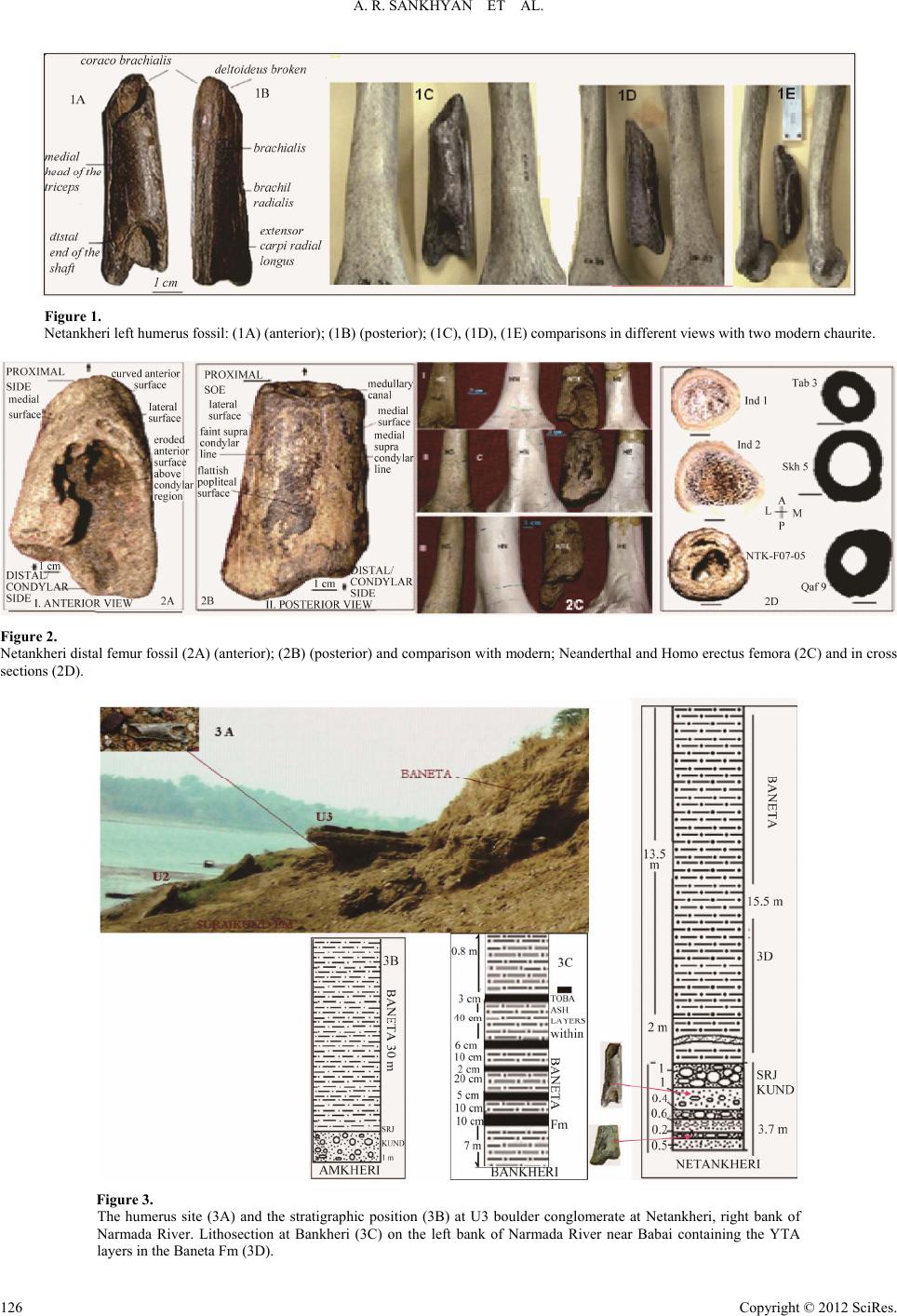
A. R. SANKHYAN ET AL.
luvium. Quaternary Research, 40, 10-19.
doi:10.1006/qres.1993.1051
Angel, J. L. (1972). A middle Paleolithic temporal bone from Darra-i-
Kur, Afghanistan. In L. Dupree (Ed.), Prehistoric research in Af-
ghanistan (1959-1966). Transactions of American Philosophical So-
ciety, 62, 54-56.
Athreya, S. (2007). Was Homo heidelbergensis in South Asia? A test
using the Narmada fossil from central India. In M. D. Petraglia, & B.
Allchin (Eds.), The evolution and history of human populations in
South Asia (pp. 137-170). New York: Springer Press.
doi:10.1007/1-4020-5562-5_7
Badam, G. L., Ganjoo, R. K., Salahuddin, M., & Rajaguru, S. N. (1986).
Evaluation of fossil hominin—The maker of Late Acheulean tools at
Hathnora, Madhya Pradesh, India. Current Science, 55, 143-145.
Badam, G. L., & Sankhyan, A. R. (2009). Evolutionary trends in Nar-
mada fossil fauna. In A. R. Sankhyan (Ed.), Asian perspectives on
human evolution (pp. 92-102). New Delhi: Serials Publications.
Barik, S. S., Sahani, R., Prasad, B. V. R., Endicott, P. et al. (2008). De-
tailed mtDNA genotypes permit a reassessment of the settlement and
population structure of the Andaman Islands. American Journal of
Physical Anthropology, 136, 19-27. doi:10.1002/ajpa.20773
Bermudez de Castro J. M., Carretero, J. M., Garcıa-Gonzalez R., Rodri-
guez-Garcıa, L., Martinon-Torres, M., Rosell, J., Blasco, R., Martın-
Frances, L., Modesto, M., & Carbonell, E. (2012). Early pleistocene
human humeri from the gran dolina-TD6 site (Sierra de Atapuerca,
Spain). American Journal of Physical Anthropology, 147, 604-617.
doi:10.1002/ajpa.22020
Cameron, D., Patnaik, R., & Sahni, A. (2004). The phylogenetic sig-
nificance of the Middle Pleistocene Narmada hominin cranium from
central India. International Journal of Osteoarchaeology, 14, 419-
447. doi:10.1002/oa.725
Carretero, J. M., Haile-Selassie, Y., Rodríguez, R., & Arsuaga, J. L.
(2009). A partial distal humerus from the Middle Pleistocene depos-
its at Bodo, middle awash, Ethiopia. Anthropological Science, 117,
19-31. doi:10.1537/ase.070413
Chandrasekar, A., Kumar, S., Sreenath, J. et al. (2009). Updating phy-
logeny of mitochondrial DNA macrohaplogroup M in India: Disper-
sal of modern human in South Asian Corridor. PLoS ONE, 4, 1-13.
doi:10.1371/journal.pone.0007447
Chesner, C. A., Rose, W. I., Drake, A. D. R., & Westgate, J. A. (1991).
Eruptive history of earth’s largest quaternary caldera (Toba, Indone-
sia) clarified. Geology, 19, 200-203.
doi:10.1130/0091-7613(1991)019<0200:EHOESL>2.3.CO;2
Churchill, S. E. (2007). Endocrine models of skeletal robusticity and
the origins of gracility. In A. R. Sankhyan, & V. R. Rao (Eds.), Hu-
man origins, genome and people of India (pp. 337-368). New Delhi:
Allied Publishers.
Ganjoo, R. K., Rajaguru, S. N., & Gupta, A. (1996). On the problem of
age and genesis of Bhedaghat waterfalls (Jabalpur), Madhya Pradesh,
Journal Geological Society of India, 48, 421-425.
Van Heteren, A. H., & Sankhyan, A. R. (2009). Hobbits and pygmies:
Trends in evolution. In A. R. Sankhyan (Ed.), Asian perspectives on
human evolution (pp. 172-187). New Delhi: Serials Publications.
James, H. V. A., & Petraglia, M. D. (2005). Modern human origins and
the evolution of behavior in the later pleistocene record of South
Asia. Current Anthropology, 46, 3-27. doi:10.1086/444365
Kennedy, K. A. R. (2000). God-apes and fossil men: The paleoanthro-
pology of South Asia. Michigan: The University of Michigan Press.
Kennedy, K. A. R. (2007). The Narmada fossil hominid. In A. R.
Sankhyan, & V. R. Rao (Eds.), Human origins, genome and people
of India (pp. 188-192). New Delhi: Allied Publishers.
Kennedy, K. A. R., & Deraniyagala, S. U. (1989). Fossil remains of
28,000 year old hominids from Sri Lanka. Current Anthropology, 30,
394-399. doi:10.1086/203757
Kennedy, K. A. R., Sonakia, A., Chiment, J., & Verma, K. K. (1991). Is
the Narmada hominin an Indian Homo erectus? American Journal of
Physical Anthropology, 86, 475-496. doi:10.1002/ajpa.1330860404
Khan, A., & Sonakia, A. (1992). Quaternary deposits of Narmada with
special reference to the hominid fossil. Journal of the Geological
Society of India, 39, 147-154.
De Lumley, H., & Sonakia, A. (1985). Contexte stratigraphique et
Archéologique de L’Homme de le Narmada, Hathnora, Madhya
Pradesh, Inde. L’Anthropologie, 89, 3-12.
Lumley, M. A., & Sonakia, A. (1985). Premiere Découverte D’un
Homo erectus Sur Le Continent Indien a Hathnora, Dans la Moyenne
vallée de la Narmada. L’Anthropologie, 89, 13-61.
Mallasse, A. D. (2009). Cranial embryogeny and hominin phylogeny.
In A. R. Sankhyan (Ed.), Asian perspectives on human evolution (pp.
103-121). New Delhi: Serials Publications.
Oppenheimer, C. (2002). Limited global change due to largest known
Quaternary eruption, Toba ~74 Kyr BP. Quaternary Science Review,
21, 1593-1609. doi:10.1016/S0277-3791(01)00154-8
Oppenheimer, C. (2003). Ice core and palaeoclimatic evidence for the
great volcanic eruption of 1257. International Journal of Clima-
tology, 23, 417-426. doi:10.1002/joc.891
Patnaik, R., Chauhan, P. R., Rao, M. R., Blackwell, B. A. B., Skinner,
A. R., Sahni, A., Chauhan, M. S., & Khan, H. S. (2009). New geochro-
nological, paleoclimatological and Paleolithic data from the Narmada
Valley hominin locality, Central India. Journal of Human Evolution,
56, 114-133. doi:10.1016/j.jhevol.2008.08.023
Pearson, O. M. (2000). Activity, climate and postcranial robusticity:
Implications for modern human origins and scenarios of adaptive
change. Current Anthropology , 41, 569-605. doi:10.1086/317382
Petraglia, M. D., & Alsharekh, A. (2003). The middle paleolithic of
Arabia: Implications for modern human origins, behaviour and dis-
persals. Research (Online).
Petraglia, M. D. (2007). Middle paleolithic assemblages from the In-
dian Subcontinent before and after the Toba Super-Eruption. Sci-
ence, 317, 114-116. doi:10.1126/science.1141564
Rose, W. I., & Chesner, C. A. (1990). Worldwide dispersal of ash and
gases from earth’s largest known eruption: Toba, Sumatra, 75 Ka.
Palaeogeography, Pala e o - C l i m a to l o g y , P a la e o e c o l o g y, 89, 269-275.
doi:10.1016/0031-0182(90)90068-I
Sankhyan, A. R. (1997a). Fossil clavicle of a Middle Pleistocene homi-
nid from the Central Narmada Valley, India. Journal of Human Evo-
lution, 32, 3-16. doi:10.1006/jhev.1996.0117
Sankhyan, A. R. (1997b). A new human fossil find from the Central
Narmada basin and its chronology. Current Science, 73, 1110-1111.
Sankhyan, A. R. (2005). New fossils of Early Stone Age man from
Central Narmada Valley. Current Science, 88, 704-707.
Sankhyan, A. R. (1999). The place of Narmada hominin in the Jigsaw
puzzle of human origins. Gondwana Geological Magazine Special
Publication, 4, 335-345.
Sankhyan, A. R. (2006). On the status of Indian hominoid and hominid
fossils. In R. Ray, & V. Jayaswal (Eds.), Status of Prehistoric Studies
in the 21st Century in India, Proceedings of 15th UISPP Congress,
Lisbon, BAR International Series 1924 (pp. 13-23), Oxford: Archaeo
Press.
Sankhyan, A. R. (2010). Pleistocene Hominins & associated findings
from central Narmada Valley bearing on the evolution of man in
South Asia. Ph.D. Thesis, Chandigarh: Panjab University.
Sankhyan, A. R. & Rao, V. R. (2007). Did ancestors of the Pygmy or
Hobbit ever live in Indian heartland? In E. Indriati (Ed.), Recent ad-
vances on Southeast Asian paleoanthropology and archeology (pp.
76-89). Yogyakarta: Gadjah Mada University.
Sankhyan, A. R., Dewangan, L. N., Sahoo, R. H., Chakravarty, R., &
Chatterjee, R. (2011). Early prehistoric signatures of man in Bastar
region, Central India. Current Science, 101, 1146-1149.
Sonakia, A. (1984). The skullcap of Early Man and associated mam-
malian fauna from Narmada Valley Alluvium, Hoshangabad area,
M.P. (India). Records Geological Survey of India, 113, 159-172.
Tiwari, M. P., & Bhai H. Y. (1997). Quaternary stratigraphy of the
Narmada Valley. Geological Survey of India Special Publication, 46,
33-63.
Todd, R. Y., & Churchill, S. E. (2006). Archaic and modern human
distal humeral morphology. Journal of Human Evolution, 51, 603-
616. doi:10.1016/j.jhevol.2006.07.006
Trinkaus, E. (2007). Appendicular robusticity and the paleobiology of
modern human emergence. Proceedings of the National Academy of
Sciences of the United States of America, 94, 13367-13373.
doi:10.1073/pnas.94.24.13367
Trinkaus, E., & Ruff, C. B. (1999). Diaphyseal cross-sectional Geome-
Copyright © 2012 SciRes.
130