S. DAS ET AL. 239
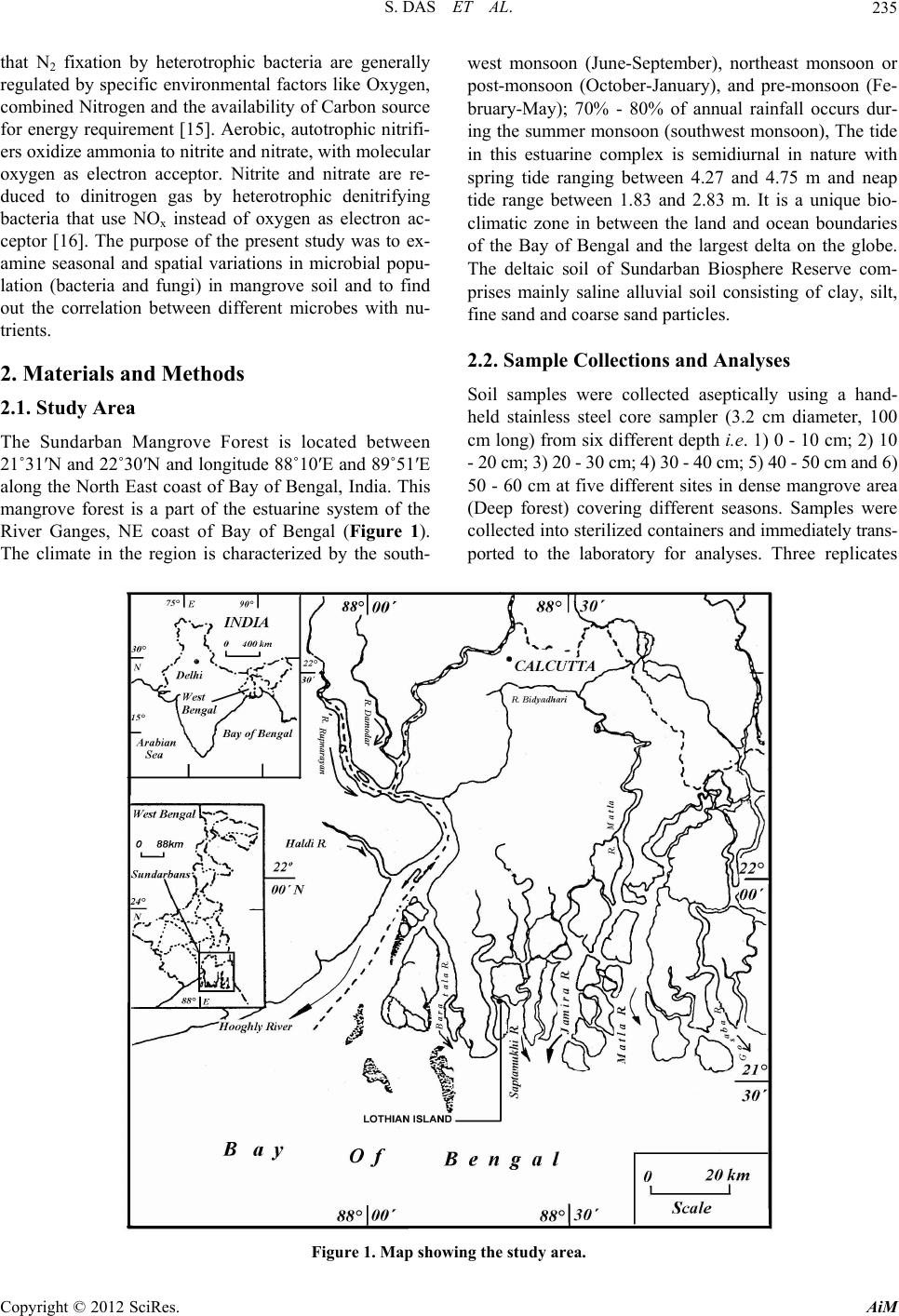
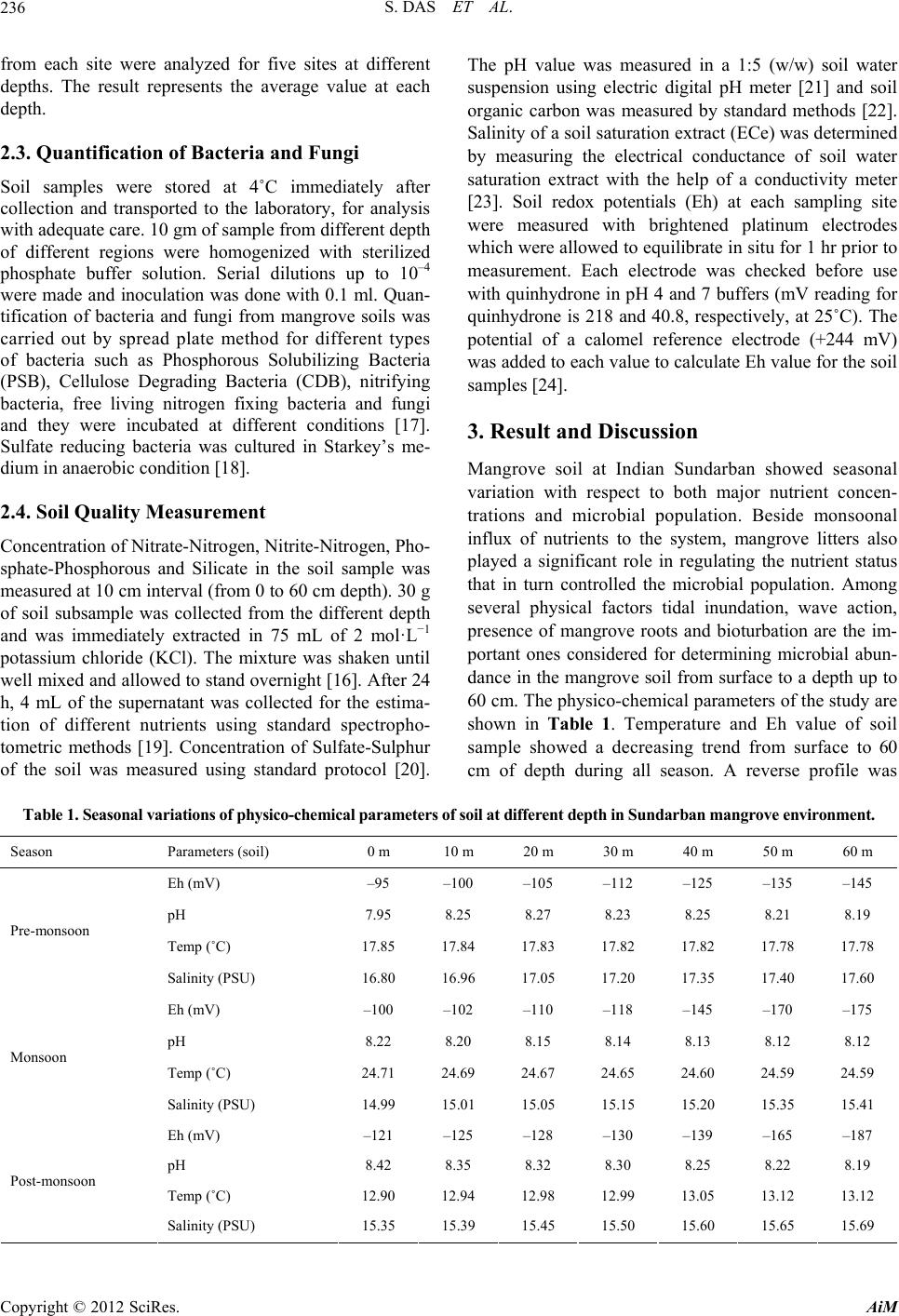
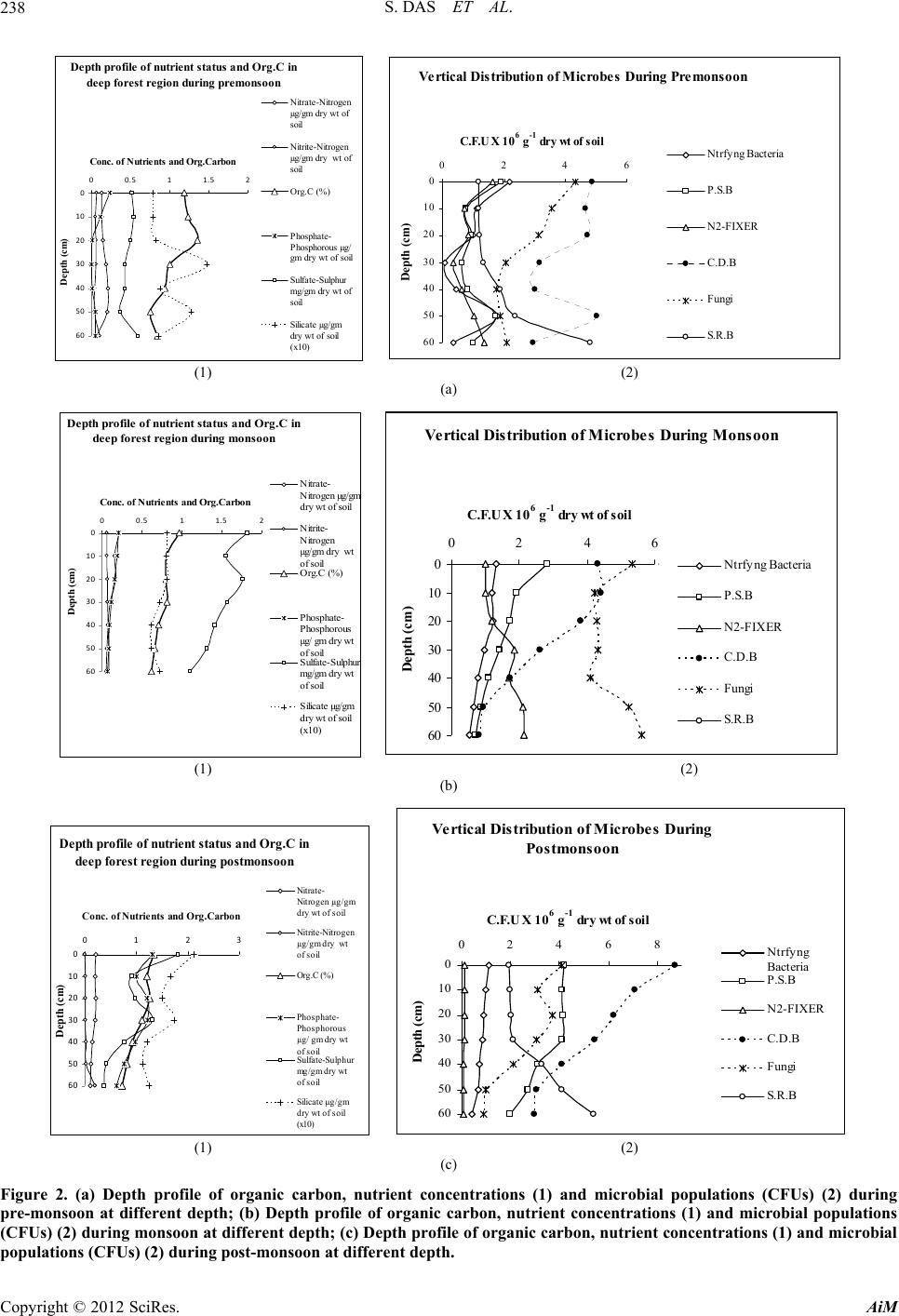
fungi. Other groups of microbes have also shown sig-
nificant population which is a good sign for a mangrove
forest with respect to mineralization of organic debris
and as a result mangrove plants can easily get nutrients in
their simplest forms. From seasonal perspective, the
monsoon period was significantly different (Student’s t
test, P < 0.01) than other periods of the year in terms of
total bacterial abundance and other key parameters. Sea
level rise due to global warming may hamper the stable
ecological zone of Sundarban mangrove forest which
may ultimately reflect to net flux of several biologically
active trace gases between soil and atmosphere. Intro-
duction of huge amount of nutrients during monsoon
have a positive feedback on the bacterial population of
mangrove sediment. Beside the changes in several phys-
icochemical parameters, transport of huge amount of
aquatic microbes could lead to the significant increase in
the microbial population in the soil of this mangrove eco-
system. This may contribute to the aquatic biogeochem-
istry of this tropical wetland.
4. Acknowledgements
The financial assistance from DOEn, Govt. of West
Bengal and U.G.C., New Delhi are gratefully acknowl-
edged. The authors are also grateful to the Forest De-
partment, Govt. of West Bengal for assisting the research
team in collecting data and providing all infrastructural
facilities to reach the remote island.
REFERENCES
[1] B. Gonzalez-Acosta, Y. Bashan, N. Y. Hernandez-Sa-
avedra, F. Ascenaio and G. Cruz-Aguero, “Seasonal
Seawater Temperature as the Major Determinant for
Populations of Culturable Bacteria in the Soils of an In-
tact Mangrove in an Arid Region,” FEMS Microbiology
Ecology, Vol. 55, No. 2, 2006, pp. 311-321.
doi:10.1111/j.1574-6941.2005.00019.x
[2] D. M. Alongi, K. G. Boto and F. Tirendi, “Effect of Ex-
ported Mangrove Litter on Bacterial Productivity and
Dissolved Organic Carbon Fluxes in Adjacent Tropical
Nearshore Soils,” Marine Ecology Progress Series, Vol.
56, 1989, pp. 133-144. doi:10.3354/meps056133
[3] D. M. Alongi, P. Christofferson and F. Tirendi, “The
Influence of Forest Type on Microbial-Nutrient Rela-
tionship in Tropical Mangrove Soil,” Journal of Experi-
mental Marine Biology and Ecology, Vol. 171, No. 2,
1993, pp. 201-223.
doi:10.1016/0022-0981(93)90004-8
[4] D. M. Alongi, “The Role of Bacteria in Nutrient Recy-
cling in Tropical Mangrove and Other Coastal Benthic
Ecosystems,” Hydrobiologia, Vol. 285, No. 1-3, 1994, pp.
19-32. doi:10.1007/BF00005650
[5] G. Holguin, Y. Bashan and P. Vazavez, “The Role of Soil
Microorganism in the Productivity, Conservation and
Rehabilitation of Mangrove Ecosystem: An Overview,”
Biology of Fertile Soils, Vol. 33, No. 4, 2001, pp. 265-
278. doi:10.1007/s003740000319
[6] G. Toledo, Y. Bashan and A. Soeldner, “Cyanobacteria
and Black Mangroves in Northwestern Mexico: Coloni-
zation, and Diurnal and Seasonal Nitrogen Fixation on
Aerial Roots,” Canadian Journal of Microbiology, Vol.
41, No. 11, 1995, pp. 999-1011.
doi:10.1139/m95-139
[7] A. Rojas, G. Holguin, B. R. Glick and Y. Bashan, “Syn-
ergism between Phyllobacterium sp. (N2-Fixer) and Ba-
cillus licheniformis (P-Solubilizer), both from a Semiarid
Mangrove Rhizosphere,” FEMS Microbiology Ecology,
Vol. 35, 2001, pp. 181-187.
doi:10.1111/j.1574-6941.2001.tb00802.x
[8] P. Vazquez, G. Holguin, M. E. Puente, A. Lopez-Cortes
and Y. Bashan, “Phosphate-Solubilizing Microorganisms
Associated with the Rhizosphere of Mangroves in a
Semiarid Coastal Lagoon,” Biology and Fertility of Soils,
Vol. 30, No. 5-6, 2000, pp. 460-468.
doi:10.1007/s003740050024
[9] D. B. Nedwell, T. H. Blackburn and W. J. Wiebe, “Dy-
namic Nature of the Turnover of Organic Carbon, Nitro-
gen and Sulpher in the Soil of a Jamaican Mangrove For-
est,” Marine Ecology Progress Series, Vol. 110, No. 9,
1994, pp. 223-231. doi:10.3354/meps110223
[10] E. Senior, E. B. Lindstrom, I. M. Banat and D. B. Ned-
well, “Sulfate Reduction and Methanogenesis in the
Sediment of a Saltmarsh on the East Coast of the United
Kingdom,” Applied Environmental Microbiology, Vol. 43,
1982, pp. 987-996.
[11] J. Sorensen, B. B. Jorgensen and N. P. Revsbech, “A
Comparison of Oxygen, Nitrate and Sulfate Respiration
in Coastal Marine Soil,” Microbial Ecology, Vol. 5, No. 2,
1979, pp. 105-111. doi:10.1007/BF02010501
[12] D. R. Lovley and M. J. Klug, “Intermediary Metabolism
of Organic Matter in the Soil of a Eutrophic Lake,” Ap-
plied Environmental Microbiology, Vol. 43, 1982, pp.
552-560.
[13] R. Mohanraju and R. Natarajan, “Methanogenic Bacteria
in Mangrove Soils,” Hydrobiologia, Vol. 247, No. 1-3,
1992, pp. 187-193. doi:10.1007/BF00008218
[14] C. R. Wang, Y. Shi, X. M. Yang, J. Wu and J. Yue, “Ad-
vances of Study on Atmospheric Methane Oxidation
(Consumption) in Forest Soil,” Journal of Forestry Re-
search, Vol. 14, No. 3, 2003, pp. 230-238.
doi:10.1007/BF02856837
[15] C. B. Teri and K. F. Mary, “Linking Microbial Com-
munity Composition and Soil Processes in a California
Annual Grassland and Mixed Conifer Forest,” Biogeo-
chemistry, Vol. 73, No. 2, 2005, pp. 395-415.
doi:10.1007/s10533-004-0372-y
[16] R. H. Riley, M. Peter and P. M. Vitousek, “Nutrient Dy-
namics and Nitrogen Trace Gas Flux during Ecosystem
Development in Montane Rain Forest,” Ecology, Vol. 76,
No. 1, 1995, pp. 292-304. doi:10.2307/1940650
[17] A. L. Ramanathan, G. Singh, J. Majumder, A. C. Samal,
R. Chowhan, R. K. Rayan, K. Roykumar and S. C. Santra,
Copyright © 2012 SciRes. AiM