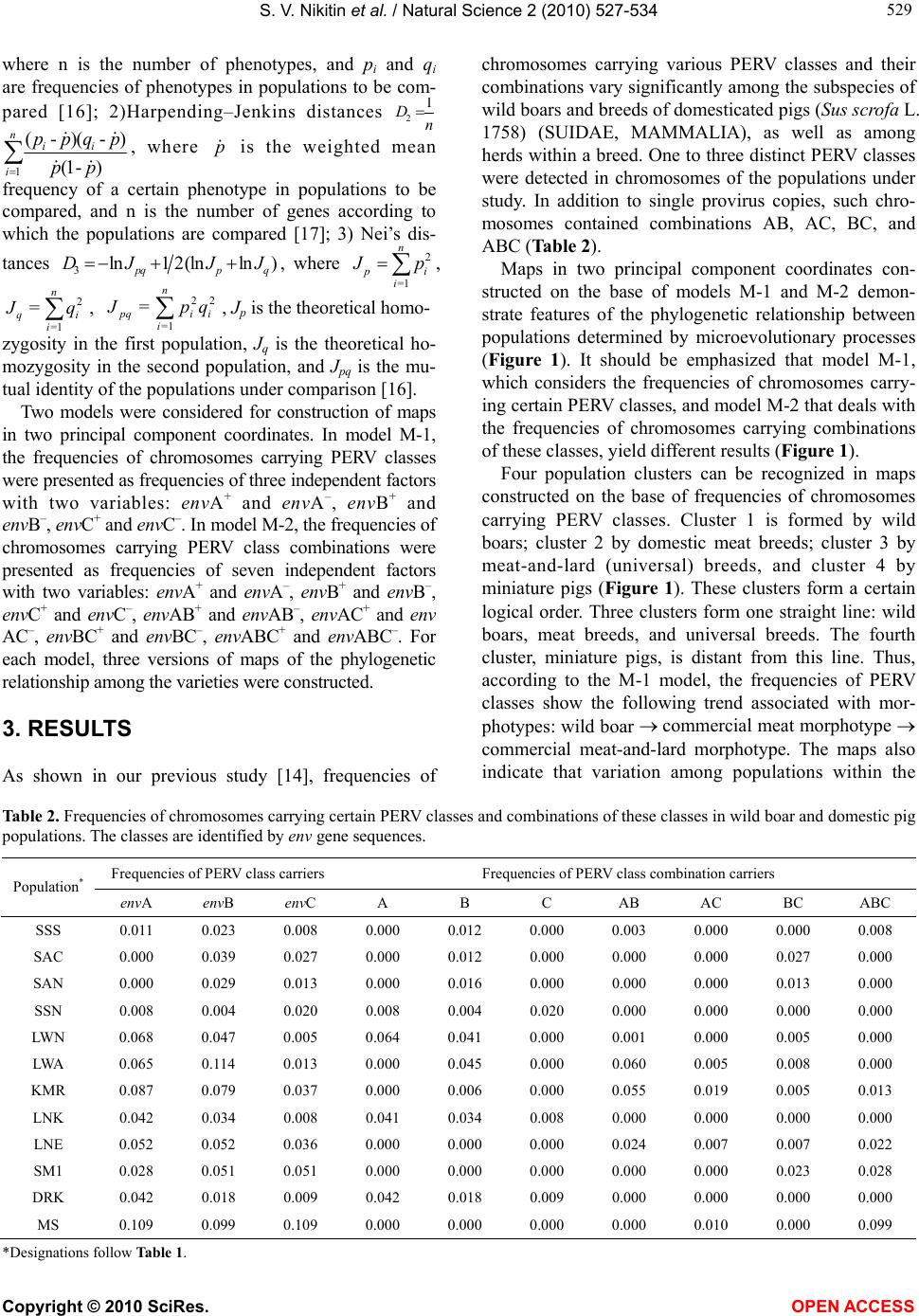
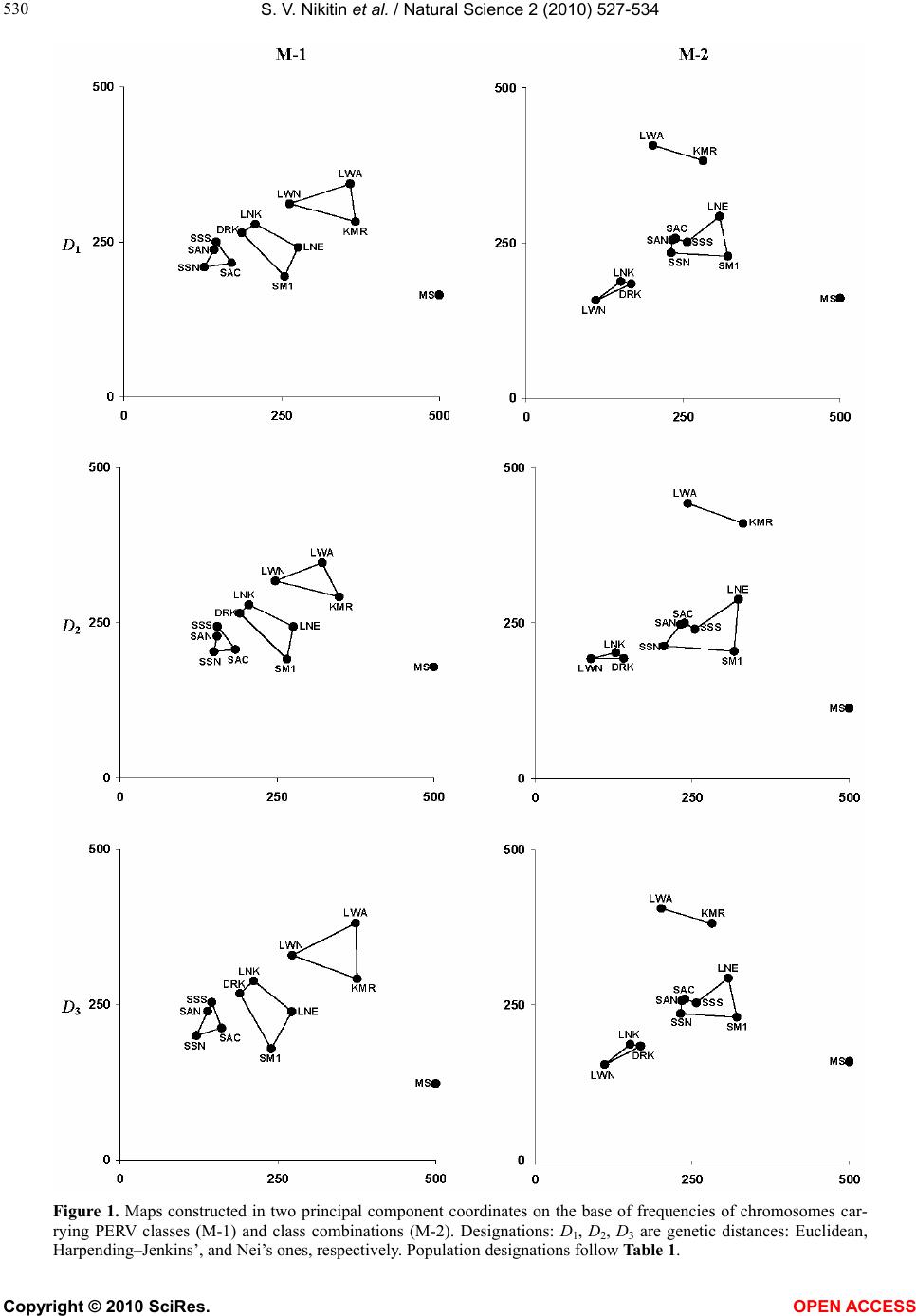
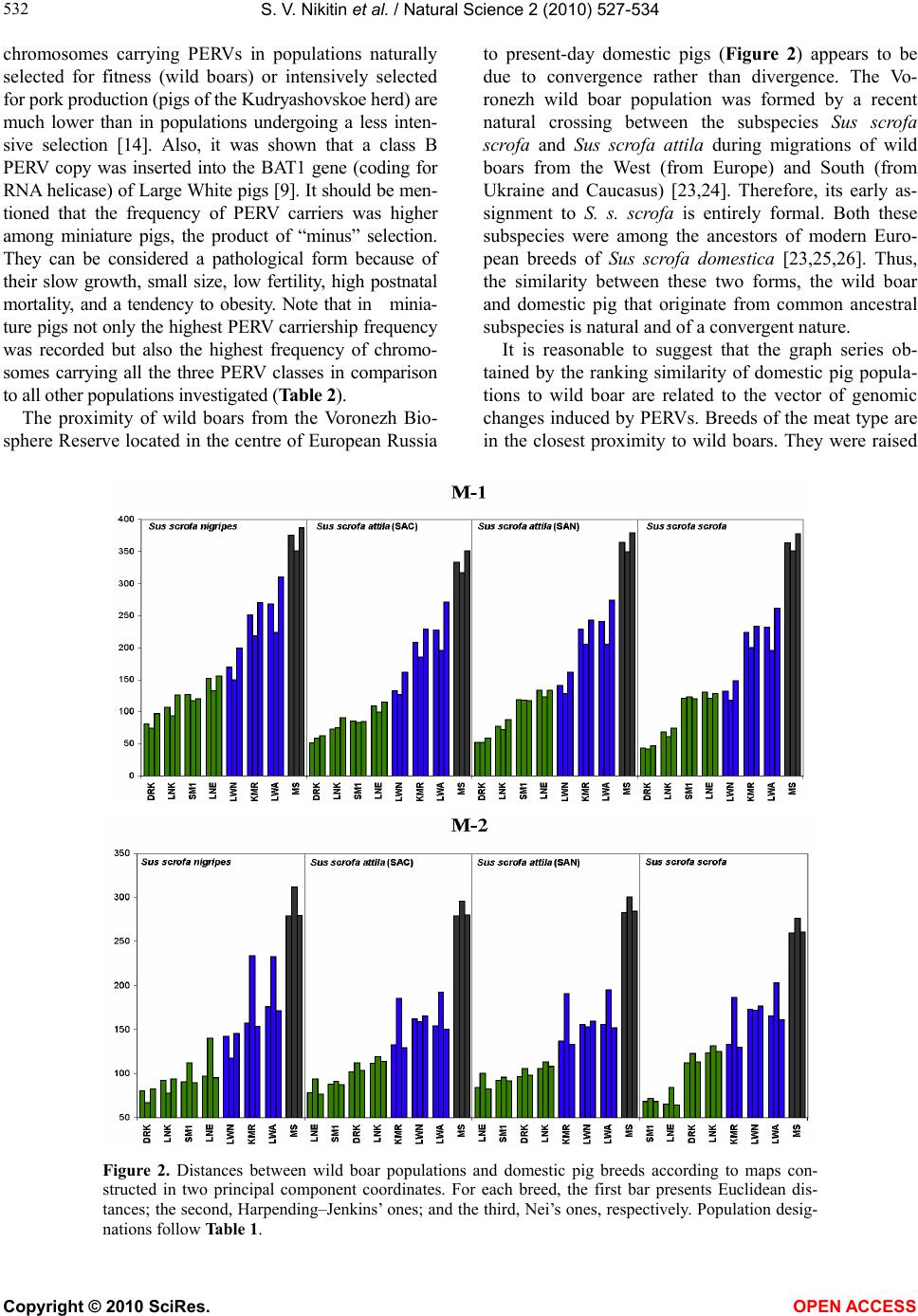
S. V. Nikitin et al. / Natural Science 2 (2010) 527-534
Copyright © 2010 SciRes. OPEN ACCESS
533
533
by selection for a less intense fat deposition. The ele-
vated fat deposition in domestic pigs in comparison with
wild or early domesticated forms is an obvious abnor-
mality, which may have been caused by the breakdown
of some genes owing to PERV insertion. Therefore, the
natural selection against these breaks favoured alleles
characteristic of the original wild boar or similar. This
may have resulted in convergent similarity between meat
pig breeds and wild boars. Domestic breeds of the uni-
versal meat-and-lard morphotype should possess a cer-
tain number of loci in the genome that would determine
the fat deposition degree corresponding to this morpho-
type. Mutations in some of these loci caused by PERV
insertion give rise to the desirable trait; therefore, this
morphotype diverges more from wild boars than breeds
of the meat type in the frequencies of chromosomes car-
rying certain PERV classes and type combinations.
Miniature pigs were raised by intensive selection for a
smaller adult body size with minimum selection for
other traits. The development of irrelevant traits should
be sufficient for no more than maintenance of the popu-
lation. For this reason, it is likely that the miniature pig
genome was enriched in loci whose function was dis-
rupted by PERV insertions. In some cases, this favoured
the desired trait (small size), and in other cases this was
of no significance, because no selection for commercial
traits was conducted.
In summary, we analyzed in this study patterns of dif-
ferentiation of domestic and wild pigs in the frequencies
of chromosomes carrying certain PERV classes and type
combinations. With regard to this differentiation, we
demonstrated that the convergence processes were at
least no less significant than the divergence ones. It ap-
pears that PERVs were not neutral elements in the evo-
lution of the pig genome.
5. ACKNOWLEDGEMENTS
This study was supported by the Gene pool dynamics project of the
Russ. Acad. Sci Presidium program Biodiversity and gene pool dy-
namics.
REFERENCES
[1] Niebert, M. and Tonjes, R.R. (2005) Evolutionary spread
and recombination of porcine endogenous retroviruses in
suiformes. Journal of Virology, 79(1), 649-654.
[2] Akiyoshi, D.E., Denaro, M., Zhu, H., et al. (1998) Iden-
tification of a full-length cDNA for an endogenous retro-
virus of miniature swine. Journal of Virology, 72(5), 4503-
4507.
[3] Le Tissier, P., Stoye, J. P., Takeuchi, Y., et al. (1997) Two
sets of human-tropic pig retrovirus. Nature, 389(6652),
681-682.
[4] Takeuchi, Y., Patience, C., Magre, S., et al. (1998) Host
range and interference studies of three classes of pig en-
dogenous retrovirus. Journal of Virology, 72(12), 9986-
9991.
[5] Bosch, S., Arnauld, C. and Jestin, A. (2000) Study of
full-length porcine endogenous retrovirus genomes with
envelope gene polymorphism in a specific-pathogen-free
large white swine herd. Journal of Virology, 74(18),
8575-8581.
[6] Herring, C., Quinn, G., Bower, R., Parsons, N., Logan, N.
A., Brawley, A., Elsome, K., Whittam, A., Fernandez-
Suarez, X.M., Cunningham, D., Onions, D., Langford, G.
and Scobie, L. (2001) Mapping full-length porcine en-
dogenous retroviruses in a large white pig. Journal of
Virology, 75(24), 12252-12265.
[7] Rogel-Gaillard, C., Bourgeaux, N., Billault, A., Vaiman,
M. and Chardon, P. (1999) Construction of a swine BAC
library: Application to the characterization and mapping
of porcine type C endoviral elements. Cytogenetics and
Cell Genetics, 85(3-4), 205-211.
[8] Lee, J.H., Webb, G.C., Allen, R.D. and Moran, C. (2002)
Characterizing and mapping porcine endogenous retrovi-
ruses in Westran pigs. Journal of Virology, 76(11), 5548-
5556.
[9] Niebert, M. and Tönjes, R.R. (2003) Analyses of preva-
lence and polymorphisms of six replication-competent
and chromosomally assigned porcine endogenous retro-
viruses in individual pigs and pig subspecies. Virology,
313(2), 427-434.
[10] Edamura, K., Nasu, K., Iwami, Y., Nishimura, R., Ogawa,
H., Sasaki, N. and Ohgawara, H. (2004) Prevalence of
porcine endogenous retrovirus in domestic pigs in Japan
and its potential infection in dogs xenotransplanted with
porcine pancreatic islet cells. Journal of Veterinary
Medical Science, 66(2), 129-135.
[11] Jin, H., Inoshima, Y., Wu, D., et al. (2000) Expression of
porcine endogenous retrovirus in peripheral blood leuco-
cytes ten different breeds. Transplant Infectious Disease,
2(1), 11-14.
[12] Zhang, L., Yu, P., Li, S.F., Bu, H., Li, Y.P., Zeng, Y.Z. and
Cheng, J.Q. (2004) Phylogenetic relationship of porcine
endogenous retrovirus (PERV) in Chinese pigs with
some type C retrovirus. Virus Research, 105(2), 167-173.
[13] Aitnazarov, R.B., Ermolaev, V.I., Nikitin, S.V., et al.
(2006) Associations between various endogenous virus
types and genetic markers in domestic and wild pig
populations. Russian Agricultural Sciences (Doklady
Rossiiskoi Akademii Sel’skokhozyaistvennykh Nauk), 4,
39-43.
[14] Nikitin, S.V., Yudin, N.S., Knyazev, S.P., et al. (2008)
Frequency of chromosomes carrying endogenous retro-
viruses in the populations of domestic pig and wild boar.
Russian Journal of Genetics, 44(6), 686-693.
[15] Zhivotovskii, L.A. (1991) Populational biometry. in
Russian, Nauka, Moscow.
[16] Weir, B.S. (1995) Genetic data analysis. in Russian, MIR,
Moscow.
[17] Harpending, H.C. and Jenkins, T. (1973) Genetic dis-
tances among Southern African Populations. In: Methods
and Theories of Anthropological Genetics. University of
New Mexico Press, Albuquerque, 177-199.
[18] Porter, V. (1993) Pigs: A handbook to the breeds of the