 Vol.2, No.4, 141-154 (2012) Stem Cell Discovery http://dx.doi.org/10.4236/scd.2012.24019 Differentiation of human epidermis-derived mesenchymal stem cell-like pluripotent cells into neural-like cells in culture and after transplantation Min Zhang, Bing Huang*, Kaijing Li, Zhenghua Chen, Jian Ge, Weihua Li, Jianfa Huang, Ting Luo, Shaochun Lin, Jie Yu, Wencong Wang, Liping Lin State Key Laboratory of Ophthalmology, Zhongshan Ophthalmic Center, Sun Yat-sen University, Guangzhou, China; *Corresponding Author: huangbing2000@hotmail.com Received 11 August 2012; revised 12 September 2012; accepted 9 October 2012 ABSTRACT Skin is the largest organ of the human body and a possible source of stem cells for research and cell-based therapy. We have isolated a popula- tion of mesenchymal stem cell-like pluripotent cells from human epidermis, termed human (h) EMSCPCs. This preliminary study tested if these hEMSCPCs can be induced to differentiate into neural-like cells. Human EMSCPCs were first cultured for four to seven days in a serum-free neural stem cell (NSC) medium for pre-induction. During pre-induction, hEMSCPCs coalesced into dense spheres that resembled neural rosettes. In the presence of a conditioned differentiation medium, pre-induced cells took on the morpho- logical characteristics of neural cells, including slender projections with inflated or claw-like ends that contacted the soma or projections of other cells as revealed by confocal microscopy. Moreover, these differentiating cells expressed the neural-specific markers β-III tubulin, MAP2, GFAP, and synapsin I as evidenced by immu- nocytochemistry. Both pre-induced hEMSCPCs and uninduced hEMSCPCs were labeled with CM-DiI and transplanted into the vitreous cavi- ties of nude mice. Transplanted cells were ex- amined four weeks later in frozen eyeball sec- tions by immunofluorescence staining, which demonstrated superior retinal migration and neural differentiation of pre-induced cells. Our study is the first to demonstrate tha t hEMSCPC s possess the capacity to differentiate into neu- ral-like cells, suggesting potential uses for the treatment of retinal di seases such as age-r elated macular degeneration. Keywords: Human Epidermis; Pluripotent Cells; Differentiation; Neural Cells; Cell Therapy 1. INTRODUCTION Neurological degenerative diseases like Alzheimer’s disease, Parkinson’s disease, and age-related macular degeneration are a group of chronic, diverse and pro- gressive disorders. Studies have shown heredity, oxida- tive stress, neurotrophic factor deficiency, dysbolism and other unknown factors could cause a main pathological change of special neurons degeneration and loss, fol- lowed by demyelination of nerve fibers [1]. These patho- physiology leads to a decreased activity in the pathway of neural conduction, and results in disturbance of me- mory, learning, moving and other activities, which causes severe public health burden, particularly in an aging po- pulation. Current treatments for these diseases include neuroprotective agents, surgery, physical stimulation, gene therapy, and cell replacement [2-4]. However, pharma- cologic, surgical, and physical therapies cannot cure these diseases. Recent reports have shown that gene and cell-replacement therapies are promising alternatives for treating or even curing neurological disorders [5-7]. How- ever, the complexity of the human genome and proteome limits the therapeutic effects of single gene therapy. In- deed, the results of clinical trials testing single gene therapies for Parkinson’s disease were less than ideal [8-9]. Theoretically, cell-replacement therapy can cure neu- rodegenerative diseases by replacing lost cells and re- constructing tissues, leading to functional recovery. Mul- tipotent stem cells are widely used for research on cell- replacement therapy and can be derived from both the embryo (embryonic stem cells, ESCs) and adult tissue (adult stem cells, ASCs). The strong plasticity of ASCs enables directional differentiation into multiple cell types. Numerous studies have revealed that ESCs, neural stem cells (NSCs), bone marrow stem cells (BMSCs), and Copyright © 2012 SciRes. OPEN ACCE SS  M. Zhang et al. / Stem Cell Discovery 2 (2012) 141-154 142 precursor cells derived from peripheral blood, umbilical cord blood, and fat tissue can differentiate into neural cells [10-17]. However, there are ethical issues sur- rounding the harvesting of ESCs; moreover, these cells are potentially oncogenic [18-19]. Neural stem cells exist in several adult human tissues but are difficult to isolate [20]. The process of isolating BMSCs is invasive and painful, and the quantities obtained are generally not suf- ficient for therapeutic applications [21]. Moreover, more accessible peripheral blood and umbilical cord blood contain relatively low numbers of precursor cells, and it is still disputed whether these precursor cells can inte- grate with the host tissue and differentiate into the ap- propriate cell types after transplantation [22-24]. Al- though fat tissue is easily extracted, techniques for the isolation and purification of precursor cells from adipose tissue are still not fully developed [25]. Skin is the largest human organ and its cells are easily harvested. Several groups have isolated pluripotent cells from mammalian skin that can differentiate into neural cells [26-32]. After transplantation into animal models, these skin-derived pluripotent cells were able to promote nerve regeneration and functional improvement after injury [33-36]. It is known that epidermal stem cell-like pluripotent cells are present in the epidermal-basal layer and function in the repair and regeneration of the epi- dermis [37-38]. We have isolated a population of me- senchymal stem cell-like pluripotent cells from mixed cultures of human epidermal cells that we refer to as human epidermis-derived mesenchymal stem cell-like pluripotent cells (hEMSCPCs) (national patent number: 201010282388.0) [39]. In this exploratory study, we exa- mined whether hEMSCPCs have the capacity to dif- ferentiate into neural-like cells. Our data revealed that hEMSCPCs can be induced to differentiate into cells with neural cell characteristics in vitro and express some neural cell-specific markers in vivo when transplanted into the mouse eye, suggesting the hEMSCPCs may be used for autologous cell-based therapies to treat neuro- logical disorders. 2. MATERIALS AND METHODS 2.1. Isolation of hEMSCPCs and Cultured in Growth Medium The hEMSCPCs were isolated from foreskin tissue obtained from circumcision surgery. Tissue donors were healthy as defined by normal blood and urine test results, normal liver and lung function, no history of genetic disease, and the absence of current infectious disease. Written informed consent was provided by the partici- pants. The study was approved by the Medical Ethics Committee of Zhongshan Ophthalmic Center, Sun Yat- sen University (No. 2008-30). Briefly, foreskin tissue was rinsed in phosphate buff- ered saline (PBS) containing gentamycin (1000 U/ml) for subsequent treatment, the tissue was cut into pieces of 3 mm × 3 mm in size using scalpes and transferred into a sterilized 15 ml centrifugation tube. Then DispaseⅡ (2 U/ml; GIBCO, USA) was added into the tube, incu- bated at 6˚C - 8˚C for 15 hours and then 37˚C for 1 hour to remove the dermis. The epidermis were transferred into a new sterilized 15 ml centrifugation tube, washed with PBS for 5 times and crushed. Then suspended with PBS containing 0.25% trypsin and gently pippetted, in- cubated at 37˚C for 30 min. Then washed with PBS twice more and centrifugated at 1200 rpm for 5 min, discarded the supernatant. Cell precipitation was sus- pended in growth medium consisting of 80% DMEM (GIBCO, USA), 18% fetal bovine serum (FBS) (Si Jiqing Ltd., China), 10 ng/ml basic fibroblast growth factor (bFGF) (PERPO-TECH, USA), 2 ng/ml stem cell factor (SCF) (PERPO-TECH, USA), and 1% MEM non- essential amino acids (NEAA) (100 × solution, GIBCO, USA), and plated in T-25 cell culture flask, incubated at 37˚C in a 5% CO2 atmosphere. The flask remained un- moved within 48 hours, then the medium was replaced according to the rate of cell growth, and the un-adherent cells were removed. Ten days later, small hEMSCPCs appeared; three weeks later, they were deplated using 0.25% trypsin-0.02% ethylene diamine tetraacetic acid (EDTA) and passaged at 1:3. They were continuously cultured and passaged over 30 times in vitro [39]. The hEMSCPCs from passages 17 to 19 derived from same biopsy (fore-skin of a 21-year-old male) were used for this study. Cryopreserved hEMSCPCs were resuscitated from liquid nitrogen and suspended in growth medium (men- tioned above). Cell suspensions (8 ml of 1.0 × 104 cells/ ml) were plated in T-25 cell culture flasks and incu- bated at 37˚C in a 5% CO2 atmosphere. The medium was replaced according to the rate of cell growth. When cells reached confluence, the hEMSCPCs were deplated using 0.25% trypsin-0.02% EDTA and passaged at 1:3. Cul- tures were observed and photographed using an in- verted microscope (Leica DMIRB, Germany). 2.2. Pre-Induction Culture of hEMSCPCs in NSC Medium For pre-induction, hEMSCPCs were deplated and suspended in NSC medium consisting of 96% DMEM/ F12 (GIBCO, USA), 2% B27 (GIBCO, USA), 20 ng/ml bFGF (PERPOTECH, USA), 20 ng/ml epidermal growth factor (EGF) (PERPOTECH, USA), 2 ng/ml SCF (PER- POTECH, USA), and 1% MEM NEAA (100 × solution, GIBCO, USA). Cell suspensions (8 ml of 1.0 × 104 cells/ml) were replated in T-25 cell culture flasks and incubated at 37˚C under 5% CO2. The medium was re- Copyright © 2012 SciRes. OPEN ACCE SS  M. Zhang et al. / Stem Cell Discovery 2 (2012) 141-154 143 placed everyday. Cultures were observed and photo- graphed as described above. 2.3. Differentiation of hEMSCPCs in Conditioned Differentiation Medium After 6 days, the NSC medium was replaced with a conditioned differentiation medium consisting of 88% DMEM/F12 (GIBCO, USA), 10% FBS (Si Jiqing Ltd., China), 20 ng/ml bFGF (PERPOTECH, USA), 20 ng/ml EGF (PERPOTECH, USA), and 1% MEM NEAA (100 × solution, GIBCO, USA). Cells were incubated for 1 or 3 weeks depending on the experiment. In addition, hEM- SCPCs cultured in growth medium but not pre-induced in NSC medium were cultured in differentiation medium as a control. Cultures were observed and photographed under an inverted microscope (Leica DMIRB, Germany). The hEMSCPCs pre-induced in the NSC medium grew slender projections during differentiation, so some cul- tures were plated at lower density (1.0 × 103 cells/ml) to aid in morphological observation. 2.4. Subculturing for Immunofluorescence and CM-DiI Staining To detect the expression of cell-specific markers in hEMSCPCs during differentiation, cells were seeded onto cover slips and immunostained (below). Other cul- tures were labeled with CM-DiI to observe cell-cell con- tacts. Seeded cover slips were divided into five groups of 10 slides each. The hEMSCPCs cultured in the growth medium but not pre-induced by NSC medium constituted group A (GM, control). The hEMSCPCs cultured in growth medium and then pre-induced in the NSC me- dium for six days constituted group B (GM + NSC). Hu- man EMSCPCs cultured in growth medium, pre-induced in NSC medium for six days, and then cultured in the conditioned differentiation medium for three weeks were group C (GM + NSC + CM 3 weeks). Human EMSCP- Cs cultured in the growth medium but not pre-induced in the NSC medium for six days before culture in the con- ditioned differentiation medium for three weeks were group D (GM + CM 3 weeks) and served as the control for group C. Group E consisted of hEMSCPCs cultured in growth medium, pre-induced in the NSC medium for six days, and then cultured in the conditioned differentia- tion medium for one week (GM + NSC + CM 1 week). At the beginning, cell densities of group A and group B were adjusted to 1.0 × 104 cells/ml, while those of groups C, D, and E were adjusted to 1.0 × 103 cells/ml for im- proved morphological observation of differentiating cells. Human EMSCPCs suspensions were plated onto sterile 22 × 22 mm2 cover slips in 35 mm culture dishes. Each culture dish contained one hEMSCPCs cover slip. All cultures were incubated at 37˚C under 5% CO2. After the treatments described above, cover slips were collected for staining. Groups A, B, C, and D cells were stained by immunofluorescence and group E cultures were labeled with CM-DiI. 2.5. CM-DiI Labeling of the Differentiating hEMSCPCs Cells were labeled with CM-DiI rather than processed for electron microscopy (EM) because EM fixation and processing/staining tend to cause contraction of pro- cesses [40,41]. Furthermore, EM is laborious and expen- sive. In contrast, CM-DiI is a lipid-soluble biomembrane stain that allows for clear visualization of cell morpho- logy and cell-cell contacts [42]. Ten hEMSCPCs-seeded cover slips of group E (de- fined above) were collected, washed twice in PBS, and stained with 5 μl/ml CM-DiI (Molecular Probes, USA) in 200 μl PBS for 3 min at 37˚C. Stained cover slips were washed twice quickly in PBS and then fixed in 4% para- formaldehyde for 40 min. The nuclei were counterstained by Hoechst (Sigma, USA) for 5 min at room temperature (RT). Slides were then treated by an anti-fade solution (Applygen, China) and imaged under a laser confocal scanning microscope (Zeiss, Germany). Ten different vi- sual fields were observed in each cover slip. 2.6. Immunofluorescence Staining of hEMSCPCs Cultured in Vitro Immunofluorescence staining was used to detect the expression of cell-specific antigens in hEMSCPCs cul- tured in vitro. The hEMSCPCs-seeded cover slips of groups A, B, C, and D were collected and fixed in 4% paraformaldehyde for 15 min, permeabilized in 0.3% Triton-X for 15 min, and then incubated at 37˚C for 40 min in the following primary antibodies: human nestin, MAP2, synapsin I (Abcam, USA), vimentin (ZSGB-BIO, China), β-III tubulin (Millipore, USA), and GFAP (Ep- tomics, USA). Slides were then incubated in secondary antibodies, either Cy3-conjugated goat anti-mouse (Mil- lipore, USA) or FITC-conjugated goat anti-rabbit (South- ern Biotech, USA) for 30 min at RT. The nuclei were stained by Hoechst (Sigma, USA) for 5 min at RT, and then all slides were treated with an anti-fade solution (Applygen, China) and imaged under a laser confocal scanning microscope (Zeiss, Germany). Ten different vi- sual fields were observed in each cover slip. 2.7. Flow Cytometry Flow cytometry was used to quantify the expression of cell-specific markers in GM + NSC 4 days and GM hEMSCPCs groups. Both direct labeling and indirect labeling were used. Antibodies used for direct labeling were specific for human CD73 and its PE-iso-type con- Copyright © 2012 SciRes. OPEN ACCE SS  M. Zhang et al. / Stem Cell Discovery 2 (2012) 141-154 144 trol (BD, USA). Primary antibodies used for indirect labeling were specific for human nestin (Abcam, USA), vimentin (ZSGB-BIO, China), MAP2, and GFAP (Ab- cam, USA). The secondary antibodies were R-PE-con- jugated goat anti-mouse and FITC-conjugated goat anti-rabbit (Southern Biotech, USA). Fix and Perm Cell Permeabilization reagents (Invitrogen, USA) were used for labeling intracellular antigens. Cell suspensions were stained and counted by flow cytometry according to the manufacturer’s directions. Cell suspensions treated with secondary antibodies but not primary antibodies served as iso-type controls for indirect labeling. Cell suspend- sions were tested immediately by flow cytometry (BD FACSAriaTM, USA) using FCS Express V3 software for data analysis. The positive value of iso-type controls was maintained at 0% to 1%. Prior to cell transplantation in vivo (below), the ex- pression levels of immunogenic markers (HLA-I and HLA-DR) in GM + NSC 4 days and GM hEMSCPCs were first detected by direct labeling and flow cytometry. Cell suspensions were treated with antibodies specific for human HLA-I (Invitrogen, USA), HLA-DR, and their FITC-iso-type control (BD, USA). Protocols were in accordance with the manufacturer’s directions. 2.8. Transplantation of hEMSCPCs into the Vitreous Cavities of Nude Mice The retina, an extension of the central nervous system containing a variety of highly differentiated cell types, was chosen to provide the internal microenvironment for hEMSCPCs differentiation. Human EMSCPCs were trans- planted into the vitreous cavities of nude mice, and mi- gration and differentiation were observed after four weeks. The GM + NSC 4 days and GM cultures were termed groups A and B in the transplantation study. Before transplantation, both groups were labeled by CM-DiI (Molecular Probes, USA) according to the manufac- turer’s instructions. In addition, one 500 μl sample from each cell suspension labeled by CM-DiI was analyzed by flow cytometry to test CM-DiI labeling efficiency. Cell suspensions (1 × 107 cells/ml) with high labeling effi- ciency were immediately injected into the eyes of nude mice. Sixteen 6-week-old BLAB/c nude mice of both gen- ders were provided by the Laboratory Animal Center, Sun Yat-sen University (Quality certificate number: 0061839). The mice were cared for in accordance with the Regulations on Administration of Experimental Ani- mals in Guangdong Province, China. They were housed in the specific pathogen-free (SPF) area of the Ophthal- mology Animal Experimental Center, ZhongShan Oph- thalmic Center, Sun Yat-sen University, with a 12 h light- dark cycle (23˚C - 25˚C, humidity 55%). These trans- plantation experiments were approved by the Laboratory Animal Administration and Ethics Committee of Zhong- shan Ophthalmic Center, Sun Yat-sen University (No. 2010-024). The 16 nude mice were randomly divided into three groups (group A, B, and C), each with a 1:1 sex ratio. The right eyes of all group A (n = 6) mice were injected with hEMSCPCs that had been pre-induced in the NSC medium for four days, while the right eyes of group B were injected with hEMSCPCs cultured in the growth medium but not pre-induced by NSC medium. The remaining four mice (group C) served as normal controls. Before transplantation, experimental mice were anes- thetized by intraperitoneal injection of 4.3% chloral hy- drate (0.01 ml per 1 g body weight) obtained from the Ophthalmologic Hospital, Sun Yat-sen University. To- bramycin eye drops were used to disinfect the experi- mental eye, followed by dicaine hydrochloride eye drops (both drugs obtained from the Ophthalmologic Hospital, Sun Yat-sen University) for superficial anesthesia. A 1-ml-injector needle was used to pierce the central cor- nea and drain part of the aqueous humor to lower the intraocular pressure. Under an operating microscopy (Topcon, Japan), a second 1-ml injector containing 10 μl of either cell suspension (pre-induced with NSC medium or untreated) was placed 1 mm outside the corneoscleral junction on the temporal side. The needle penetrated at a 15˚C acute angle relative to the eyeball coronal plane. When the needle reached the vitreous cavity, cell sus- pendsions were injected. The needle was immediately withdrawn at the first signs of eyeball puffing. The ex- perimental eye was then washed with tobramycin eye drops, followed by tobramycin eye ointment. All proce- dures were performed in the SPF area operating room. Appropriate body temperature was maintained during the operation and intraoperative animal care conformed to institutional guidelines. After mice regained conscious- ness, they were sent back to the feeding room. To prevent infection, tobramycin eye drops were applied three times daily for three days after the operation. 2.9. Frozen Eyeball Sectioning and Immunofluorescence Staining Four weeks after cell transplantation, all the mice were anesthetized by intraperitoneal injection of 4.3% chloral hydrate and sacrificed by cervical dislocation. All right eyes were enucleated, frozen in OCT embedding com- pound (Sakura, USA), and stored at –20˚C before cry- ostat sectioning at 6 μm on a freezing microtome (Leica, Germany). Sections were then processed for immunofl- uorescence labeling as described (see Immunofluores- cence staining of hEMSCPCs cultured in vitro). Primary antibodies were specific for human nestin, MAP2, rhodopsin (Abcam, USA), β-III tubulin (Sigma, USA), Copyright © 2012 SciRes. OPEN ACCE SS  M. Zhang et al. / Stem Cell Discovery 2 (2012) 141-154 145 and GFAP (Epitomics, USA). An FITC-conjugated goat anti-rabbit antibody (Southern Biotech, USA) was used for fluorescence tagging. The migration and differentia- tion of the transplanted hEMSCPCs in nude mice were examined by laser confocal scanning microscopy (Zeiss, Germany). Ten different visual fields were observed in each section. 2.10. Statistical Analysis Analysis of variance (ANOVA) with repeated meas- ures was used to compare treatment means. A value of P < 0.05 was considered statistically different. The SPSS v13.0 software package was used for all statistical ana- lyses. 3. RESULTS 3.1. Morphology of hEMSCPCs Cultured in Growth Medium When cultured in growth medium, hEMSCPCs ap- peared as small spindle-shaped cells that adhered to the bottoms of culture flasks in a monolayer arranged in a vortex pattern (Figures 1(A) and (B)). They proliferated rapidly in growth medium; when seeded at 1.0 × 104 cells/ml × 8 ml in T-25 plastic culture flasks, they rea- ched confluence in two days [39]. 3.2. Changes in hEMSCPCs Morphology during Pre-Induction in NSC Medium To direct hEMSCPCs toward the neuronal lineage, we provided a conducive environment using a neural stem cell (NSC) medium. One day after culture in NSC me- dium, the hEMSCPCs somata aggregated to form small dense light-reflective spheres that were fully or partly adherent to the bottoms of the flasks. A few small spin- dle-shaped cells began to stretch out from the edges of the spheres (Figures 1(C) and (D)). After three days in NSC medium, many spindle-shaped cells stretched out from the edges of the spheres and formed strongly light reflective rosette-shaped clusters at the edges (Figures 1(E) and (F)). As days went on, the spindle-shaped cells stretched out further and the rosettes enlarged (Figures 1(G) and (H)). 3.3. Changes in hEMSCPCs Morphology during Culture in Conditioned Differentiation Medium as Revealed by Light Microscopy At day 6, NSC medium was replaced with a condi- tioned differentiation medium. Within one day, most cells began to grow one or more slender projections that resembled neurites (Figures 2(B)- (D)). Three days later, as these projections became longer and continued to extend, contacts were formed between cells (Figures 2(E)-(G)). After one week in conditioned medium, the projections continued to extend and branches emerged at the ends (Figure 2(H)). In contrast, hEMSCPCs cultured in the growth medium but not in NSC medium showed no obvious changes in morphology during incubation in the conditioned differentiation medium (Figure 2(A)). 3.4. Changes in hEMSCPCs Morphology during Culture in Conditioned Differentiation Medium as Revealed by CM-DiI Labeling To examine cell morphology and cell-cell contacts in detail, cultures were stained with the membrane dye CM-DiI and viewed under laser confocal scanning mi- croscopy. The profiles of differentiated hEMSCPCs were clearly distinguished by CM-DiI, including cell bodies and slender projections that exhibited inflated ends or even claw-like ends (Figures 3(A)-(C)). Many of these CM-DiI-labeled differentiated cells contacted each other, either through projection-soma or projection-projection contacts (Figures 3(A)-(C)). 3.5. Immunofluorescence Staining of Cultured hEMSCPCs Immunofluorescence staining was used to detect the expression of cell-specific markers during culture in the three culture media (growth, NSC, and conditioned dif- ferentiation media). When cultured only in the growth medium, hEMSCPCs were positive for the NSC marker nestin as well as the neural precursor cell and mesen- chymal cell marker vimentin (Figures 3(D) and (E)), but negative for neural cell markers β-III tubulin, micro- tubule-associated protein-2 (MAP2), glial fibrillary acidic protein (GFAP), and the synaptic marker synapsin I (data not shown). After pre-induction in the NSC me- dium, the expression of these markers was not signifi- cantly changed, though cells remained positive for nestin and vimentin (Figure 3(F)). Consistent with the marked morphological transformation (Figures 2 and 3(A)-(C)), cells cultured in the conditioned differentiation medium following pre-induction in NSC medium expressed neu- ral cell markers β-III tubulin, MAP2, GFAP, and synap- sin I (Figures 3(G)-3(J)). In contrast, hEMSCPCs cul- tured in the growth medium but not pre-induced in NSC medium did not express these neural cell markers during culture in conditioned differentiation medium (data not shown). 3.6. Flow Cytometry Analysis Flow cytometry was used to quantify the different protein expression phenotypes. During culture in growth medium, most hEMSCPCs stably expressed the mesen- Copyright © 2012 SciRes. OPEN ACCE SS 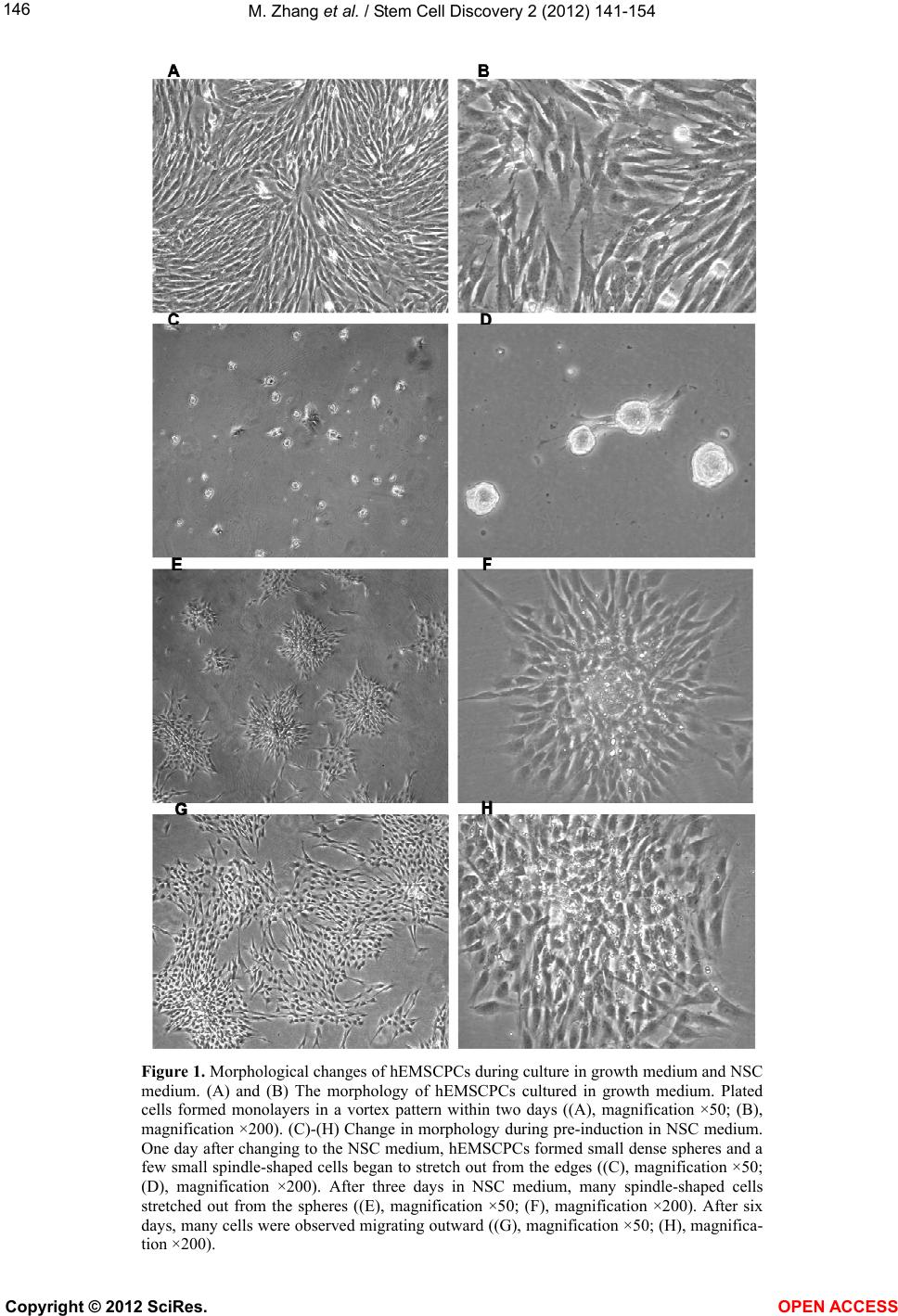 M. Zhang et al. / Stem Cell Discovery 2 (2012) 141-154 Copyright © 2012 SciRes. 146 Figure 1. Morphological changes of hEMSCPCs during culture in growth medium and NSC medium. (A) and (B) The morphology of hEMSCPCs cultured in growth medium. Plated cells formed monolayers in a vortex pattern within two days ((A), magnification ×50; (B), magnification ×200). (C)-(H) Change in morphology during pre-induction in NSC medium. One day after changing to the NSC medium, hEMSCPCs formed small dense spheres and a few small spindle-shaped cells began to stretch out from the edges ((C), magnification ×50; (D), magnification ×200). After three days in NSC medium, many spindle-shaped cells stretched out from the spheres ((E), magnification ×50; (F), magnification ×200). After six days, many cells were observed migrating outward ((G), magnification ×50; (H), magnifica- tion ×200). OPEN ACCESS 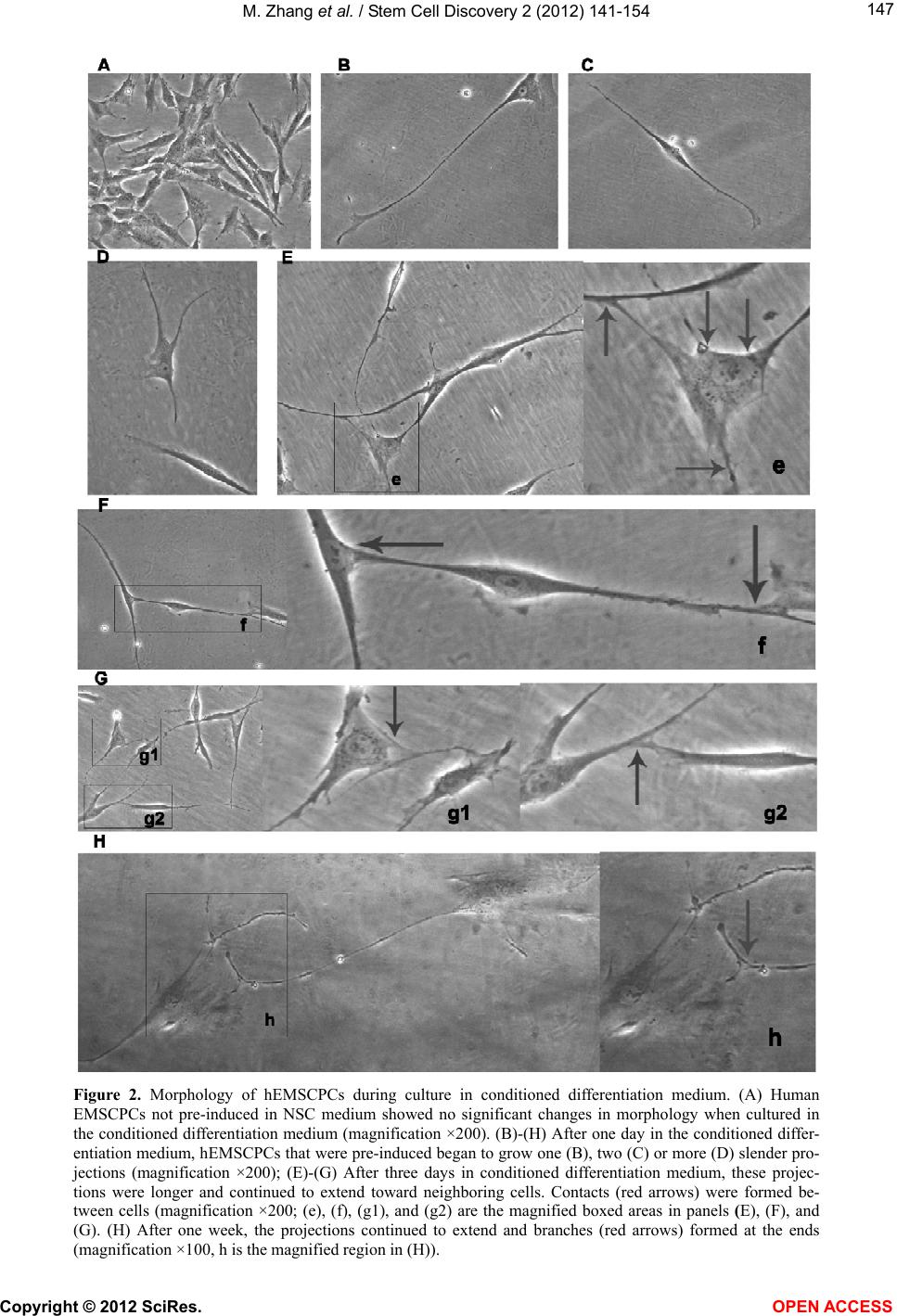 M. Zhang et al. / Stem Cell Discovery 2 (2012) 141-154 147 Figure 2. Morphology of hEMSCPCs during culture in conditioned differentiation medium. (A) Human EMSCPCs not pre-induced in NSC medium showed no significant changes in morphology when cultured in the conditioned differentiation medium (magnification ×200). (B)-(H) After one day in the conditioned differ- entiation medium, hEMSCPCs that were pre-induced began to grow one (B), two (C) or more (D) slender pro- jections (magnification ×200); (E)-(G) After three days in conditioned differentiation medium, these projec- tions were longer and continued to extend toward neighboring cells. Contacts (red arrows) were formed be- tween cells (magnification ×200; (e), (f), (g1), and (g2) are the magnified boxed areas in panels (E), (F), and (G). (H) After one week, the projections continued to extend and branches (red arrows) formed at the ends (magnification ×100, h is the magnified region in (H)). Copyright © 2012 SciRes. OPEN ACCE SS 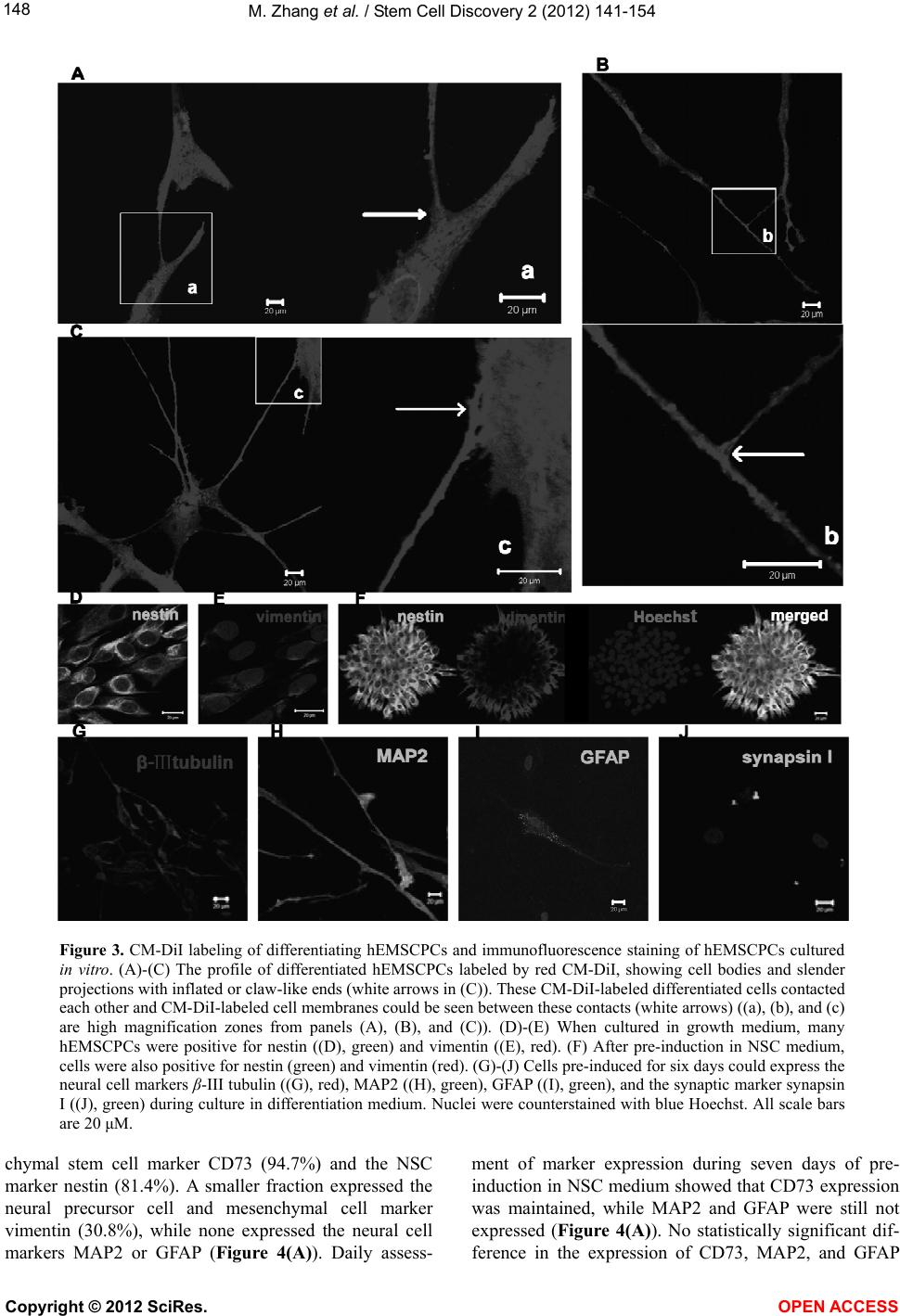 M. Zhang et al. / Stem Cell Discovery 2 (2012) 141-154 Copyright © 2012 SciRes. 148 Figure 3. CM-DiI labeling of differentiating hEMSCPCs and immunofluorescence staining of hEMSCPCs cultured in vitro. (A)-(C) The profile of differentiated hEMSCPCs labeled by red CM-DiI, showing cell bodies and slender projections with inflated or claw-like ends (white arrows in (C)). These CM-DiI-labeled differentiated cells contacted each other and CM-DiI-labeled cell membranes could be seen between these contacts (white arrows) ((a), (b), and (c) are high magnification zones from panels (A), (B), and (C)). (D)-(E) When cultured in growth medium, many hEMSCPCs were positive for nestin ((D), green) and vimentin ((E), red). (F) After pre-induction in NSC medium, cells were also positive for nestin (green) and vimentin (red). (G)-(J) Cells pre-induced for six days could express the neural cell markers β-III tubulin ((G), red), MAP2 ((H), green), GFAP ((I), green), and the synaptic marker synapsin I ((J), green) during culture in differentiation medium. Nuclei were counterstained with blue Hoechst. All scale bars are 20 μM. chymal stem cell marker CD73 (94.7%) and the NSC marker nestin (81.4%). A smaller fraction expressed the neural precursor cell and mesenchymal cell marker vimentin (30.8%), while none expressed the neural cell markers MAP2 or GFAP (Figure 4(A)). Daily assess- ment of marker expression during seven days of pre- induction in NSC medium showed that CD73 expression was maintained, while MAP2 and GFAP were still not expressed (Figure 4(A)). No statistically significant dif- ference in the expression of CD73, MAP2, and GFAP OPEN ACCESS 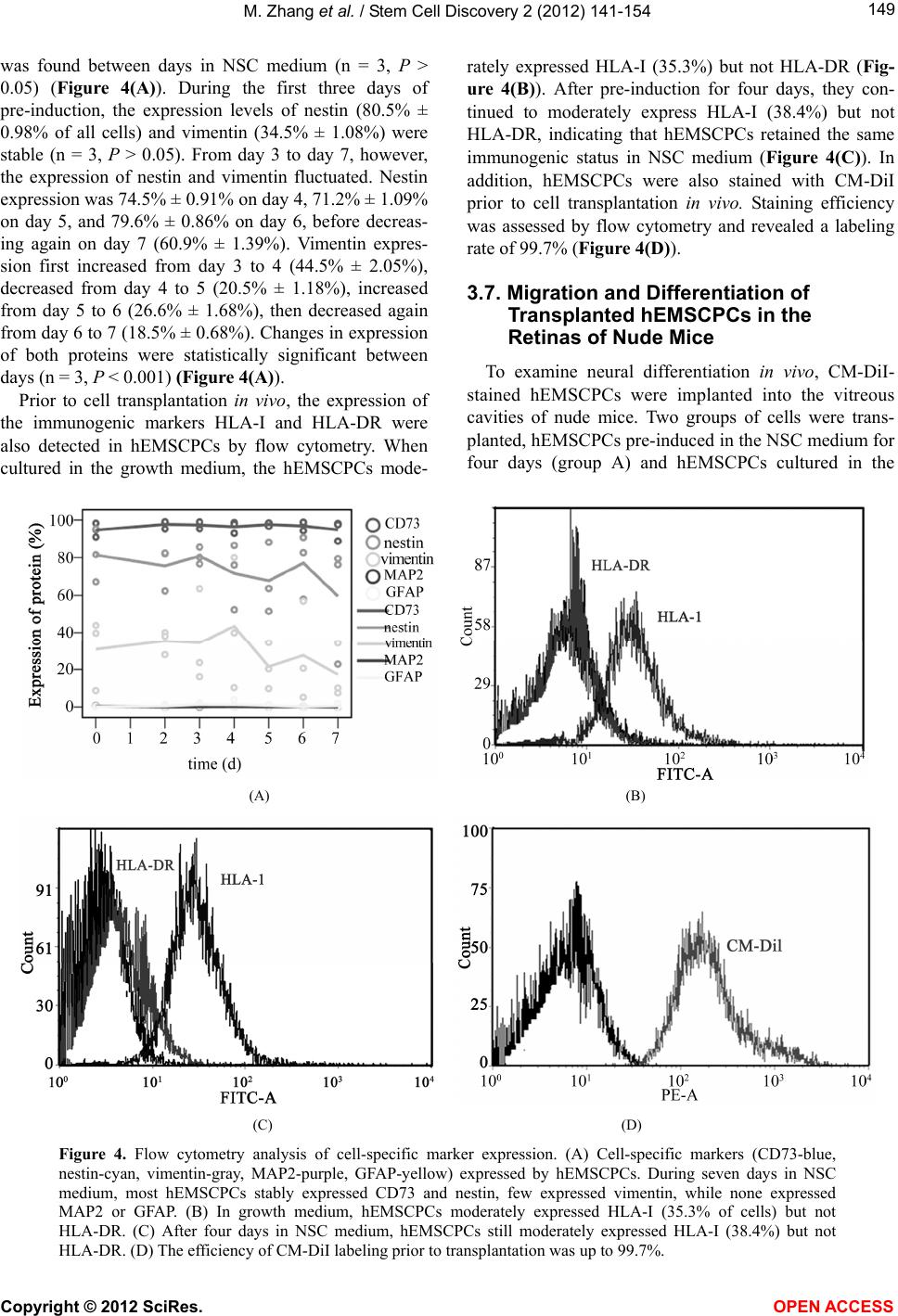 M. Zhang et al. / Stem Cell Discovery 2 (2012) 141-154 149 was found between days in NSC medium (n = 3, P > 0.05) (Figure 4(A)). During the first three days of pre-induction, the expression levels of nestin (80.5% ± 0.98% of all cells) and vimentin (34.5% ± 1.08%) were stable (n = 3, P > 0.05). From day 3 to day 7, however, the expression of nestin and vimentin fluctuated. Nestin expression was 74.5% ± 0.91% on day 4, 71.2% ± 1.09% on day 5, and 79.6% ± 0.86% on day 6, before decreas- ing again on day 7 (60.9% ± 1.39%). Vimentin expres- sion first increased from day 3 to 4 (44.5% ± 2.05%), decreased from day 4 to 5 (20.5% ± 1.18%), increased from day 5 to 6 (26.6% ± 1.68%), then decreased again from day 6 to 7 (18.5% ± 0.68%). Changes in expression of both proteins were statistically significant between days (n = 3, P < 0.001) (Figure 4(A)). Prior to cell transplantation in vivo, the expression of the immunogenic markers HLA-I and HLA-DR were also detected in hEMSCPCs by flow cytometry. When cultured in the growth medium, the hEMSCPCs mode- rately expressed HLA-I (35.3%) but not HLA-DR (Fig- ure 4(B)). After pre-induction for four days, they con- tinued to moderately express HLA-I (38.4%) but not HLA-DR, indicating that hEMSCPCs retained the same immunogenic status in NSC medium (Figure 4(C)). In addition, hEMSCPCs were also stained with CM-DiI prior to cell transplantation in vivo. Staining efficiency was assessed by flow cytometry and revealed a labeling rate of 99.7% (Figure 4(D)). 3.7. Migration and Differentiation of Transplanted hEMSCPCs in the Retinas of Nude Mice To examine neural differentiation in vivo, CM-DiI- stained hEMSCPCs were implanted into the vitreous cavities of nude mice. Two groups of cells were trans- planted, hEMSCPCs pre-induced in the NSC medium for four days (group A) and hEMSCPCs cultured in the (A) (B) (C) (D) Figure 4. Flow cytometry analysis of cell-specific marker expression. (A) Cell-specific markers (CD73-blue, nestin-cyan, vimentin-gray, MAP2-purple, GFAP-yellow) expressed by hEMSCPCs. During seven days in NSC medium, most hEMSCPCs stably expressed CD73 and nestin, few expressed vimentin, while none expressed MAP2 or GFAP. (B) In growth medium, hEMSCPCs moderately expressed HLA-I (35.3% of cells) but not HLA-DR. (C) After four days in NSC medium, hEMSCPCs still moderately expressed HLA-I (38.4%) but not HLA-DR. (D) The efficiency of CM-DiI labeling prior to transplantation was up to 99.7%. Copyright © 2012 SciRes. OPEN ACCE SS  M. Zhang et al. / Stem Cell Discovery 2 (2012) 141-154 150 growth medium but not pre-induced in the NSC medium (group B). Cells of the two groups were separately trans- planted into the vitreous cavities of nude mice. Four weeks later, migration and differentiation in the retina were assessed by immunostaining in frozen sections of the eyes. The retinas of normal uninjected nude mice were clear and regular, with no fluorescent (red- or green-labeled) cells (Figure 5(A)). In mice injected with group A cells, five of six experimental eyes showed mi- gration of transplanted cells after four weeks (Figures 5(B)-(F)). In addition, pre-induced CM-DiI-stained cells expressed the NSC marker nestin and the neural cell markers β-III tubulin and GFAP (Figures 5(I)-(K)), but not MAP2 (data not shown). In mice injected with group B cells, however, only one of six experimental eyes showed migration of transplanted cells (Figures 5(G) and (H)) as well as expression of β-III tubulin (Figure 5(L)). No nestin-, MAP2-, or GFAP-positive cells were found (data not shown). Cells of group A exhibited supe- rior migration and differentiation compared to cells of group B. In both experimental groups, no transplanted cells expressed the photoreceptor cell marker rhodopsin (data not shown). The transplanted cells that migrated into the retinas presented as either single cells or as ag- glomerates (Figures 5(B)-(H)). Most cells concentrated in the subretinal cavities, the retinal pigment epithelium (RPE) layer, and other nearby areas (Figures 5(D)-(F), and 5(H)). Thus, injected cells migrated across all retinal layers to reach the RPE. 4. DISCUSSION The shear size and reparative capacity of human skin makes it an ideal source of pluripotent cells for research and possible autologous cell-based therapies. As a first step toward utilizing these pluripotent cells for neural regeneration therapy, we developed a two step-culture method that gradually induced the appearance of a neu- ral-like morphology and the expression of several neu- ral-specific cell markers. When introduced into the retina, cells from the first culture step (pre-induction) migrated cross multiple cell layers and expressed neural-specific cell markers. Although they were not functionally inte- grated into the healthy retina (at least after four weeks), it is possible that these pre-neural-like cells may be in- duced to replenish lost cells in the degenerating or dam- age retina, such as photoreceptor cells. The capacity of these hEMSCPCs to express neuron-like and glia-like phenotypes in vitro and in vivo suggests that these cells are a potential source for neural stem cells to repair damaged neural tissue. A number of research groups have isolated precursor cells from skin and shown that these cells can differenti- ate into multiple cell types under appropriate conditions. Miller et al. [27,31,43-49] isolated precursors from neo- natal mammalian skin that could differentiate into other cell types, and even mediate regeneration after injury in animal models. They retained a normal karyotype and capacity to differentiate even after regular passage for one year in vitro, but pluripotency was markedly lower in skin-derived precursors from adult mammals. Further- more, the biosafety of adult-derived cells is unknown. Katsuoka et al. [33-35,50-52] isolated a population of stem cells from mammalian dermal hair-follicles that could also differentiate into other cell types and pro- mote regeneration after injury, but they were difficult to isolate in sufficient quantities for clinical applications. In contrast, the hEMSCPCs that we isolated from adult human epidermis were easily obtained in large quantities, could be continuously passaged over fifty times without changes to the normal karyotype, and demonstrate good biosafety in vitro [39]. The growth patterns of hEMSCPCs changed markedly when cultured in NSC medium; individual cells coa- lesced into dense, highly light-reflective spheres. Spin- dle-shaped cells educed from these spheres began to spread out so that the aggregates resembled neural ro- settes. Once these cells were cultured in a conditioned differentiation medium, many differentiated into cells with a neural phenotype. Some cells grew slender pro- jections with inflated or claw-like ends that contacted the soma or projections of other cells as revealed by the cell tracker CM-DiI, a lipid soluble biomembrane stain that stably labels growing cells for clear imaging of fine morphological features [40-42]. Many also expressed neuronal or glial markers, including β-III tubulin, MAP2, GFAP, and synapsin I. Flow cytometry and immunofluo- rescence staining showed that hEMSCPCs expressed the NSC marker nestin, the neural precursor and mesenchy- mal cell marker vimentin, but not the neural cell markers β-III tubulin, MAP2, GFAP, or synapsin I during sequen- tial culture in the growth medium and NSC medium. However, expression of neural cell markers was de- pendent on pre-induction in NSC medium, as hEMSCP- Cs cultured only in differentiation medium did not ex- press neural markers (β-III tubulin, MAP2, GFAP, or synapsin I). Moreover, uninduced cells showed inferior migration and differentiation compared to pre-induced cells after transplantation into the vitreous cavities of nude mice. Thus, pre-induction did not markedly alter neural marker expression in hEMSCPCs but was neces- sary to allow differentiation in a special conditioned me- dium and in the mouse eye (part of the central nervous system). Flow cytometry was used to quantify the expression of cell-specific markers. The cell surface glucoprotein CD73 is a marker for mesenchymal stem cells [53]. Nestin is a class VI intermediate filament protein once thought to be a specific marker of NSCs, but recent re- Copyright © 2012 SciRes. OPEN ACCE SS  M. Zhang et al. / Stem Cell Discovery 2 (2012) 141-154 151 Figure 5. Migration and differentiation of transplanted hEMSCPCs in the retinas of nude mice. (A) The retinas of normal nude mice were clear and regular, and no red- or green-labeled cells were observed. (B)-(L) Four weeks after transplantation into the vitreous cavities of nude mice, many pre-induced hEMSCPCs (white arrows) had mi- grated into the retinal ganglion cell layer (B), (C), the sub-retinal space (D), (E), and the retinal pigment epithelium layer (E), (F). In addition, pre-induced cells expressed β-III tubulin (I), GFAP (J), and nestin (K). Transplanted cells that had not been pre-induced by NSC medium in vitro (white arrows) could migrate into the retinal ganglion cell layer (G) and the sub-retinal space (H), and express β-III tubulin (L). “V” represents the vitreous cavities or nearby areas. Nuclei were counterstained with blue Hoechst. All scale bars represent 20 μM. Copyright © 2012 SciRes. OPEN ACCE SS  M. Zhang et al. / Stem Cell Discovery 2 (2012) 141-154 152 ports indicated that it is also expressed by multipotent precursor cells and that expression correlates with the potential for proliferation, migration, and differentiation [54,55]. Vimentin is a class III intermediate filament protein and a marker for neural precursor cells and mes- enchymal-derived cells [56-59]. MAP2 is a mature neu- ronal marker, while the intermediate filament protein GFAP is expressed almost exclusively by astrocytes. Flow cytometry showed that the expression levels of these markers by hEMSCPCs were not significantly changed during culture in growth medium or brief (2 - 3 day) culture in NSC medium. However, from the 3rd to 7th day in NSC medium, expression of nestin and vi- mentin began to fluctuate, possibly indicating the be- ginning of a phenotypic transition. However, hEMSCPCs pre-induced in NSC medium grew slender projections that could be easily sheared off the soma during harvest- ing for flow cytometry, so this fluctuation in expression may reflect variable loss of markers localized to pro- cesses. A more accurate determination of marker expres- sion patterns during NSC culture will require the use of alternative methods like real time PCR or Western blot- ting. Approximately the same percentage of pre-induced and uninduced hEMSCPCs expressed HLA-I, indicating that these cells have a stable immunogenicity. Nude mice were chosen for hEMSCPC transplantation experiments because they are immunologically defective and so would not generate an immune response to transplanted cells. Four weeks after injection of hEMSCPCs, both in- duced and uninduced CM-DiI-labeled cells had migrated primarily to the subretinal space, retinal pigment epithet- lium layer, and other nearby areas. The reasons for this selective migration are still obscure. Both hEMSCPCs and retinal pigment epithelium originate from the ec- toderm, and this homology may have allowed for better survival of transplanted hEMSCPCs. The subretinal ca- vity may also accumulate transplanted cells even in im- munologically active mice because it is an immunopri- vileged zone. While both induced and uninduced cells migrated, pre-induced cells showed superior migration and neural differentiation. There were differences in neural differentiation in vi- tro versus in vivo. In contrast to observations in vitro, the mature neuron marker MAP-2 was not expressed by ei- ther group of transplanted hEMSCPCs, although this might reflect the short transplantation time (four weeks). In addition to common markers like MAP-2, transplanted cells did not express retina-specific markers. For exam- ple, rhodopsin, a marker of retinal photoreceptors, was not expressed by transplanted hEMSCPCs that migrated into the retina, possibly because the healthy retina re- leases no factors that would induce differenttiation. A longer transplantation time or induction of growth factor expression may be required for full expression of neural markers, but these questions require further study. The epidermis is easily harvested, so hEMSCPCs may be a productive source of cells for autologous cell-based therapy against neurological diseases. Our results show that hEMSCPCs possess the capacity to differentiate into neural-like cells if pre-induced in NSC medium. Whether hEMSCPCs can replenish endogenous neural cells after induction and differentiation requires further study. To reach this goal, further studies are required to understand the neural lineage capacity of these cells, including tests of differentiation efficiency and function in vitro, and to assess the biological safety of these cells in vivo. Ex- periments testing the restorative efficacy of these cells in animal models of neurodegenerative diseases are clearly warranted. 5. ACKNOWLEDGEMENTS We are grateful to members of the State Key Laboratory of Oph- thalmology, Zhongshan Ophthalmic Center, Sun Yat-sen University, for discussion and advice. This work was supported by the Science and Technology Projects of Guangdong Province, China (2009B060600002, 2010B060500006). REFERENCES [1] Jiang, W. (2010) Neurology. 2nd Edition, People’s Medi- cal Publishing House, Beijing. [2] Carsten, R.B. and Jens, C.S. (2004) Therapeutic strategies for neurodegenerative disorders: Emerging clues from Parkinson’s disease. Biological Psychiatry, 56, 213-216. doi:10.1016/j.biopsych.2003.12.025 [3] John, B.S. (2004) Pharmacotherapeutic approaches to the treatment of Alzheimer’s disease. Clinical Therapeutics, 26, 615-630. doi:10.1016/S0149-2918(04)90064-1 [4] Zhou, J.W. (2010) Recent progress in neurodegenerative disorder research in China. Science China Life Sciences, 53, 348-355. doi:10.1007/s11427-010-0061-0 [5] Peng, L.S. and Li, C.R. (2009) RNA interference and neural degenerative diseases. West China Medical Jour- nal, 21, 1806-1808. [6] William, J.M., Raymond, T.B., Joao, S., et al. (2010) Gene delivery of AAV2-neurturin for Parkinson’s disease: A double-blind, randomised, controlled trial. The Lancet Neurology, 9, 1164-1172. doi:10.1016/S1474-4422(10)70254-4 [7] Wu, J.J., Yu, W.B., Chen, Y., et al. (2010) Intrastriatal transplantation of GDNF-engineered BMSCs and its neuroprotection in Lactacystin-induced Parkinsonian Rat Model. Neurochemical Research, 35, 495-502. doi:10.1007/s11064-009-0086-6 [8] William, J.M., Jill, L.O., Leonard, V., et al. (2008) Safety and tolerability of intraputaminal delivery of CERE-120 (adeno-associated virus serotype 2-neurturin) to patients with idiopathic Parkinson’s disease: An open-label, phase Copyright © 2012 SciRes. OPEN ACCE SS  M. Zhang et al. / Stem Cell Discovery 2 (2012) 141-154 153 I trial. The Lancet Neurology, 7, 400-408. doi:10.1016/S1474-4422(08)70065-6 [9] Kerri, S. (2010) Treatment frontiers. Nature, 466, S15- S18. doi:10.1038/nature09476 [10] Jiao, J.W. (2010) Embryonic and adult neural stem cell research in China. Science China Life Sciences, 53, 338- 341. doi:10.1007/s11427-010-0070-z [11] Stefano, P., Lucia, Z., Michela, D. and Gianvito, M. (2005) Neural stem cells and their use as therapeutic too in neurological disorders. Brain Research Reviews, 48, 211-219. doi:10.1016/j.brainresrev.2004.12.011. [12] Gu, Y., Hu, N., Liu, J., et al. (2010) Isolation and differ- entiation of neural stem/progenitor cells from fetal rat dorsal root ganglia. Science China Life Sciences, 53, 1057-1064. doi:10.1007/s11427-010-4053-x [13] Keun-Hwa, J., Kon, C., Soon-Tae, L., et al. (2008) Iden- tification of neuronal outgrowth cells from peripheral blood of stroke patients. Annals of Neurology, 63, 312- 322. doi:10.1002/ana.21303 [14] Sarugaser, R., Ennis, J., Stanford, W.L. and Gianvito, M. (2009) Isolation, propagation, and characterization of hu- man umbilical cord perivascular cells (HUCPVCs). Me- thods in Molecular Biology, 482, 269-279. doi:10.1007/978-1-59745-060-7-17 [15] Lin-ya, H., Jia-lin, Y., Fang, L., et al. (2009) Synapse function of neuron-like cells induced from mesenchymal stem cells by Salvia miltiorrhiza. Acta Academiae Medi- cinae Militaris Tertiae, 31, 144-147. doi:1000-5404(2009)02-0144-04 [16] Shi, Y.F., Hu, G.Z., Su, J.J., et al. (2010) Mesenchymal stem cells: A new strategy for immunosuppression and tissue repair. Cell Research, 20, 510-518. doi:10.1038/cr.2010.44 [17] Wu, L., Chen, R.K., Yang, L., et al. (2009) Differentia- tion of adipose-derived stem cells into nerve stem cells across embryonic layer. Fourth Military Medical Univer- sity, 30, 70-72. doi:1000-2790(2009)01-0070-03 [18] Stefan, A., Helmut, K., Irina, S., et al. (2004) Neurally selected embryonic stem cells induce tumor formation after long-term survival following engraftment into the subretinal space. Investigative Ophthalmology & Visual Science, 45, 4251-4255. doi:10.1167/iovs.03-1108 [19] Oscar, H.M. and Angela, N. (2010) Epithelial plasticity, stemness and pluripotency. Cell Research, 20, 1086-1088. doi:10.1038/cr.2010.127 [20] Dengke, K.M., Bonaguidi, M.A., Ming, G.L. and Song, H.J. (2009) Adult neural stem cells in the mammalian central nervous system. Cell Research, 19, 672-682. doi:10.1038/cr.2009.56 [21] Chase, L.G, Lakshmipathy, U, Solchaga, L.A., et al. (2010) A novel serum-free medium for the expansion of human mesenchymal stem cells. Stem Cell Research & Therapy, 1, 8. doi:10.1186/scrt8 [22] Raymond, D.L., Shaomei, W., Bin, L., et al. (2007) Cells isolated from umbilical cord tissue rescue photoreceptors and visual functions in a rodent model of retinal disease. Stem Cells, 25, 602-611. doi:10.1634/stemcells.2006-0308 [23] Andrew, J.H., Isabel, Z., Henry, H.T., et al. (2009) Hu- man umbilical cord blood-derived mesenchymal stem cells do not differentiate into neural cell types or integrate into the retina after intravitreal grafting in neonatal rats. Stem Cells and Development, 18, 399-409. doi:10.1089/scd.2008.0084 [24] Isabel, Z., Andrew, J.H., Faisal, A., et al. (2009) Umbili- cal cord blood mesenchymal stromal cells are neuropro- tective and promote regeneration in a rat optic tract model. Experimental Neurology, 216, 439-448. doi:10.1016/j.expneurol.2008.12.028 [25] Xue, G.S., Zhang, Y. and Qi, Z.L. (2008) Current research situation of adipose-derived stem cells and its application in tissue engineering. Journal of Tissue Engineering and Reconstructive Surgery, 4, 174-176. [26] Hideo, O., Ariane, R., Ce’cile, K., et al. (2001) Morpho- genesis and renewal of hair follicles from adult multipo- tent stem cells. Cell, 104, 233-245. doi:10.1016/S0092-8674(01)00208-2 [27] Jean, G.T., Mahnaz, A., Karl, J.L., et al. (2001) Isolation of multipotent adult stem cells from the dermis of mam- malian skin. Nature Cell Biology, 3, 778-786. doi:10.1038/ncb0901-778 [28] Young, H.E., Steele, T.A., Bray, R.A., et al. (2001) Hu- man reserve pluripotent mesenchymal stem cells are pre- sent in the connective tissues of skeletal muscle and der- mis derived from fetal, adult, and geriatric donors. Ana- tomy & Physiology, 264, 51-62. doi:10.1002/ar.1128 [29] Shi, C. and Cheng, T. (2003) Effects of acute wound en- vironment on the neonatal dermal multipotent cells. Cells Tissues Organs, 175, 177-185. doi:10.1159/000074939 [30] Shih, D.T. (2005) Isolation and characterization of neu- rogenic mesenchymal stem cells in human scalp tissue. Stem Cells, 23, 1012-1025. doi:10.1634/stemcells.2004-0125 [31] Jean, G.T., Ian, A.M., Darius, B. and Freda, D.M. (2005) Isolation and characterization of multipotent skin-derived precursors from human skin. Stem Cells, 23, 727-737. doi:10.1634/stemcells.2004-0134 [32] Karl, J.L. Fernandes, I.A. McKenzie, Pleasantine, M. et al. (2004) A dermal niche for multipotent adult skin-de- rived precursor cells. Nature Cell Biology, 6, 1082-1093. doi:10.1038/ncb1181 [33] Yasuyuki, A., Lingna, L., Kensei, K. and Robert, M.H. (2004) Multipotent hair follicle stem cells promote repair of spinal cord injury and recovery of walking function. Cell Cycle, 7, 1865-1869. doi:10.4161/cc.7.12.6056 [34] Yasuyuki, A., Lingna, L., Kensei, K., Sheldon, P. and Robert, M.H. (2005) Multipotent nestin-positive, keratin- negative hair-follicle bulge stem cells can form neurons. Proceedings of the National Academy of Sciences, 102, 5530-5534. doi:10.1073/pnas.0501263102 [35] Yasuyuki, A., Lingna, L., Raul, C., et al. (2005) Implan- ted hair follicle stem cells form Schwann cells that sup- port repair of severed peripheral nerves. Proceedings of the National Academy of Sciences, 102, 17734-17738. doi:10.1073/pnas.0508440102 [36] Patrizia, T., Jeff, W.M., Maria, G.B., et al. (2006) Brain Copyright © 2012 SciRes. OPEN ACCE SS  M. Zhang et al. / Stem Cell Discovery 2 (2012) 141-154 Copyright © 2012 SciRes. OPEN ACCE SS 154 engraftment and therapeutic potential of stem/progenitor cells derived from mouse skin. The Journal of Gene Medicine, 8, 506-513. doi:10.1002/jgm.866 [37] So, P.L. and Epstein, E.H. (2004) Adult stem cells: Cap- turing youth from a bulge. Trends in Biotechnology, 22, 493-496. doi:10.1016/j.tibtech.2004.08.007 [38] Morasso, M.I. and Omic-Canic, M. (2005) Epidermal stem cells: The cradle of epidermal determination, dif- ferentiation and wound healing. Biology of the Cell, 97, 173-183. doi:10.1042/BC20040098 [39] Huang, B., Li, K.J., Yu, J., et al. (2011) Generation of Human epidermis-derived mesenchymal stem cell-like pluripotent cells and their reprogramming in mouse chi- meras. http://precedings.nature.com/documents/6016/version/1 [40] Yamamoto, N., Higashi, S. and Toyama, K. (1997) Stop and branch behaviors of geniculocortical axons: A time- lapse study in organotypic cocultures. The Journal of Neuroscience, 17, 3653-3663. [41] Cai, Q., Ji, M., Zhang, J., et al. (2011) Comparative study on glutamatergic synaptic connections in rat striatum with laser scanning confocal microscopy and electron micros- copy. Chinese Journal of Histochemistry and Cytoche- mistry, 20, 236-240. [42] Matsubayashi, Y., Iwai, L. and Kawasaki, H. (2008) Fluo- rescent double-labeling with carbocyanine neuronal trac- ing and immunohistochemistry using a cholesterol-spe- cific detergent digitonin. Journal of Neuroscience Me- thods, 174, 71-81. doi:10.1016/j.jneumeth.2008.07.003 [43] Ian, A.M., Jeff, B., Jean, G.T., et al. (2006) Skin-derived precursors generate myelinating Schwann cells for the in- jured and dysmyelinated nervous system. Journal of Neu- roscience Methods, 26, 6651-6660. doi:10.1523/JNEUROSCI.1007-06.2006 [44] Karl, J.L., Fernandes, N.R., Kobayashi, C.J. et al. (2006) Analysis of the neurogenic potential of multipotent skin- derived precursors. Experimental Neurology, 201, 32-48. doi:10.1016/j.expneurol.2006.03.018 [45] Jeff, B., Joseph, S.S., Liu, J., et al. (2007) Skin-derived Precursors generate myelinating Schwann cells that pro- mote remyelination and functional recovery after contu- sion spinal cord injury. Journal of Neuroscience Methods, 27, 9545-9559. doi:10.1523/JNEUROSCI.1930-07.2007 [46] Karl, J.L., Jean, G.T. and Freda, D.M. (2008) Multipotent skin-derived precursors: Adult neural crest-related pre- cursors with therapeutic potential. Philosophical Trans- actions of the Royal Society B, 363, 185-198. doi:10.1098/rstb.2006.2020 [47] Jean-Francois, L., Jeffrey, A.B., Yan, C., et al. (2009) Skin-derived precursors differentiate into skeletogenic cell types and contribute to bone repair. Stem Cells and Development, 18, 893-905. doi:10.1089/scd.2008.0260 [48] Jeffrey, B., Maryline, P., Olena, M., et al. (2009) SKPs derive from hair follicle precursors and exhibit properties of adult dermal stem cells. Cell Stem Cell, 5, 610-623. doi:10.1016/j.stem.2009.10.019 [49] Hiroyuki, J., Olena, M., Jones, K.L., et al. (2010) Con- vergent genesis of an adult neural crest-like dermal stem cell from distinct developmental origins. Stem Cells, 28, 2027-2040. doi:10.1002/stem.525 [50] Yasuyuki, A., Lingna, L., Kensei, K. and Robert, M.H. (2010) Embryonic development of hair follicle pluripo- tent stem (hfPS) cells. Medical Molecular Morphology, 43, 123-127. doi:10.1007/s00795-010-0498-z [51] Yasuyuki, A., Kensei, K., Robert, M.H. (2010) The ad- vantages of hair follicle pluripotent stem cells over em- bryonic stem cells and induced pluripotent stem cells for regenerative medicine. Journal of Dermatological Sci- ence, 60, 131-137. doi:10.1016/j.jdermsci.2010.09.007 [52] Fang, L., Aisada, U., Hiroaki, K., et al. (2010) The bulge area is the major hair follicle source of nestin-expressing pluripotent stem cells which can repair the spinal cord compared to the dermal papilla. Cell Cycle, 10, 830-839. doi:10.4161/cc.10.5.14969 [53] Florian, H., Wolf, C.P., David, A., et al. (2009) Morpho- logical and immunocytochemical characteristics indicate the yield of early progenitors and represent a quality con- trol for human mesenchymal stem cell culturing. Journal of Anatomy, 214, 759-767. doi:10 .1111/j.1469-7580.2009.01065.x [54] Su, P.H., Wang, T.C., Wong, Z.R., et al. (2011) The ex- pression of nestin delineates skeletal muscle differentia- tion in the developing rat esophagus. Journal of Anatomy, 218, 311-323. doi:10.1111/j.1469-7580.2010.01331.x. [55] Svachovaa, H., Pour, L., Sana, J., et al. (2011) Stem cell marker nestin is expressed in plasma cells of multiple myeloma patients. Leukemia Research, 35, 1008-1013. doi:10.1016/j.leukres.2011.03.001 [56] Frederiksen, K. and McKay, R.D.G. (1988) Proliferation and differentiation of rat neuroepithelial precursor cells in vivo. The Journal of Neuroscience, 8, 1144-l151. [57] Yvan, A., Jean-Guy, V., Jean-Francois, B., et al. (2001) Isolation of multipotent neural precursors residing in the cortex of the adult human brain. Experimental Neurology, 170, 48-62. doi:10.1006/exnr.2001.7691 [58] Jahan, A., Saskia, F., Anli, Z. and Melissa, F. (2010) Cha- racterization of neural stem/progenitor cells expressing VEGF and its receptors in the subventricular zone of newborn piglet brain. Neurochemical Research, 35 , 1455- 1470. doi:10.1007/s11064-010-0207-2 [59] Chanchai, B., Kerstin, K., Sombat B., et al. (2011) Fibro- sis and evidence for epithelial-mesenchymal transition in the kidneys of patients with staghorn calculi. British Jour- nal of Urology International, 107, 1847. doi:10.1111/j.1464-410X.2011.10350.x
|