M. Chandrasekharan et al. / J. Biomedical Science and Engineering 6 (2013) 209-212
212
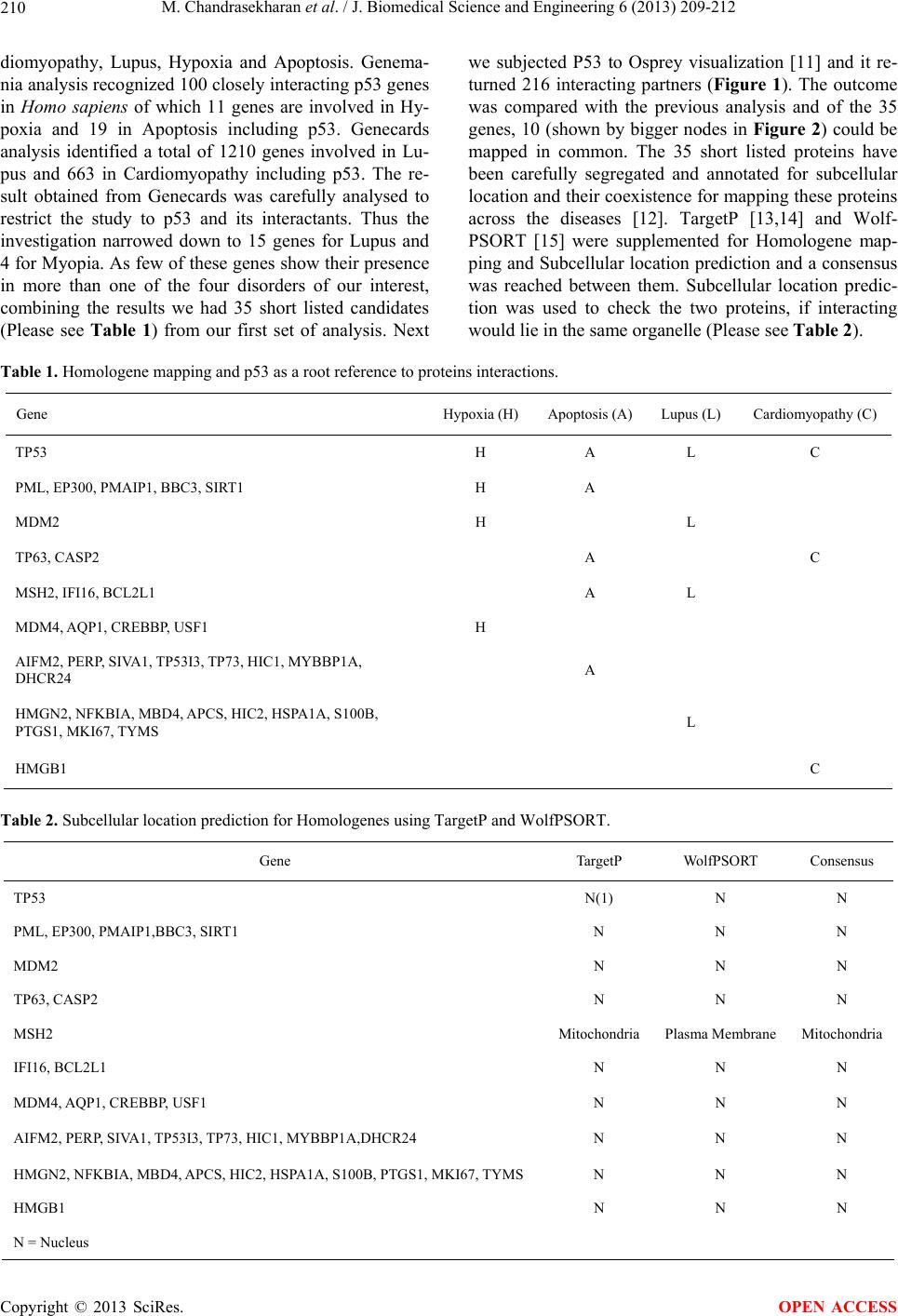
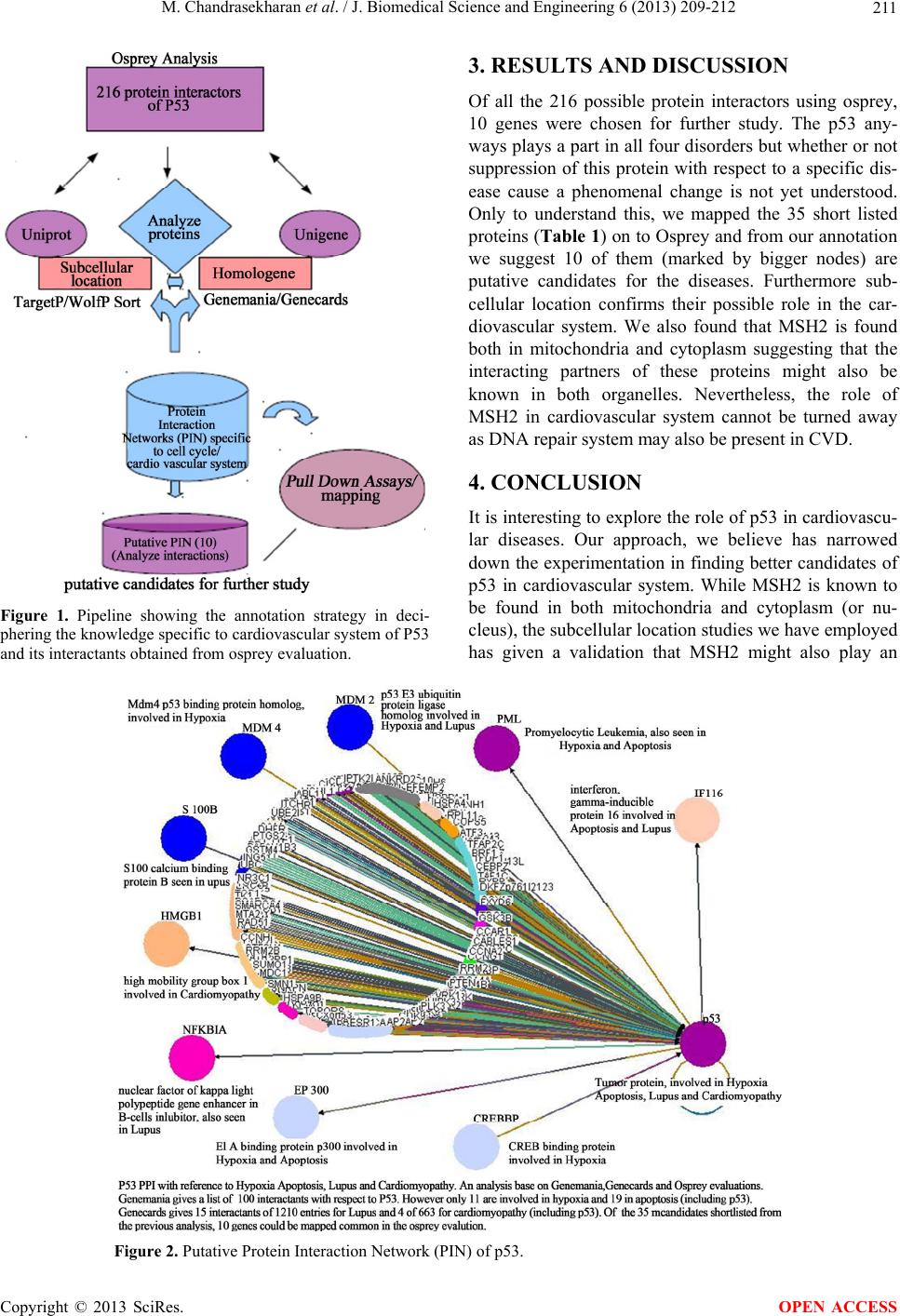
important role in validating the role of these proteins
linked to p53, towards CVD. We believe and envisage
that the other proteins (highlighted as bold and bigger
nodes) are better candidates and it would be interesting
to run pull down assays for these proteins to check for
their candidacy in CVD.
5. ACKNOWLEDGEMENTS
We thank Dr. Tiratha Raj Singh for providing kind inputs.
REFERENCES
[1] Bai, L. and Zhu, W.-G. (2006) p53: Structure, function
and therapeutic applications. Journal of Cancer Mole-
cules, 2, 141-153.
[2] National Center for Biotechnology Information (US)
(1998) The p53 tumor suppressor protein. Genes and
Disease, National Center for Biotechnology Information
(US), Bethesda.
http://www.ncbi.nlm.nih.gov/books/NBK22268/
[3] Levine, A.J. and Oren, M. (2009) The first 30 years of
p53: Growing ever more complex. Nature Reviews Can-
cer, 9, 749-758. doi:10.1038/nrc2723
[4] Borradaile, N.M. and Pickering, J.G. (2009) NAD(+),
sirtuins, and cardiovascular disease. Current Pharmaceu-
tical Design, 15, 110-117.
doi:10.2174/138161209787185742
[5] Sano, M. and Komuro, I. (2008) P53 and its role in the
development of heart failure. Japanese Journal of Clini-
cal Medicine, 66, 1013-1021.
[6] Sano, M., Minamino, T., Toko, H., Miyauchi, H., Orimo,
M., Qin, Y., Akazawa, H., Tateno, K., Kayama, Y., Ha-
rada, M., et al. (2007) p53-induced inhibition of hif-1
causes cardiac dysfunction during pressure overload. Na-
ture, 446, 444-448.
[7] Tsoporis, J.N., Mohammadzadeh, F., Parker, T.G. (2011)
S100B: A multifunctional role in cardiovascular patho-
physiology. Amino Acids, 41, 843-847.
doi:10.1007/s00726-010-0527-1
[8] Mercer, J., Mahmoudi, M. and Bennett, M. (2007) DNA
damage, p53, apoptosis and vascular disease. Mutation
Research, 621, 75-86.
doi:10.1016/j.mrfmmm.2007.02.011
[9] Warde-Farley, D., Donaldson, S.L., Comes, O., Zuberi,
K., Badrawi, R., Chao, P., Franz, M., Grouios, C., Kazi,
F., Lopes, C.T., Maitland, A., Mostafavi, S., Montojo, J.,
Shao, Q., Wright, G., Bader, G.D. and Morris, Q. (2010)
The GeneMANIA prediction server: Biological network
integration for gene prioritization and predicting gene
function. Nucleic Acids Research, 38, W214-W220.
doi:10.1093/nar/gkq537
[10] Stelzer, G., Dalah, I., Iny Stein, T., Satanower, Y., Rosen,
N., Nativ, N., Oz-Levi, D., Olender, T., Belinky, F., Bahir,
I., Krug, H., Perco, P., Mayer, B., Kolker, E., Safran, M.
and Lancet, D. (2011) In-silico human genomics with
genecards. Human Genomics, 5, 709-717.
doi:10.1186/1479-7364-5-6-709
[11] Breitkreutz, B.J., Stark, C. and Tyers, M. (2003) Osprey:
A network visualization system. Genome Biology, 4, R22.
doi:10.1186/gb-2003-4-3-r22
[12] Nemchenko, A., Chiong, M., Turer, A., Lavandero, S.
and Hill, J.A. (2011) Autophagy as a therapeutic target in
cardiovascular disease. Journal of Molecular and Cellu-
lar Cardiology, 51, 584-593.
doi:10.1016/j.yjmcc.2011.06.010
[13] Emanuelsson, O., et al. (2007) Locating proteins in the
cell using TargetP, SignalP and related tools. Nature
protocols, 2, 953-971. doi:10.1038/nprot.2007.131
[14] Emanuelsson, O., Nielsen, H., Brunak, S. and von Heijne,
G. (2000) Predicting subcellular localization of proteins
based on their N-terminal amino acid sequence. Journal
of Molecular Biology, 300, 1005-1016.
doi:10.1006/jmbi.2000.3903
[15] Horton, P., Park, K.-J., Obayashi, T., Fujita, N., Harada,
H., Adams-Collier, C.J. and Nakai, K. (2007) WoLF
PSORT: Protein localization predictor. Nucleic Acids Re-
search, 35, W585-W587. doi:10.1093/nar/gkm259
Copyright © 2013 SciRes. OPEN ACCESS