A. W. A. VAN GEMMERT
Copyright © 2012 SciRes. 87
occurring at the same time as when the next target appears will
improve detection of the go-stimulus. Both of these tone condi-
tions could result in narrowing the focus of attention on the
point-to-point aiming task which means that all available atten-
tional resources would be directed to the most optimal per-
formance of the task. Furthermore, when the tone is presented
during execution it could act as a distraction resulting in dimin-
ishing some of the resources used to execute the point-to-point
aiming task. When compared to the condition when no tones
were presented during acquisition, these assumptions about
tones affecting attentional resource allocation should have re-
sulted into improved retention performance for the two tone
conditions and deteriorated performance for the condition in
which a tone was presented during execution of the aiming
movement. Moreover, this prediction for the pattern of reten-
tion is based on the view that when more attentional resources
are dedicated to the acquisition of a task, the task is learned
better and thus performance during retention should be better
than retention performance when less attentional resources were
dedicated to the task during training. In the current study this
pattern of retention performance was not found, because reten-
tion performance was the same for all conditions, therefore it is
deemed to be unlikely that the tone presentation in the current
study altered normal management of attentional resources. Off
course, it should be noted that the current study did not include
conditions in which the participants were instructed to attend to
the tones so one cannot make strong arguments about the use of
attentional resources and its effects on task performance.
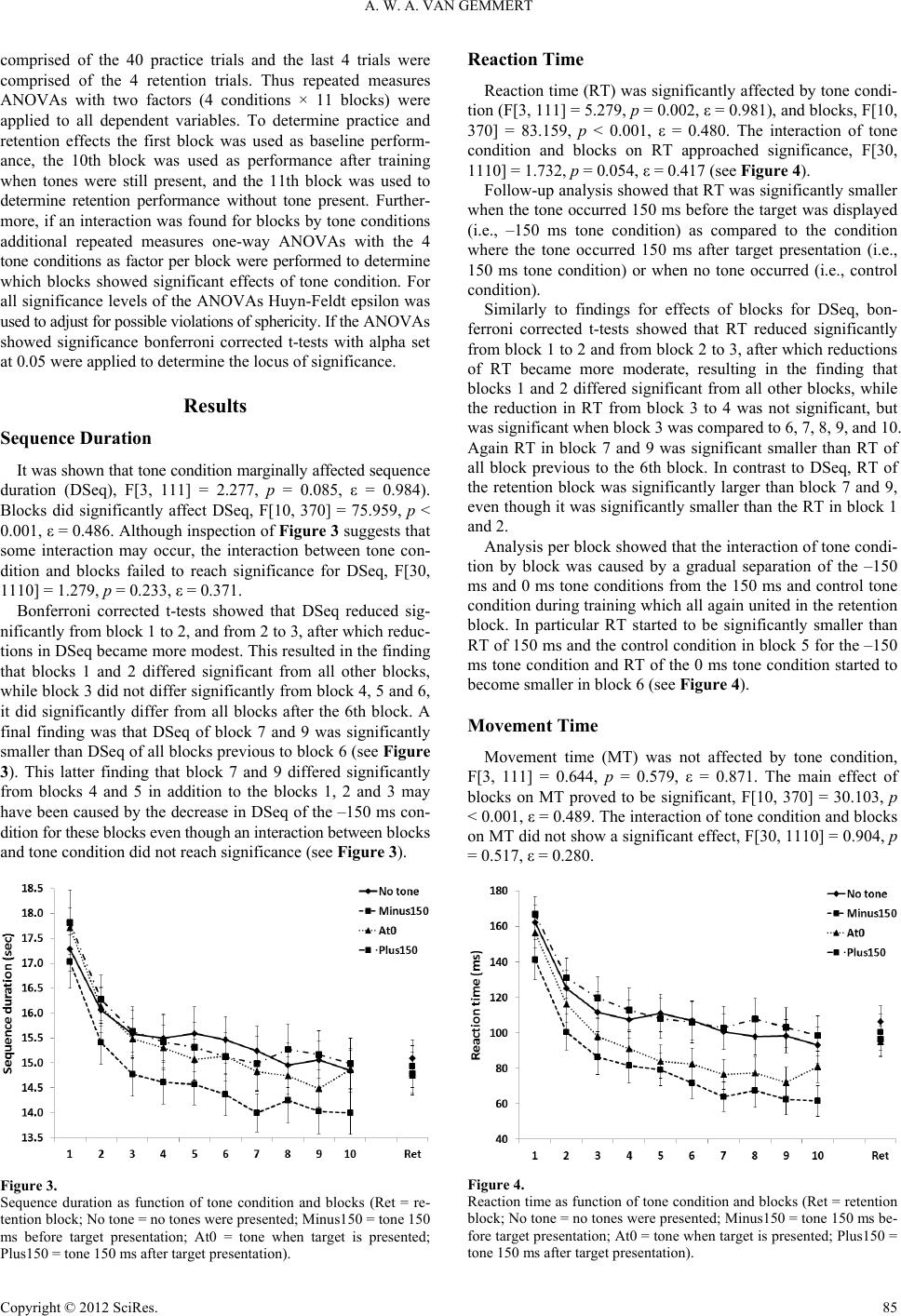
In depth analyses, in which each movement sequence was
parsed into a reaction and execution portion, showed that reac-
tion time decreased, in addition to the normal reductions ob-
served during acquisition, when tones were presented before
and at the same time as the presentation of the movement target.
Nevertheless, these benefits for reaction time during acquisition
did not translate to additional benefits for retention performance
when compared to retention performance without tones.
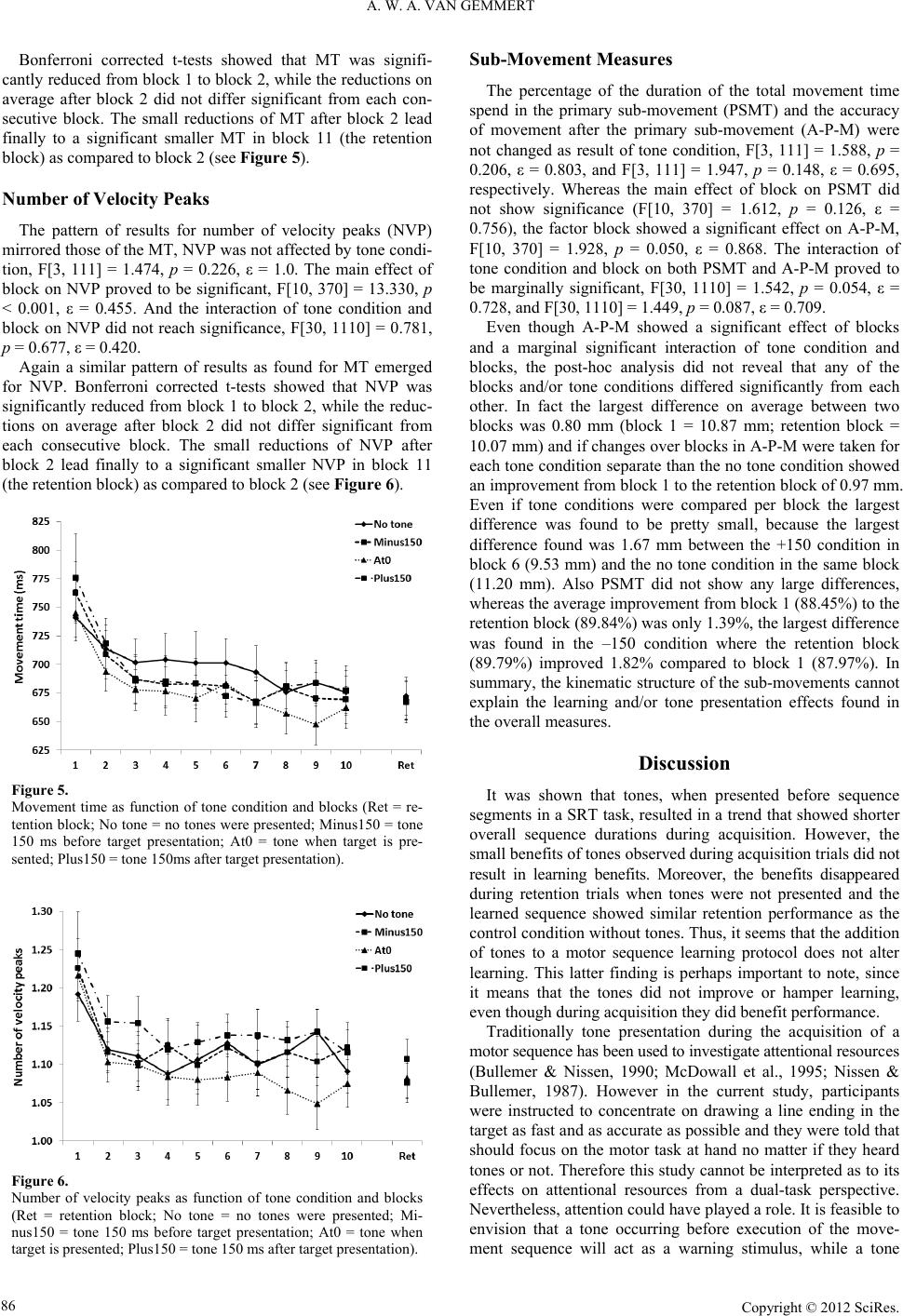
Whereas reaction time was affected by tones during acquisi-
tion, movement speed, smoothness, and execution efficiency
were not altered by the tones as indicated by the findings that
tone condition did not show main effects for movement time,
number of velocity peaks or any of the sub-movement time meas-
ures. Therefore, it can be concluded that only transient prepara-
tory processes which are only involved in movement initiation
related activities benefit of the tones. When assumed that the
task is easy or becomes very fast easy with practice, the results
that the tones benefit preparatory processes is in line with the
theoretical perspective that tones increase non-specific activa-
tion in the motor system, which benefits simple aiming tasks
(Van Gemmert & Van Galen, 1998). Another possibility for
improvements in reaction time during acquisition is that the
organization of the trials was very consistent, and it has been
suggested that learning depends on practicing run of trials that
are consistently organized (Stadler, 1995). However, this ex-
planation has difficulty to explain the finding that improve-
ments in reaction time do disappear during retention when the
trials do not include a tone. Moreover, since the trials during
acquisition include tones and the trials during retention do not
include tones, it is expected that reaction time would be worse
than retention for the no tone condition, because the latter con-
dition has consistency of organization across acquisition and
etention, while the experimental conditions are not consistent
in organization across acquisition and retention.
r
In summary, the current study showed evidence that tones
affect acquisition performance depending on the time of pres-
entation, however, it does not alter learning. More specifically,
the tones prove to be performance variables that do not alter the
execution phase of performance, but it has a direct impact on
reaction time. This pattern together with the finding that tones
do not alter learning supports the hypothesis that these tones
result in a non-specific activation of the motor system (Van
Gemmert & Van Galen, 1997).
REFERENCES
Bullemer, P., & Nissen, M. J. (1990). Attentional orienting in the ex-
pression of procedural knowledge. Bulletin of the Psychonomic Soci-
ety, 28, 505-505.
Cohen, A., Ivry, R. I., & Keele, S. W. (1990). Attention and structure in
sequence learning. Journal of Experimental Psychology-Learning Me-
mory and Cognition, 16, 17-30. doi:10.1037/0278-7393.16.1.17
Ketcham, C. J., Seidler, R. D., Van Gemmert, A. W., & Stelmach, G. E.
(2002). Age-related kinematic differences as influenced by task dif-
ficulty, targetsize, and movement amplitude. The Journals of Ger-
ontology: Series B , 57 , 54-64. doi:10.1093/geronb/57.1.P54
McDowall, J., Lustig, A., & Parkin, G. (1995). Indirect learning of
event sequences: The effects of divided attention and stimulus con-
tinuity. Canadian Journal of Experimental Psychology-Revue Cana-
dienne De Psychologie Experimentale, 49, 415-436.
doi:10.1037/1196-1961.49.4.415
Meulenbroek, R. G., & Van Galen, G. P. (1988). Foreperiod duration
and the analysis of motor stages in a line-drawing task. Acta Psy-
chologica, 69, 19-34. doi:10.1016/0001-6918(88)90027-3
Nissen, M. J., & Bullemer, P. (1987). Attentional requirements of learn-
ing—Evidence from performance-measures. Cognitive Psychology,
19, 1-32. doi:10.1016/0010-0285(87)90002-8
Richard, M. V., Clegg, B. A., & Seger, C. A. (2009). Implicit motor
sequence learning is not represented purely in response locations.
Quarterly Journal of Experimental Psychology, 62, 1516-1522.
doi:10.1080/17470210902732130
Romero, D. H., Van Gemmert, A. W., Adler, C. H., Bekkering, H., &
Stelmach, G. E. (2003). Time delays prior to movement alter the
drawing kinematics of elderly adults. Human Movement Science, 22,
207-220. doi:10.1016/S0167-9457(02)00160-4
Schumacher, E. H., & Schwarb, H. (2009). Parallel response selection
disrupts sequence learning under dual-task conditions. Journal of
Experimental Psychology-General, 138, 270-290.
doi:10.1037/a0015378
Song, S., Howard, J. H., & Howard, D. V. (2008). Perceptual sequence
learning in a serial reaction time task. Experimental Brain Research,
189, 145-158. doi:10.1007/s00221-008-1411-z
Stadler, M. A. (1995). Role of attention implicit learning. Journal of
Experimental Psychology-Learning Memory and Cognition, 2, 674-
685. doi:10.1037/0278-7393.21.3.674
Tucha, O., Mecklinger, L., Walitza, S., & Lange, K. W. (2006). Atten-
tion and movement execution during handwriting. Human Movement
Science, 25, 536-552. doi:10.1016/j.humov.2006.06.002
Van Gemmert, A. W., & Van Galen, G. P. (1994). Effects of a second-
dary, auditory task on graphic aiming movements. In C. Faure, P.
Keuss, G. Lorette, & A. Vinter (Eds.), Advances in handwriting and
drawing: A multidisciplinary approach (pp. 421-439). Paris: Europia.
Van Gemmert, A. W., & Van Galen, G. P. (1997). Stress, neuromotor
noise, and human performance: A theoretical perspective. The Jour-
nal of Experimental Psychology: Human Perception and Perform-
ance, 23, 1299-1313. doi:10.1037/0096-1523.23.5.1299
Van Gemmert, A. W., & Van Galen, G. P. (1998). Auditory stress
effects on preparation and execution of graphical aiming: A test of
the neuromotor noise concept. Acta Psychologica, 98, 81-101.
doi:10.1016/S0001-6918(97)00049-8