Intracellular Transport of HIV-1 Matrix Protein Associated with Viral RNA
Copyright © 2013 SciRes. WJA
35
get the fractions of cytosol, membranes, nuclei and cy-
toskeleton. Localization of MA in cellular fractions was
determined using immunoblot.
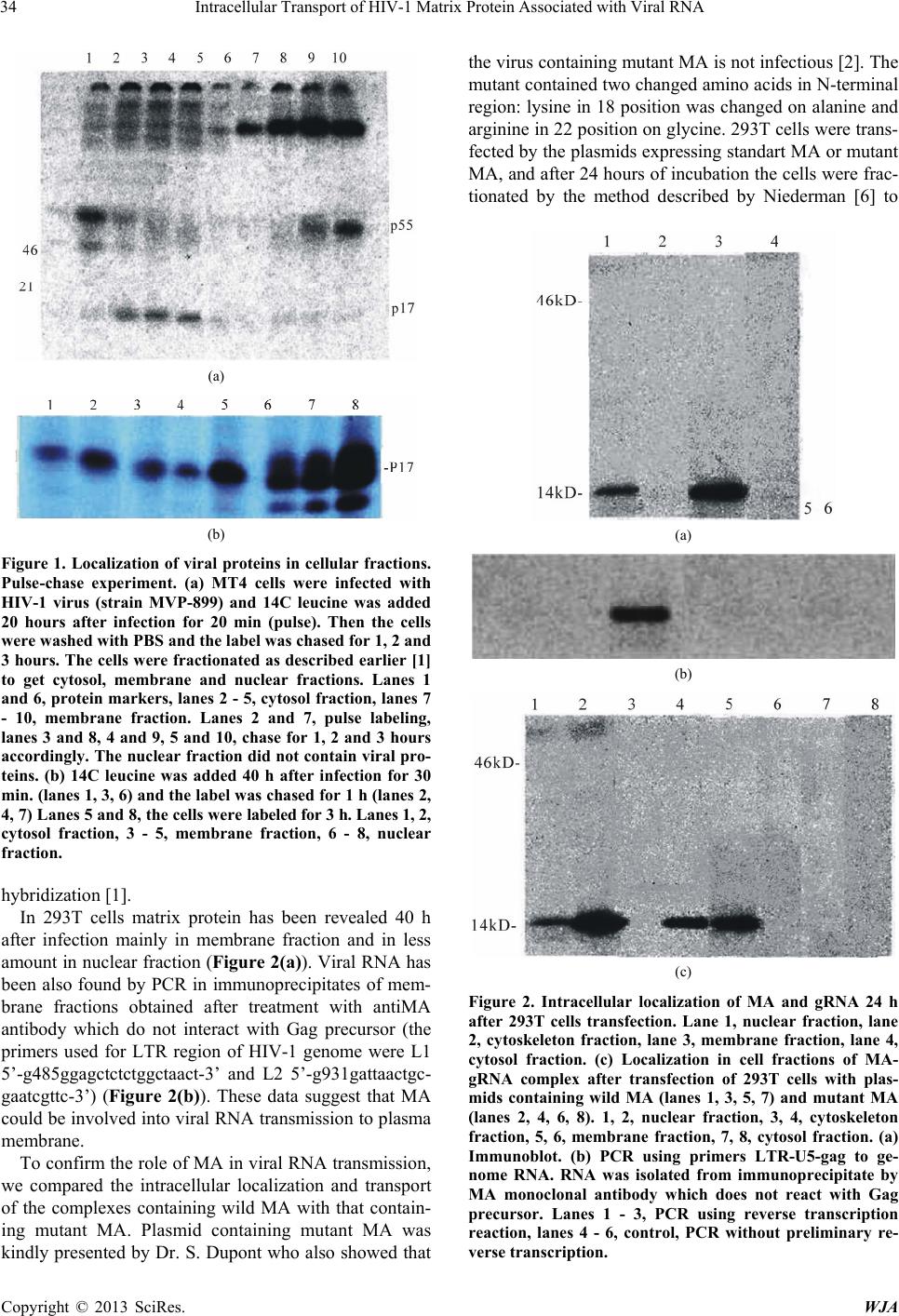
After transfection with plasmid expressing standard
MA, the small amount of MA was found in the nuclear
fraction while the main part of the protein was localized
in the membrane fraction. However, after transfection
with plasmid expressing mutant M4 matrix the protein
was not found in the membrane fraction but was local-
ized in the nuclear and cytoskeleton fractions (Figure
2(c)). These data show that the association of the mutant
MA with membranes is blocked, probably because mu-
tant MA lost its membranophylic signal (myristic acid)
on its N terminus, and the mutant MA protein associated
with viral RNA “sticks” in nuclei and cytoskeleton, the
fractions of its previous location. These results allow
suggesting that the complex MA-viral RNA is trans-
ported from the nuclei to membranes by cytoskeleton.
The reverse transport from membranes to nuclei was
described for viral reverse transcription and preintegra-
tion complexes [7] showing that the virus widely uses
cytoskeleton for its intracellular transport.
2. Conclusions
Despite the progress in defining determinates of Gag
participation in HIV assembly, Gag intracellular interac-
tions and trafficking to the assembly sites in the infected
cells are poorly understand. Our results indicate, that MA
is connected with viral RNA early after infection and the
complex is transported to the cellular membrane—the
place of viral assembly. When the mutant MA was used
instead of standard MA, the complex of MA and viral
RNA was not found in the cellular membrane, but sticks
in nucleus and cytoskeleton fractions. These results show
that MA-RNA complex is transported from the nuclei to
the cellular membrane by cytoskeleton.
The data also show that MA is cleaved from Gag pre-
cursor very early after Gag synthesis and thus could be
involved in to virus assembly as an individual protein.
The significant role of MA in HIV-1 replication shows
that this protein is the candidate antigen for therapeutic
vaccine against AIDS.
REFERENCES
[1] A. G. Bukrinskaya, G. K. Vorkunova and Y. Y. Tentsov,
“HIV-1 Matrix Protein P17 Resides in Cell Nuclei in As-
sociation with Genomic RNA,” AIDS Research and Hu-
man Retroviruses, Vol. 8, No. 10, 1992, pp. 1795-1801.
doi:10.1089/aid.1992.8.1795
[2] S. Dupont, N. Sharova, C. DeHoratius, C. M. A. Vir-
basius, X. C. Zhu, A. G. Bukrinskaya, M. Stevenson and
M. R. Green, “A Novel Nuclear Export Activity in HIV-1
Matrix Protein Required for Viral Replication,” Nature,
Vol. 402, No. 6762, 1999, pp. 681-685.
doi:10.1038/45272
[3] A. H. Kaplan and R. Swanstrom, “HIV-1 Gag Proteins
Are Processed in Two Cellular Compartments,” Procee-
dings of the National Academy of Sciences of USA, Vol.
88, No. 10, 1991, pp. 4528-4532.
doi:10.1073/pnas.88.10.4528
[4] A. Bukrinskaya, “HIV-1 Matrix Protein: A Mysterious
Regulator of the Viral Life Cycle,” Virus Research, Vol.
124, No. 1-2, 2007, pp. 1-11.
doi:10.1016/j.virusres.2006.07.001
[5] N. Sharova and A. Bukrinskaya, “P17 and P17-Con-
taining Gag Precursor of Input HIV Are Transported into
the Nuclei of Infected Cells,” AIDS Research and Human
Retroviruses, Vol. 7, No. 3, 1991, pp. 303-306.
doi:10.1089/aid.1991.7.303
[6] T. M. J. Niederman, W. R. Hastings and L. Ratner, “My-
ristoylation-Enhanced Binding of the HIV-1 Net Protein
to T Cell Skeletal Matrix,” Virology, Vol . 1 9 7 , N o . 1 , 1993,
pp. 420-425. doi:10.1006/viro.1993.1605
[7] A. Bukrinskaya, B. Brichacek, A. Mann and M. Steven-
son, “Establishment of a Functional HIV-1 Reverse Tran-
scription Complex Involves the Cytoskeleton,” The Jour-
nal of Experimental Medicine, Vol. 188, No. 11, 1998, pp.
2113-2125. doi:10.1084/jem.188.11.2113