American Journal of Plant Sciences
Vol.07 No.06(2016), Article ID:66215,10 pages
10.4236/ajps.2016.76094
Synergistic Effect of Watermelon Powder and Lactococcus lactis subsp lactis Supplemented Diet Partially Ameliorates Aβ42-Dependent Lifespan Shorten and Flight Impairment in Transgenic Drosophila
Dong Gyun Ko1, Young Bum Eun1, Jong Uk Na1, Sang-Tae Kim2*
1Korean Minjok Leadership Academy, Hoengseong-gun, Republic of Korea
2Biomedical Research Institute, Department of Neuropsychiatry, Bundang Hospital of Seoul National University, College of Medicine, Seongnam, Republic of Korea

Copyright © 2016 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 1 February 2016; accepted 26 April 2016; published 29 April 2016
ABSTRACT
Oxidative stress has been strongly related with Alzheimer disease pathogenesis. We determined the effects of watermelon powder (WMP) and Lactococcus lactis subsp lactis (LAL) supplementation on the generated Aβ42-induced phenotypes in a Drosophila melanogaster model of AD. Enhanced Aβ42 expression in D. melanogaster neurons can diminish lifespan and flight ability. We have observed longevity methods to assay the effects of WMP and LAL on D. melanogaster survival. Furthermore, flies expressing Aβ42 in their body fed WMP and LAL had up to 90 days, or 35% longer median lifespan than those fed a control diet. In addition, synergistic effect of WMP and LAL improved Aβ42-induced flight impairments in the Drosophila wing tissues. Our microscope experiments revealed that individuals fed synergistic effect of WMP and LAL had ameliorated Aβ42 expression as well as increment of flight ability than Aβ42-induced flies. We propose that WMP is typically rich in L-citrulline and LAL, rich in naturally occurring probiotics and antioxidants, and that it promotes the survival of neurons in brain and wing muscle tissues with increased levels of Aβ42 via a protective cell survival mechanism.
Keywords:
Drosophila melanogaster, Lactococcus lactis, Watermelon, Lifespan, Ommatidia, Aβ42

1. Introduction
Alzheimer’s disease (AD) is an age-related neurodegenerative disease and the most common cause of dementia. The pathogenesis of AD is yet entirely clear and despite the increasing knowledge regarding the mechanism, no effective disease-modifying therapy is available. Recently, many reports have shown to play a pivotal role in the synaptic damage, impairment of homeostasis, inflammation as well as toxicity in relation to AD etiology. Membranes can also be injured by the ROS (reactive oxygen species) that are produced by Aβ aggregation in the presence of metals such as copper, zinc or iron [1] . Subsequent pathophysiological processes include mitochondrial damage [2] , phosphor-tau with consequent axonal transport damage and trigger of cell death [3] [4] . However, until recent it was impossible to take a global view to ask which biological processes are essential for the induction of the disease and which are downstream consequences of cytotoxicity [5] . Knowing which biological processes are directly involved in initiating AD will allow us to key on those upstream targets that have the greatest therapeutic potential.
Oxidative stress (OS) is consistently related with the etiology of AD; however, its role in disease progression remains unclear. A cell undergoes OS when the net balance between the generated reactive oxygen species (ROS) and the available antioxidant defense mechanisms favors the former. Autopsy analysis of AD patients’ brains certifies higher levels of OS biomarkers like dysfunctional mitochondria, decreased levels of reduced glutathione, and deficiencies in antioxidant enzymes in the hippocampus of affected individuals [5] . Recent report suggests that production of H2O2 in Arctic Aβ42 flies is a vital step in the oxidative stress caused by the Aβ and is susceptible to OS [6] [7] . This data suggest that the combination of OS and excess Tau and Aβ42 may play a pivotal role in the progression of AD.
Watermelon (WM, Citrullus lanatus) is typically rich in l-citrulline, varying in amounts from 0.7 to 3.6 mg/g fresh [8] , and can possibly produce similar or better effects than l-arginine supplementation. Studies have found that C. lanatus juice retains antioxidant and anti-inflammatory properties [9] [10] . Watermelon is already known to play a protective role in reducing oxidative stress through the phytochemical lycopene [11] . Watermelon is also high in other antioxidants, which have been linked to decreased risk of coronary heart disease [12] . Citrullus lanatus juice supplementation was also shown to raise serum levels of l-arginine and l-citrulline as effectively as elemental supplementation [8] . Also, l-arginine supplementation rescues the effects of an oxidative stress by decreasing inflammation and increasing antioxidant status [11]
Various studies have determined the impacts of individual components of C. lanatus; however, little has been done to observe the effects of whole watermelon supplementation in powder form on lipid profiles, antioxidant ability, and inflammation [12] . It remains unclear whether watermelon powder supplementation has the same beneficial effects for antioxidant and anti-inflammatory properties on neuronal degenerations as elemental supplementation of these bioactive components. The purpose of this study was to examine the effects of l-citrulline- rich watermelon powder supplementation on antioxidant capacity and inflammation.
Lactococcus lactis subsp lactis (LAL) is an excellent source of dietary antioxidants. The scavenger potential of LAL in OS and vascular disease has been described and recent studies in Drosophila suggest that microbiota may be beneficial to individuals suffering from neurodegenerative diseases [13] . Therefore, in this study we describe the protective effects of LAL concentrate on a Drosophila transgenic (Tg) model of AD. Extension of lifespan and enhanced flight impairments caused by directed expression of Aβ42 in the brain and developing eye, respectively, is improved by supplementing growth media with LAL.
We have designed a model of AD that is based on the expression of the human Aβ42 in fly neurons by coupling it to an N-terminal secretion signal peptide [14] . The Aβ1-42 but not the Aβ1-40 control accumulates in the brain and results in decreased lifespan and impaired locomotor performance. These phenotypes are more marked in Tg (transgenic) flies expressing the mutant of the Aβ1-42. It causes increased aggregation of Aβ and is responsible for early stage familial AD [15] . We hypothesized that WBP and LAL supplementation would extend lifespanand improve flight impairment for AD by promoting positive effects on antioxidant and neuronal defect. Here, Drosophila Tg (transgenic) model with AD can help unravel the role of OS and unveil potential antioxidant therapies in AD that are critical for lifespan loss and flight impairments from Aβ-mediated neurotoxicity with 1.35 g/Lor 3 g/L of WMP, and 3 g/L of WMP and 5 × 108 100 μl Lactococcus lactis subsp lactis (LAL) supplementation in vivo.
2. Materials and Methods
2.1. Fly stocks and Reagents
UAS-Aβ [16] flies were generously provided by Professor G.S. Jo (University of Kunkuk), respectively. GMR- Gal412 and UAS-mCD8-GFP flies were obtained from the Bloomington Drosophila Stock Center at Indiana University. Adult-onset, eye-specific expression of Aβ42 peptide was achieved by using the GMR driver (GMR- Gal4/UAS system). Control flies were generated by crossing tissue specific drivers to Green Fluorescent Protein (UAS-mCD GFP) for suppressive experiments. To extend these findings and in consideration of the need to develop tractable animal models to study neuronal diseases, we employed the UAS-GAL4 system to examine the beneficial effect of antioxidant and neuronal damage. The homozygous GMR-GAL4 driver was crossed to homozygous UAS lines and the progeny were subjected to study. The same procedure was followed with the UAS-mCD GFP; GMR-Gal4 (glass multiple reporter) promoter elements, has been commonly utilized to express target transgenes, specifically in the developing eye. Crosses were set up in standard food vials and environmental conditions, and the flies were passed every three days.
The transgenes are each representative of three independent transgenic lines. The watermelon powder consisted of freeze-dried watermelon solids that had been sieved and was originally manufactured by Milne Fruit Products (Prosser, WA) [16] . Casein (Halran, Madison, WI) contains 3.4% of l-arginine; therefore, the control diet contains 6.8 g l-arginine and 0 g l-citrulline per kilogram diet. The watermelon powder contains l-citrulline (1.35 g) and l-arginine (0.65 g) for 3.3 g of the powder (WMP). Watermelon diet provides l-arginine (7.45 g) and l-citrulline (1.35 g and 3 g) per kilogram watermelon-supplemented diet. Flies were maintained at 25˚C in a 12:12 light:dark period on a standard cornmeal-yeast-molasses-agar medium (65 g/L cornmeal, 15 g/L nutritional yeast extract, 5.5 g/L agar in water supplemented with 0.1 g/ml methyl 4-hydroxybensoate in ethanol and 2.5 ml propionic acid per L of medium).
2.2. Longevity Assays
Longevity assays in the secondary screen and flies were reared on either 0.25% sucrose or (1.35 g and 3 g) WMP and 5 × 108 100 μl LAL supplemented medium was then collected under gaseous CO2 every 24 hours until a minimum of 100 adult females of each genotype were obtained. In more detail, flies were reared on either 0.25% sucrose or 2%, 1.35 g/L, 3 g/L WMP, or 3 g/L WMP with 5 × 108 100 μl cell number of LAL with 0.25% sucrose supplemented Tomato Juice medium was then collected under gaseous CO2 every 24 hours until a minimum of 100 adult males of each genotype were obtained. Standard control is used with 1 mg/L curcumin (C1386, Sigma-Aldrich Co. USA). Live flies were counted and their food changed on days 1, 3 and 5 of a 7 day cycle. In order to assess the efficacy, Lactococcus lactis subsp lactis (LAL, Korean Collection Culture Center, Korea) was dissolved in PBS and the solution was added to fly food to give a final concentration of 5% v/v. Survival curves were plotted using the Kaplan-Meier estimator. The statistical significance was calculated using the log rank test within the SPSS 11.0 statistical package. The null hypothesis in all of the longevity assays was that the presence of the GMR-Gal4 made no difference to the longevity of the flies expressing the Aβ1-42 transgene.
Selected flies were then transferred to upright standard plastic shell vials containing the above food medium they were initially exposed to. Each group was maintained at 25˚C and kept in non-crowded conditions (1 to 20 individuals per vial). Flies were scored for viability every 2 days and transferred to fresh medium without anaesthesia according to established protocol [17] .
2.3. Behavioural Assays
For the determine of climbing speed, groups of ten 3-day-old females were driven into 18-cm-long vials and incubated for 1 h at room temperature for environmental adaptation. After tapping the flies completely down to the bottom, we marked their climbing time at the 15-cm finish line when more than five flies had arrived. Five trials were performed for each group and was repeated with four different groups (n = 50). The average climbing time (±S.D) was calculated for each genotype. Flight assay was performed as previously described [18] with 3-day-old males (n > 100).
2.4. Immunocytochemistry Analysis
Aβ/GFP immunocytochemistry was used as a pacemaker in the whole body of flies in constant fluorescence image for 7 days (n = 20). Identification of specific GFP-positive expressing flies was accomplished by fluorescencing behavior for the membrane-targeted protein. The fluorescence microscopic examinations were carried out with a Nikon microscope with appropriate filter combinations and the images were recorded with a Nikon DXM 1200 digital camera. The different objectives used for microscopy were 5× (0.3NA, Plan Fluor). All the images were assembled using Adobe Photoshop 7.0. RU indicate relative fluorescence arbitrary unit. Scar bars; 100 μm.
2.5. Statistics
Data were analyzed by 2-way analysis of variance (ANOVA) procedures (diet and DSS treatment) using SPSS (IBM, Armonk, NY) to evaluate the effects of diets. Results are presented as means ± SD. Values not sharing common superscript are significantly different at *p < 0.05, **p < 0.01 and #p < 0.001
3. Results
3.1. Increased Concentration of WMP and LAL Protect against Aβ42-Induced Initially Mortality
Here we study a reduced lifespan in flies when Aβ42 expression is enhanced in the brain neurons (Figure 1). The median survival time of Aβ42-expressing flies was reduced by 34% compared to wild type when both groups were fed a control diet. A diet rich in WMP or LAL partially rescued the reduced lifespan by increasing neuronal amounts of Aβ42 in D. melanogaster (Figure 1). Aβ42-expressing flies fed a diet containing 1.35 g/L WMP (73 days), 3 g/L WMP (85 days), 5 × 108/100 μl LAL (78 days), 3 g/L WMP and 5 × 108/100 μl LAL (90 days), and 1 mg/ml curcumin (92 days) had approximately 13 to 30 days longer median lifespan than those fed a control diet (n = 100 flies). Therefore, a concentration of dual 3 g/L WMP and 5 × 108 100 μl LAL dose was significantly observed the survival ratio with Aβ42-expressing flies. The median survival values for each group are found in Table 1.

Table 1. 50% Median survival times of transgenic Drosophila melanogaster cultivated on either a control or WMP and LAL-supplemented diet.
*p < 0.05, flies supplemented with LAL have suspension levels comparable to control; **Represents p < 0.01. Genotypes were UAS-Aβ/GMR-Gal4 (control), and UAS-Aβ/GMR-Gal4 with WMP and LAL supplementation. #Represents p < 0.001. Genotypes were UAS-Aβ/GMR-Gal4 (control) and UAS-Aβ/ GMR-Gal4 with curcumin supplementation.
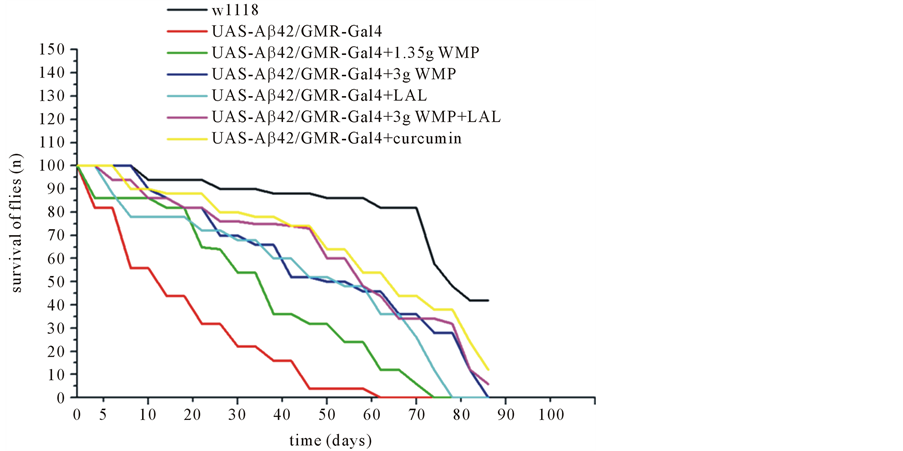
Figure 1. WMP and LAL partially rescues Drosophila melanogaster against Aβ42-induced initially mortality. Directed expression of Aβ42 in lifespan loss in Drosophila fed a standard diet, as compared to a wild type (94 days) (n = 100 flies). Flies fed diets containing 3 g/L WMP and 5 × 108 LAL (90 days) were partially protected against the Aβ42-induced mortality (62 days) (p < 0.01), whereas 1.35 g/L (73 days), 3 g/L WMP (85 days) or 5 × 108/100 μl LAL (78 days) had slightly significant effect. *p < 0.05, flies supplemented with LAL have suspension levels comparable to control. Genotypes are w1118; UAS-Aβ42/GMR-Gal4 (control) and UAS-Aβ42/GMR-Gal4 with WMP or/and LAL supplementation. Errors bars indicate standard error of the mean. p-values were counted by the log-rank (Mantel-Cox) test and multiple comparisons were corrected for using Bonferroni method.
3.2. Dual WMP and LAL Supplementation Ameliorates Aβ42-Induced Developmental Defects in the Drosophila Body
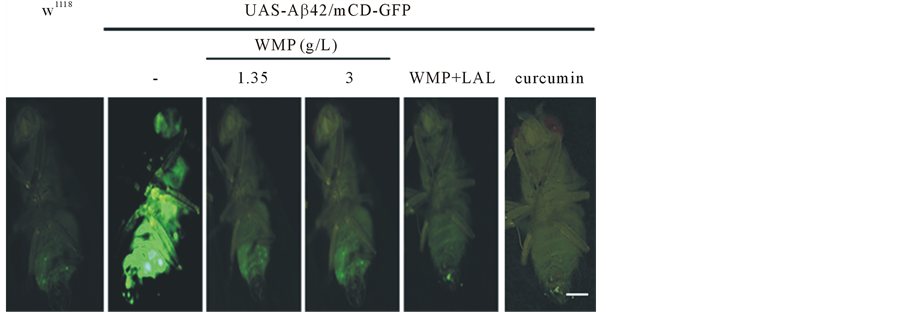
An internal body fluorescence signals occurs when mCD-GFP is used to drive expression of Aβ42 in the developing Drosophila body (Figure 2(a), Figure 2(b)). Also, we tested the neuroprotective effect of WMP and LAL supplementation using Aβ42-expressing Drosophila AD models. When Aβ42 was expressed in developing eyes, strong eye degeneration was observed, compared to the w1118 flies, and imaginal disc images of Aβ42-ex- pressing flies significantly showed a more than approximately eighty percentage reduction in w1118 flies; however, this was not the case for the eyes of control flies. Also, supporting these neurological phenotypes, ectopic expression of Aβ42 strongly induced apoptosis in the larval brain, and the apoptosis was significantly reduced by feed feeding of WMP or both WMP and LAL(data not shown). WMP or both WMP and LAL supplementation significantly reduces mean Aβ42-induced defects to control levels (Figure 2(b)). In Aβ42-expressing flies, the mean expression was ameliorated from those fed a control diet to 52%, 76%, 83% and 92% for flies fed 1.35 g/L WMP, 3 g/L WMP, 3 g/L WMP and 5 × 108/100 μl LAL and 1 mg/ml curcumin supplementation, respectively (n = 20 flies). This provides another example of 3 g/L WMP or 5 × 108/100 μl LAL-induced protection in a Drosophila tissue that is rich in body.
3.3. WMP and LAL Mitigates Aβ42-Induced Flight Impairments in the Drosophila Wing
To confirm further mitochondria impairment in the muscles, we analyzed quality of mitochondria abundance in the mutants by flight ability. As expected, they showed complete defects in flight ability and these phenotypes were significantly rescued by dual WMP and LAL supplementation. Compared to the w1118 flies, Aβ42-ex- pressing flies impaired more than approximately ninety-eight percentage in w1118 flies, and this was significantly rescued by WMP or LAL supplementation (Figure 3) (p < 0.05) (n = 50). Because substantial result represented that anti-apoptotic Bcl-2 families are involved in the protection of mitochondrial integrity and function [19] - [21] . Also, some reports found that aging enhances indirect flight muscle impairment and relation to mitochondrial cytochrome c and wing muscle [22] - [26] . Collectively, these results demonstrate that Aβ42 expression induces indirect flight muscle degeneration and mitochondria impairment. Overall, these results strongly suggest that mitochondrial dysfunction is the main cause of these aberrant phenotypes of Aβ42 expression.
 (a)
(a)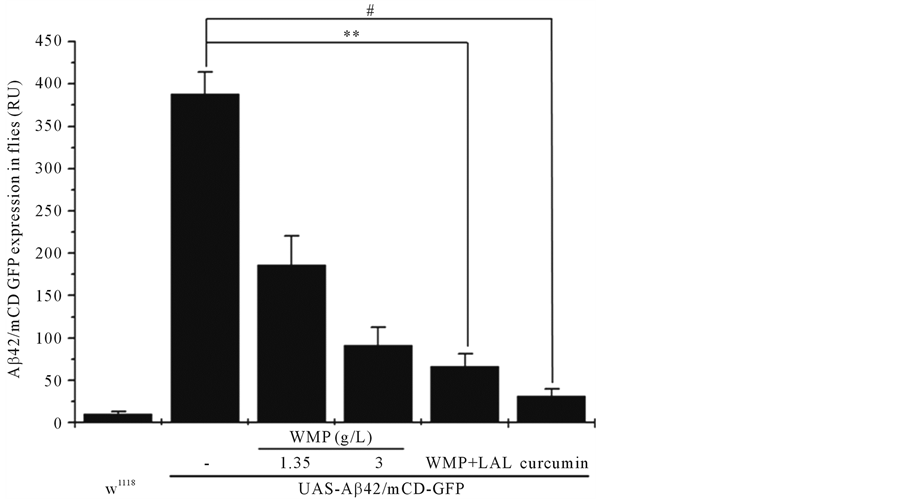 (b)
(b)
Figure 2. Both WMP and LAL supplementation counteracts Aβ42-induced the pacemaker fluorescence signal of the Drosophila whole body. Over expression of Aβ42 during initially whole body observes a significant internal GFP signal. Flies supplemented with WMP and LAL have suspension levels comparable to control. Stereo-fluorescence microscope images (a), and histograms (b). The whole body of wild-type flies is no observed of a fluorescence signal Scale Bars: 100 μm. Error Bars indicate mean ± S.D. N = 20 flies for each group without or with x WMP and LAL supplementation. **Represents p < 0.01. Genotypes were UAS-Aβ42/GMR-Gal4 (control), and UAS-Aβ42/ GMR-Gal4 with WMP and LAL supplementation. #Represents p < 0.001. Genotypes were UAS-Aβ42/GMR-Gal4 (control), and UAS-Aβ42/ GMR-Gal4 with curcumin supplementation.

Figure 3. Comparison of flight ability between w1118 and Aβ42 flies with WMP, LAL or WMP and LAL supplementation. Error Bars indicate mean ± S.D. N = 50 flies for each group without or with x WMP and LAL supplementation. **Represents p < 0.01. Genotypes were UAS-Aβ42/GMR-Gal4 (control), and UAS-Aβ42/GMR-Gal4 with WMP and LAL supplementation. #Represents p < 0.001. Genotypes were UAS-Aβ42/GMR-Gal4 (control), and UAS-Aβ42/GMR-Gal4 with curcumin supplementation.
4. Discussion
Recent reports evidence has suggested that a diet rich in oriental medicine may help delay the Aβ-related degeneration of fly [16] [27] . In other animal study, oriental medicine supplementation improved memory function in older mouse with early memory loss [28] . Moreover, it has been demonstrated that short-term LMK02 (oriental medicine randomly combination agent, LMK02-Jangwonhwan included with red Panax ginseng C.A. Meyer and white Poriacocos [Schw.] Wolf, respectively, by following the guideline posted by the Korean FDA, 2007) supplementation increased calbindin-mediated protection against inflammation in aged mice hippocampal neuronal cells [29] . Also, Probiotic Dahi with Lactobacillus acidophilus and Bifidobacterium bifidum supplementation alleviates age-oxidative stress and improves expression of biomarkers of aged mice [30] [31] . Similar reports have been represented for several other foods high in dietary antioxidants, including extracts of plant polyphenols [32] - [35] , green tea catechins [36] , black tea [37] and ayurvedic herbs [38] . These reports suggest that the protective effects of oriental medicine or probiotic Lactobacillus sp. are not confirmed to Drosophila melanogaster and may extend to mammals. Here we adverts the first demonstration of the neuroprotective effects of dual WMP and LAL in a Drosophila model of Aβ42-AD. Previous findings have demonstrated that Oriental medicine improved both life span and fly eye in a similar Drosophila model of AD [16] [27] . Taken together, these results epitomize the value of using of using Drosophila to study AD aetiology. Though the literature is relatively new, studies in this versatile organism have helped develop interest in the potential protective effects of dietary antioxidants in medical research. Future studies could aim toward unraveling the interaction of dietary antioxidants, probiotic and the activity of mitochondria biogenesis, such as the ROS and enzymatic antioxidant pathways.
Meanwhile, in flight, the superior temporal accuracy of ab1C neurons, olfactory receptor neurons (ORNs) on the antennae may also improve the spatio-temporal resolution of internal representations of stimulus distribution by “sharpening” the borders of odour filaments in time. The fact that CO2-sensitive neurons project only ipsilaterally to their glomeruli, whereas all other ORN classes have both ipsilateral and contralateral projections [39] , suggests that CO2 stimuli are encoded in a different way in the brain. Whereas the identity of odorants in a mixture may have a positive hedonic value, the addition of CO2 might improve the precision with which these odours can be tracked [39] . Even though Drosophila does not require pulsed stimulation for upwind flight as some moth species do [40] , CO2 at low concentrations may enhance attraction by improving navigation through turbulent odour plumes. These neurons differ from most ORNs because their response is mediated by two members of the gustatory receptor family (Gr21a and Gr63b) [41] whereas most other ORNs express a member of the odorant receptor (Or) family co-expressed with the Orco receptor [42] .
The free radical/oxidative stress theory of ageing originated in the 1950’s and suggests that an organism ages due, in part, to the accumulation of free radical-induced damage to cellular macromolecules. Previous reports have shown that probiotic microbiota supplementation can extend lifespan in a Caenorhabditis elegans [32] and Drosophila melanogaster [31] . Our results are novel as we have shown that WMP and LAL supplementation can extend lifespan and flight ability in a Drosophila model of a neurodegenerative disease. The additional antioxidants provided by WMP and LAL supplementation may alleviate some of the excess ROS generated during the progression of AD-like cell death resulting in less cellular injury and a longer median survival time in affected flies.
5. Conclusions
The Drosophila wing consists of multiple subunits, muscle composed of several neurons and wing muscle cells. Directed expression of Aβ42 during early developmental results is premature degeneration of the muscle with abnormal development of external morphology [4] . Our results suggest that a diet containing WMP is originally manufactured by Milne Fruit Products (Prosser, WA) [16] and LAL protects neurons in wing muscle against Aβ42-dependent developmental defects. Both the morphology of atypical wings was improved in flies fed a WMP and LAL-supplemental diet. Aβ42 is a point of major interest in AD pathogenesis and the protective effects of WMP and LAL on Aβ42-induced injury in the Drosophila eye may be due in part to enhanced activity of the mitochondria biogenesis.
In point of fact, WMP and LAL supplement rescue Aβ42-induced flies under conditions of increased oxidative stress and impairment of flight ability, and may be contributed to protect against from impairment of ab1C neurons, olfactory receptor neurons (ORNs) in fly’s brain or wing muscles. Although dietary antioxidants likely provide an invaluable secondary support to cells suffering oxidative stress, it appears their defective effects are dependent on extrinsic/intrinsic antioxidant protection systems. Our finding may be referred to a fortified overall antioxidant defense mechanism in Aβ42-expressing flies. WMP and LAL can increase the expression of enteric probiotic defense molecules in D. melanogaster. Additionally, WMP and LAL extend lifespan and partially protect Aβ42-induced flies under conditions of increased oxidative stress and impairment of flight ability.
Cite this paper
Dong Gyun Ko,Young Bum Eun,Jong Uk Na,Sang-Tae Kim, (2016) Synergistic Effect of Watermelon Powder and Lactococcus lactis subsp lactis Supplemented Diet Partially Ameliorates Aβ42-Dependent Lifespan Shorten and Flight Impairment in Transgenic Drosophila. American Journal of Plant Sciences,07,991-1000. doi: 10.4236/ajps.2016.76094
References
- 1. Abramov, A.Y., Canevari, L. and Duchen, M.R. (2004) Beta-Amyloid Peptides Induce Mitochondrial Dysfunction and Oxidative Stress in Astrocytes and Death of Neurons through Activation of NADPH Oxidase. The Journal of Neuroscience, 24, 565-575.
http://dx.doi.org/10.1523/JNEUROSCI.4042-03.2004 - 2. Kienlen-Campard, P., Miolet, S., Tasiaux, B. and Octave, J.-N. (2002) Intracellular Amyloid-Beta 1-42, but Not Extracellular Soluble Amyloid-Beta Peptides, Induces Neuronal Apoptosis. The Journal of Biological Chemistry, 277, 5666-15670.
http://dx.doi.org/10.1074/jbc.M200887200 - 3. Wei, W., Norton, D.D., Wang, X. and Kusiak, J.W. (2002) Abeta 17-42 in Alzheimer’s Disease Activates JNK and Caspase-8 Leading to Neuronal Apoptosis. Brain, 125, 2036-2043.
http://dx.doi.org/10.1093/brain/awf205 - 4. Cao, W., Song, H.J., Gangi, T., Kelkar, A., Antani, I., Garza, D. and Konsolaki, M. (2008) Identification of Novel Genes That Modify Phenotypes Induced by Alzheimer’s β-Amyloid Overexpression in Drosophila. Genetics, 178, 1457-1471.
http://dx.doi.org/10.1534/genetics.107.078394 - 5. Schapira, A.H. and Jenner, P. (2011) Etiology and Pathogenesis of Parkinson’s Disease. Movement Disorders, 26, 1049-1055.
http://dx.doi.org/10.1002/mds.23732 - 6. Byers, B., Cord, B., Nguyen, H.N., Schüle, B., Fenno, L., Lee, P.C., Deisseroth, K., Langston, J.W., Pera, R.R. and Palmer, T.D. (2011) SNCA Triplication Parkinson’s Patient’s iPSC-Derived DA Neurons Accumulate α-Synuclein and Are Susceptible to Oxidative Stress. PLoS ONE, 6, e26159.
http://dx.doi.org/10.1371/journal.pone.0026159 - 7. Huang, X., Atwood, C.S., Hartshorn, M.A., Multhaup, G., Goldstein, L.E., Scarpa, R.C., Cuajungco, M.P., Gray, D.N., Lim, J., Moir, R.D., Tanzi, R.E. and Bush, A.I. (1999) The A Beta Peptide of Alzheimer’s Disease Directly Produces Hydrogen Peroxide through Metal Ion Reduction. Biochemistry, 38, 7609-76167.
http://dx.doi.org/10.1021/bi990438f - 8. Bush, A.I. (2003) The Metallobiology of Alzheimer’s Disease. Trends in Neurosciences, 26, 207-214.
http://dx.doi.org/10.1016/S0166-2236(03)00067-5 - 9. Rimando, A.M. and Perkins-Veazie, P.M. (2005) Determination of Citrulline in Watermelon Rind. Journal of Chromatography A, 1078, 196-200.
http://dx.doi.org/10.1016/j.chroma.2005.05.009 - 10. Altas, P.M., Kizil, G., Kizil, M., Ketani, A. and Haris, P.I. (2011) Protective Effect of Diyarbakir Watermelon Juice on Carbon Tetrachloride-Induced Toxicity in Rats. Food and Chemical Toxicology, 49, 2433-2438.
http://dx.doi.org/10.1016/j.fct.2011.06.064 - 11. Naz, A., Butt, M.S., Sultan, M.T., Qayyum, M.N.N. and Niaz, R.S. (2014) Watermelon Lycopene and Allied Health Claims. EXCLI Journal, 13, 650-666.
- 12. Poduri, A., Rateri, D., Saha, S., Saha, S. and Daugherty, A. (2012) Citrullus lanatus “Sentinel” (Watermelon) Extract Reduces Atherosclerosis in LDL Receptor-Deficient Mice. The Journal of Nutritional Biochemistry, 24, 882-886.
- 13. Linder, J.E. and Promislow, D.E. (2009) Cross-Generational Fitness Effects of Infection in Drosophila melanogaster. Fly, 3, 143-150.
http://dx.doi.org/10.4161/fly.8051 - 14. Crowther, D.C., Kinghorn, K.J., Miranda, E., Page, R., Curry, J.A., Duthie, F.A., Gubb, D.C. and Lomas, D.A. (2005) Intraneuronal Aβ, Non-Amyloid Aggregates and Neurodegeneration in a Drosophila Model of Alzheimer’s Disease. Neuroscience, 132, 123-135.
http://dx.doi.org/10.1016/j.neuroscience.2004.12.025 - 15. Nilsberth, C., Westlind-Danielsson, A., Eckman, C.B., Condron, M.M., Axelman, K., Forsell, C., Stenh, C., Luthman, J., Teplow, D.B., Younkin, S.G., Naslund, J. and Lannfelt, L. (2001) The “Arctic” APP Mutation (E693G) Causes Alzheimer’s Disease by Enhanced Aβ Protofibril Formation. Nature Neuroscience, 4, 887-893.
http://dx.doi.org/10.1038/nn0901-887 - 16. Wu, G., Collins, J.K., Perkins-Veazie, P., Siddiq, M., Dolan, K.D., Kelly, K.A., Heaps, C.L. and Meininger, C.J. (2007) Dietary Supplementation with Watermelon Pomace Juice Enhances Arginine Availability and Ameliorates the Metabolic Syndrome in Zucker Diabetic Fatty Rats. Journal of Nutrition, 137, 2680-2685.
- 17. Abramoff, M.D., Magalhaes, P.J. and Ram, S.J. (2004) Image Processing with ImageJ. Biophotonics International, 11, 36-42.
- 18. Pesah, Y., Pham, T., Burgess, H., Middlebrooks, B., Verstreken, P., Zhou, Y., Harding, M., Bellen, H. and Mardon, G. (2004) Drosophila parkin Mutants Have Decreased Mass and Cell Size and Increased Sensitivity to Oxygen Radical Stress. Development, 131, 2183-2194.
http://dx.doi.org/10.1242/dev.01095 - 19. Leulier, F., Ribeiro, P.S., Palmer, E., Tenev, T., Takahashi, K., et al. (2006) Systematic in Vivo RNAi Analysis of Putative Components of the Drosophila Cell Death Machinery. Cell Death & Differentiation, 13, 1663-1674.
http://dx.doi.org/10.1038/sj.cdd.4401868 - 20. Vander Heiden, M.G., Chandel, N.S., Williamson, E.K., Schumacker, P.T. and Thompson, C.B. (1997) Bcl-xL Regulates the Membrane Potential and Volume Homeostasis of Mitochondria. Cell, 91, 627-637.
http://dx.doi.org/10.1016/S0092-8674(00)80450-X - 21. Vander Heiden, M.G. and Thompson, C.B. (1999) Bcl-2 Proteins: Regulators of Apoptosis or of Mitochondrial Homeostasis? Nature Cell Biology, 1, E209-E216.
http://dx.doi.org/10.1038/70237 - 22. Godenschwege, T., Forde, R., Davis, C.P., Paul, A., Beckwith, K. and Duttaroy, A. (2009) Mitochondrial Superoxide Radicals Differentially Affect Muscle Activity and Neural Function. Genetics, 183, 175-184.
http://dx.doi.org/10.1534/genetics.109.103515 - 23. Miller, M.S., Lekkas, P., Braddock, J.M., Farman, G.P., Ballif, B.A., Irving, T.C., Maughan, D.W. and Vigoreaux, J.O. (2008) Aging Enhances Indirect Flight Muscle Fiber Performance yet Decreases Flight Ability in Drosophila. Biophysical Journal, 95, 2391-2401.
http://dx.doi.org/10.1529/biophysj.108.130005 - 24. Yan, L.J. and Sohal, R.S. (1998) Mitochondrial Adenine Nucleotide Translocase Is Modified Oxidatively during Aging. Proceedings of the National Academy of Sciences of the United States of America, 95, 12896-12901.
- 25. O’Brien, S.J. and Shimada, Y. (1974) The α-Glycerophosphate Cycle in Drosophila Melanogaster. IV. Metabolic, Ultrastructural, and Adaptive Consequences of αGpdh-l “Null” Mutations. The Journal of Cell Biology, 63, 864-882.
- 26. Levenbook, L. and Williams, C.M. (1956) Mitochondria in the Flight Muscles of Insects. III. Mitochondrial Cytochrome c in Relation to the Aging and Wing Beat Frequency of Flies. The Journal of General Physiology, 39, 497-512.
- 27. Hong, Y.K., Park, S.H., Lee, S., Hwang, S., Lee, M.J., Kim, D., Lee, J.H., Han, S.Y., Kim, S.T., Kim, Y.K., Jeon, S., Koo, B.S. and Cho, K.S. (2011) Neuroprotective Effect of SuHeXiang Wan in Drosophila Models of Alzheimer’s Disease. Journal of Ethnopharmacology, 134, 1028-1032.
http://dx.doi.org/10.1016/j.jep.2011.02.012 - 28. Cai, Z.L., Wang, C.Y., Jiang, Z.J., Li, H.H., Liu, W.X., Gong, L.W., Xiao, P. and Li, C.H. (2013) Effects of Cordycepin on Y-Maze Learning Task in Mice. European Journal of Pharmacology, 714, 249-253.
http://dx.doi.org/10.1016/j.ejphar.2013.05.049 - 29. Seo, J.S., Yun, J.H., Baek, I.S., Leem, Y.H., Kang, H.W., Cho, H.K., Lyu, Y.S., Son, H.J. and Han, P.L. (2010) Oriental Medicine Jangwonhwan Reduces Aβ(1-42) Level and β-Amyloid Deposition in the Brain of Tg-APPswe/PS1dE9 Mouse Model of Alzheimer Disease. Journal of Ethnopharmacology, 128, 206-212.
http://dx.doi.org/10.1016/j.jep.2010.01.014 - 30. Kaushal, D. and Kansal, V.K. (2012) Probiotic Dahi Containing Lactobacillus acidophilus and Bifidobacterium bifidum Alleviates Age-Inflicted Oxidative Stress and Improves Expression of Biomarkers of Ageing in Mice. Molecular Biology Reports, 39, 1791-1799.
http://dx.doi.org/10.1007/s11033-011-0920-1 - 31. Lee, K.A. and Lee, W.J. (2014) Drosophila as a Model for Intestinal Dysbiosis and Chronic Inflammatory Diseases. Developmental & Comparative Immunology, 42, 102-110.
http://dx.doi.org/10.1016/j.dci.2013.05.005 - 32. Grompone, G., Martorell, P., Llopis, S., González, N., Genovés, S., Mulet, A.P., Fernández-Calero, T., Tiscornia, I., Bollati-Fogolín, M., Chambaud, I., Foligné, B., Montserrat, A. and Ramón, D. (2012) Anti-Inflammatory Lactobacillus rhamnosus CNCM I-3690 Strain Protects against Oxidative Stress and Increases Lifespan in Caenorhabditis elegans. PLoS ONE, 12, e52493.
http://dx.doi.org/10.1371/journal.pone.0052493 - 33. Schriner, S.E., Katoozi, N.S., Pham, K.Q., Gazarian, M., Zarban, A. and Jafari, M. (2012) Extension of Drosophila Lifespan by Rosa damascena Associated with an Increased Sensitivity to Heat. Biogerontology, 13, 105-117.
http://dx.doi.org/10.1007/s10522-011-9357-0v - 34. Peng, C., Chan, H.Y., Huang, Y., Yu, H. and Chen, Z.Y. (2011) Apple Polyphenols Extend the Mean Lifespan of Drosophila Melanogaster. Journal of Agricultural and Food Chemistry, 59, 2097-2106.
http://dx.doi.org/10.1021/jf1046267 - 35. Long, J.G., Gao, H.X., Sun, L.J., Liu, J.K. and Zhao, W.X. (2009) Grape Extract Protects Mitochondria from Oxidative Damage and Improves Locomotor Dysfunction and Extends Lifespan in a Drosophila Parkinson’s Disease Model. Rejuvenation Research, 12, 321-331.
http://dx.doi.org/10.1089/rej.2009.0877 - 36. Li, Y.M., Chan, H.Y., Huang, Y. and Chen, Z.Y. (2007) Green Tea Catechins Upregulate Superoxide Dismutase and Catalase in Fruit Flies. Molecular Nutrition & Food Research, 51, 546-554.
http://dx.doi.org/10.1002/mnfr.200600238 - 37. Peng, C., Chan, H.Y., Li, Y.M., Huang, Y. and Chen, Z.Y. (2009) Black Tea Theaflavins Extend the Lifespan of Fruit Flies. Experimental Gerontology, 44, 773-783.
http://dx.doi.org/10.1016/j.exger.2009.09.004 - 38. Jansen, R.L., Brogan, B., Whitworth, A.J. and Okello, E.J. (2014) Effects of Five Ayurvedic Herbs on Locomotor Behaviour in a Drosophila melanogaster Parkinson’s Disease Model. Phytotherapy Research, 28, 1789-1795.
http://dx.doi.org/10.1002/ptr.5199 - 39. Jones, W.D., Cayirlioglu, P., Kadow, I.G. and Vosshall, L.B. (2006) Two Chemosensory Receptors Together Mediate Carbon Dioxide Detection in Drosophila. Nature, 445, 86-90.
http://dx.doi.org/10.1038/nature05466 - 40. Kwon, J.Y., Dahanukar, A., Weiss, L.A. and Carlson, J.R. (2007) The Molecular Basis of CO2 Reception in Drosophila. Proceedings of the National Academy of Sciences of the United States of America, 104, 3574-3578.
- 41. Scott, K., Brady Jr., R., Cravchik, A., Morozov, P., Rzhetsky, A., Zuker, C. and Axel, R. (2001) A Chemosensory Gene Family Encoding Candidate Gustatory and Olfactory Receptors in Drosophila. Cell, 104, 661-673.
http://dx.doi.org/10.1016/S0092-8674(01)00263-X - 42. Budick, S.A. and Dickinson, M.H. (2006) Free-Flight Responses of Drosophila melanogaster to Attractive Odors. Journal of Experimental Biology, 209, 3001-3017.
http://dx.doi.org/10.1242/jeb.02305
NOTES

*Corresponding author.