, S. and Lehmann, R. (2003) An essential role of DmRad51/SpnA in DNA repair and meiotic checkpoint control. EMBO Journal, 22, 5863- 5874. doi:10.1093/emboj/cdg564
- Baker, B.S. and Hall, J.C. (1976) Meiotic mutants: Genetic control of meiotic recombination and chromosome segregation. In: Ashburner, M. and Novitski, E., Eds., The Genetics and Biology of Drosophila, Academic Press, San Francisco, 351-434.
- Sandler, L., Lindsley, D.L., Nicoletti, B. and Trippa, G. (1968) Mutants affecting meiosis in natural populations of Drosophila melanogaster. Genetics, 60, 525-558.
- McVey, M., Andersen, S.L., Broze, Y. and Sekelsky, J.J. (2007) Multiple functions of Drosophila Blm helicase in maintenance of genome stability. Genetics, 176, 1979- 1992. doi:10.1534/genetics.106.070052
- Baker, B.S. and Carpenter, A.T.C. (1972) Genetic analysis of sex chromosomal meiotic mutants in Drosophila melanogaster. Genetics, 71, 255-286.
- Fujitani, Y., Mori, S. and Kobayashi, I. (2002) A reaction-diffusion model for interference in meiotic crossing over. Genetics, 161, 365-372.