Journal of Biophysical Chemistry
Vol.1 No.2(2010), Article ID:2539,9 pages DOI:10.4236/jbpc.2010.12010
Distance-dependent coherent charge transport in DNA: crossover from tunneling to free propagation*
![]()
1Department of Microand Nanoelectronics, Belarusian State University of Informatics and Radioelectronics, Minsk, Belarus; #Corresponding Author: ngrib@uvic.ca
2Institute for Theoretical Physics, University of Regensburg, Regensburg, Germany
3Institute for Material Science and Max Bergmann Center for Biomaterials, Dresden University of Technology, Dresden, Germany
Received 8 June 2010; revised 4 July 2010; accepted 10 July 2010.
Keywords: DNA; Electron Transport; Modeling of DNA; Electronic Structure of DNA
ABSTRACT
Using a tight-binding model, we investigate the influence of intraand interstand coupling parameters on the charge transport properties in a G-(T)j-GGG DNA sequence and its (G:C)-(T:A)j- (G:C)3 duplex attached to four electrodes. Dependences of the transmission function and of the corresponding conductance of the system on the number of bridging sites were obtained. Simulation results of a recently proposed two- strand superexchange (tunneling) model were reproduced and extended. It is demonstrated that the crossover from strong to weak distance-dependent charge transport is elucidated by a transition from under-barrier tunneling mechanism to free over-barrier propagation in the coherent regime, controlled by temperature and coupling parameters. The role of DNAelectrode coupling has been also considered. It was found that an asymmetry in the DNA-electrode coupling has a drastic effect on the conductance leading to an increase in delocalization of the electronic states in the DNA duplex.
1. INTRODUCTION
The discovery of charge migration in deoxyribonucleic acid (DNA) stimulated intensive investigations of the electronic properties of DNA due to their significance in biosynthesis and radiation-induced damage and repair processes [1-3]. Furthermore, considerable interest in nanodimensional structures of DNA possessing unique self-assembling and self-recognition properties has increased the last decade in connection with the possibility of the development of molecular nanoelectronic devices which are expected to provide high storage of information and high-speed signal processing within a wide temperature range [4-6]. In fact, DNA molecules can be well combined with silicon technology transcending the potential of the present quantum wires and are supposed to be used in modern computer technology as a binary data structure by applying a programmable linear selfassembly of the sequence of complementary nucleic base pairs of DNA [7,8].
Until now numerous experimental and theoretical data on charge migration through DNA molecules show an apparently contradictory behavior which can be elucidated by supposing two primary mechanisms. They include the single-step superexchange (tunneling) charge transfer that is strongly dependent on length of a molecular chain, and the multi-step hopping mechanism that is characterized by a weak change in the charge transfer efficiency (CTE) with increasing of the donoracceptor distance in the double helix [9-11]. However, experimental measurements of DNA molecule do not give any unequivocal evidence in favor of one or other mechanism of charge transfer in DNA. Many of them demonstrate a combined hopping-superexchange mechanism with a transition from the coherent superexchange to the thermally induced incoherent hopping process.
Beside electron transfer experiments, also transport experiments became an important field of modern research. In transport measurements a molecule is placed between metallic leads and steady-state current can be produced by finite voltage. A direct measurement of electrical transport through single biological molecules, such as DNA and peptides [12,13], is a very appealing, although challenging, issue in molecular electronics because of the potential peculiar capabilities of forming selfassembled nanodevices at the molecular scale. Quantum transport experiments through single DNA oligomers can be performed in both molecular junction configurations [14-17] and STM setups [18]. These enable the investigation of charge migration in both longitudinal and transverse configurations and stimulate theoretical interpretations.
Theoretically, single-step superexchange and multistep hopping models have been considered within various approaches such as variable range hopping [19,20], one-dimensional quantum mechanical tight-binding models [21-24], and nonlinear methods [25,26]. Recently, Wang and Chakraborty [27] have proposed a ladder model to describe charge transfer via DNA bridges and applied this model to the (G:C)-(T-A)n-(G:C)3 sequence studied experimentally by Giese et al. [28]. There, the presence of both transfer mechanisms was obtained by analyzing strand cleavage efficiencies that are caused by oxidized guanines. It has been shown that the superexchange mechanism exponentially decreased with distance and predominated for short bridges (n ≤ 3) consisting of A:T base pairs. For long bridges (n > 3), the CTE showed an almost distance-independent behavior. This distance dependence was further proved experimentally by Lewis and co-workers [29], who clarified the rates of photoinduced charge transfer in short DNA hairpins. The theoretical analysis in [27], which addresses the conductance of the same sequence rather than the CTE, rests on Landauer theory and implicitly assumes purely coherent charge propagation upon injection. As shown by the authors, by tuning the inter-strand coupling the experimentally observed [28,29] length dependence vs. n can be reproduced. Wang and Chakraborty interpreted the crossover between strong and weak length dependencies as a transition to a quasitwo dimensional behavior characterized by a topological effect. While there is a consensus that the exponential decrease of CTE with number of A:T base pair for short bridges is related to a superexchange tunneling process between the distant G-bases, the saturation of CTE for large n seems more difficult to understand.
In this paper, we consider a ladder model within a nearest-neighbor tight-binding picture to describe the distance dependent charge transport in DNA. We introduce the model Hamiltonian and discuss the transmission function which is required to calculate the conductance in DNA in order to reproduce and to extend the WangChakraborty model [27]. We show that different electronic coupling regimes of nucleobases in DNA within its singleand double-stranded configurations can determine the crossover point from exponential distance dependence of charge transfer to length-independent behavior.
2. MODEL
The DNA molecules under study are considered to consist of repeated stacks of nucleobases formed by either A-T/T-A or G-C/C-G pairs located along a sugar-phosphate frame, as shown schematically in Figure 1. Hydrogen bonds between basic and acid centers of the complementary bases and also their stacking interaction stabilize the spiral structure of the molecule and do not participate in the charge carrier transport. It has determined that a p - p orbital overlapping of neighboring nucleobases forms the basis of charge transfer mechanism in DNA [30] allowing the basic charge carriers (holes) to jump from one base to another with increasing their transfer rate for the sequence of identical nucleotides. Since the guanine has the lowest oxidation potential [31], holes are effectively trapped at the sites of guanines in the DNA duplex, where (5'-G) segment supposed to be a donor and (GGG-3') site performs an acceptor function (see red regions in Figure 1). We assume that charge carriers are moving along the base-pair stack in the direction of an applied electric field, thus neglecting the slow migration of holes in the opposite direction. In the model we do not take into account environmental effects and helical impact arising from the real structure of the DNA.
Along the lines of [21] and [32], we represent the DNA molecule with N-base-pair nucleic chains, attached to four semi-infinite electrodes by the following tightbinding Hamiltonian (Figure 1):
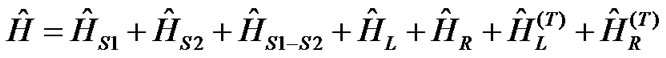 (1)
(1)
Hamiltonians for the first ĤS1, second ĤS2 DNA strands and the coupling between them ĤS1–S2 are described as:
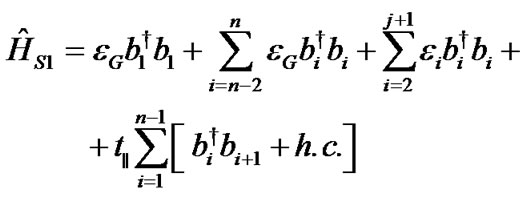 (2)
(2)
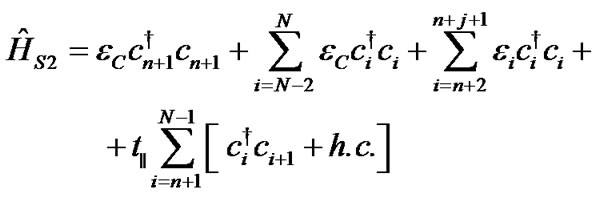 (3)
(3)
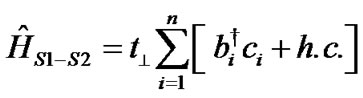 , (4)
, (4)
where bi†(bi) is the creation (annihilation) operator of hole on ith site in the first DNA strand (1 ≤ i ≤ n) and ci†(ci) is the same operator in the second molecular strand (n + 1 ≤ i ≤ N). The on-site energies εi are evaluated by ionization potentials of the respective nucleobases as the HOMO hole energies given in [10,33]: εG = 7.75 eV, εC = 8.87 eV, εT = 9.14 eV and εA = 8.24 eV. The coupling parameter t|| is the corresponding nearest-neighbor electronic hopping
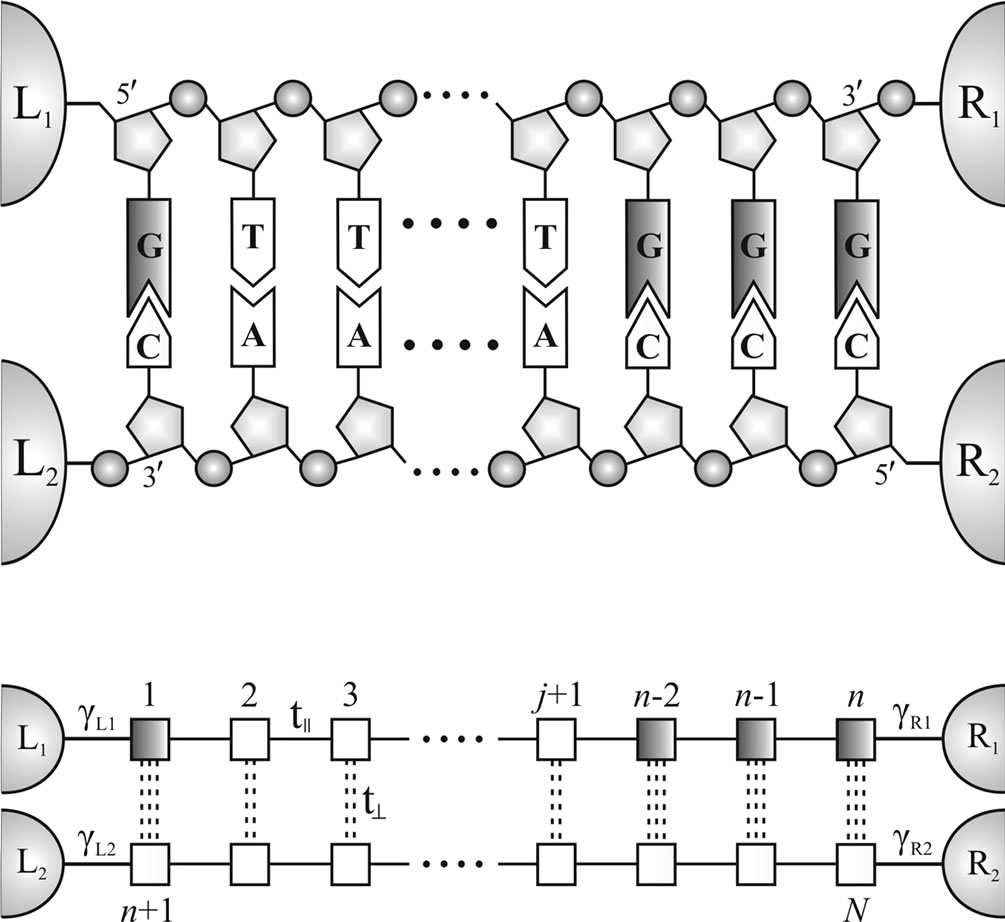
Figure 1. Upper panel: Schematic representation of the double-strand DNA attached to four semi-infinite left (L1,2) and right (R1,2) electrodes. A nucleobase-pair sequence with the donor (5'-G) and acceptor (GGG-3') segments is placed between two sugar-phosphate backbones. Lower panel: A two-leg ladder model of the DNA with the (G:C)-(T:A)j-(G:C)3 configuration used to imitate the double-strand structure of a DNA molecule. Charge carriers are moved along the 5'-G-(T)j-GGG-3' chain from the emitter electrode L1 to the collector electrode R1. 
integral along the two strands while t^ describes the inter-strand hopping.
The Hamiltonians of the left and right electrodes represented by the terms ĤL and ĤR in (1) and their contact Hamiltonians with the DNA duplex ĤL(T) and ĤL(T) are given as:
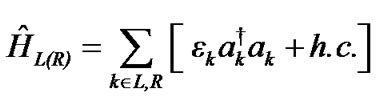 (5)
(5)
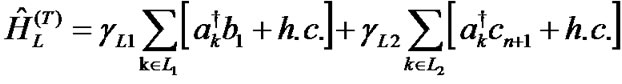 (6)
(6)
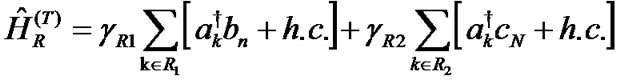 (7)
(7)
Here, εk is the energy of electrons in the leads, ak†(ak) is the creation (annihilation) operator of hole in the state k, γL1(L2) and γR1(R2) are the coupling terms to the left and right electrodes, respectively (Figure 1).
Studying the injected hole transport along the 5'-G- (T)j-GGG-3' DNA chain located between the emitter electrode L1 and the collector electrode R1 as in [27], the choice of the intraand inter-strand electronic transfer integrals is of considerable importance. The least change in their values can lead to quite different quantum transfer properties in DNA duplexes because the coupling parameters are rather sensitive to the relative position of two neighboring nucleobases due to their highly anisotropic p-bonds [34]. In order to reduce the number of model parameters and to simplify our computation we have adopted a simple parameterization taking a homogenous hopping along both legs in a range of 0.1-0.5 eV for the coupling parameter t|| and 0.01-0.07 eV for inter-strand hopping integral t^. While in [35] has been shown that hopping integral values are different for each combination of nucleotide pairs and the inter-strand hopping is usually very small, ~ few meV [36], we do not consider the hopping integrals as bare tight-binding parameters but as effective ones, thus keeping some freedom in the choice of their specific values.
To diminish contact effects on quantum tunneling-transport process in the DNA molecule, the contact strength with the left and the right DNA chain ends is kept constant and the coupling parameter γL(R) supposed to be much larger than the coupling parameter between the nucleobases in the DNA sequence [27]. We choose equal couplings to all electrodes (γL1 = γL2 = γR1 = γR2 = 1.5 eV) that provides a decrease in the influence of interference effects at the electrode-DNA interface [37] and allows to ascertain the veritable conduction properties of the DNA.
The Green’s function theory is used to calculate the transmission coefficient in the open system “electrodeDNA-electrode” by the expression
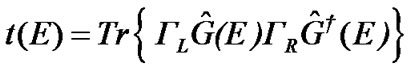 (8)
(8)
with the Green function,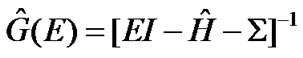 . Here the self-energy Σ = ΣL + ΣR is introduced due to the contacts of the DNA molecules to the left and the right metallic electrodes, and the corresponding coupling matrices are related to the self-energies as ΓL(R) = Im(ΣL(R) – Σ†L(R)). Then the N × N ladder-lead energy-independent coupling matrices within the wide-band limit in the electrodes spectral densities are given by
. Here the self-energy Σ = ΣL + ΣR is introduced due to the contacts of the DNA molecules to the left and the right metallic electrodes, and the corresponding coupling matrices are related to the self-energies as ΓL(R) = Im(ΣL(R) – Σ†L(R)). Then the N × N ladder-lead energy-independent coupling matrices within the wide-band limit in the electrodes spectral densities are given by
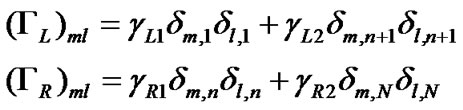 (9)
(9)
Finally, the conductance of the DNA molecule in the coherent regime is defined as
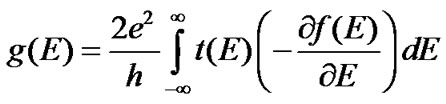 , (10)
, (10)
where f (E) = (exp[(E – EF)/kBT ] +1)-1 is the Fermi function at the room temperature T. As an essential physical factor controlling the injection efficiency of holes onto the stack of nucleobases is the local position of the electrode Fermi level EF relative to the unoccupied molecular orbitals, we assume the energy coincides with the on-site energy of guanine in the chain at the fixed voltage threshold. In this case the conductance of the DNA molecule starts to be governed by the ability of the base pair sequence to transport a charge rather than by the injection process.
3. SIMULATION RESULTS AND DISCUSSION
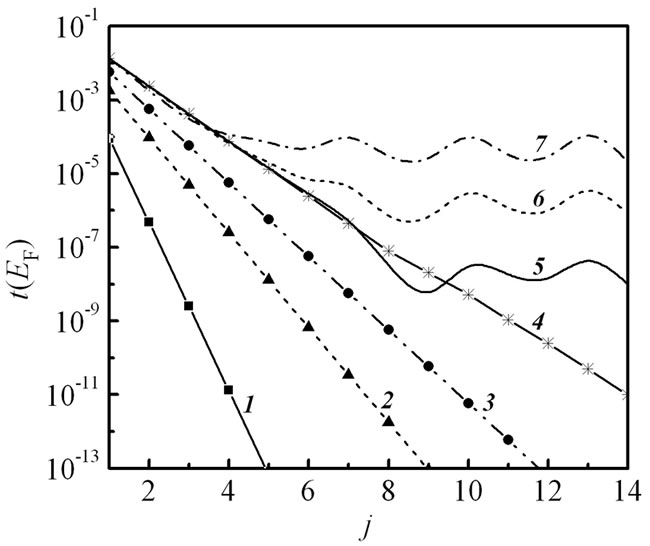
We evaluated the transmission coefficient t(EF) at the Fermi energy and the corresponding conductance g(E) as functions of number j (T:A) nucleobase pairs that compose the molecular bridge and determine the length of the (G:C)-(T:A)j-(G:C)3 DNA duplex (Figure 2). At first the calculations have been carried out for different values of intrastrand hopping integral t|| in the single-stranded 5'-G-(T)j-GGG-3' DNA sequence when t^= 0 (1-4 lines in Figure 2(a)). For this case the simulation results show a typical exponential distance-dependent behavior of single-step hole-transfer process in DNA with increasing T-bases as has been found for the current through the same DNA chain in [27]. A temperature contribution to a conduction mechanism of the one-leg DNA configuration
 (a)
(a)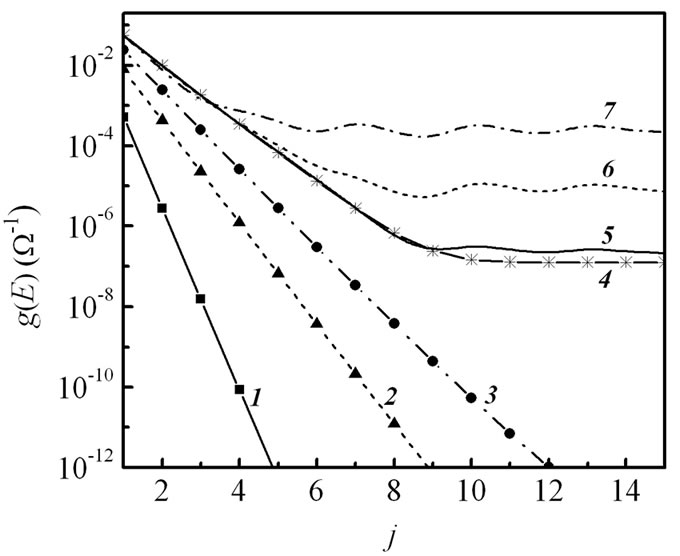 (b)
(b)
Figure 2. (a) Transmission coefficient t(EF) and (b) corresponding conductance g(E) vs. number of j (T:A) base pairs at T = 300 K for different intrastrand hopping integrals t|| (1 – 0.1 eV, 2 – 0.3 eV, 3 – 0.4 eV, 4 – 0.5 eV) in the single-strand structure of 5'-G-(T)j-GGG-3' DNA (equivalent to t^= 0) and for various values of the interstrand hopping t^ (5 – 0.01 eV, 6 – 0.03 eV, 7 – 0.07 eV) at fixed t|| = 0.5 eV in the double-stranded (G:C)-(T:A)j-(G:C)3 molecule. Coupling to the electrodes are γL1= γL2= γR1= γR2= 1.5 eV.
results in modification of the distance dependence when the overlapping of adjacent nucleobases becomes rather big. In Figure 2(b) it is seen a transition from the strong to the weak distance dependence of the conductance g(E) at the intrastrand hopping integral t|| = 0.5 eV (curve 4). The presence of such the crossover was obtained to appear only for the two-stranded DNA in the Wang-Chakraborty model [27]. The point is that the T-bridge forms an energy barrier for charge carries, propagating free through electronic states of G-bases, which have the on-site energies close to the Fermi energy in the electrodes (Figure 3, mechanism 1). This barrier is lower for longer bridges since the splitting between the bridge states of the identical bases is larger for longer bridge lengths leading to a
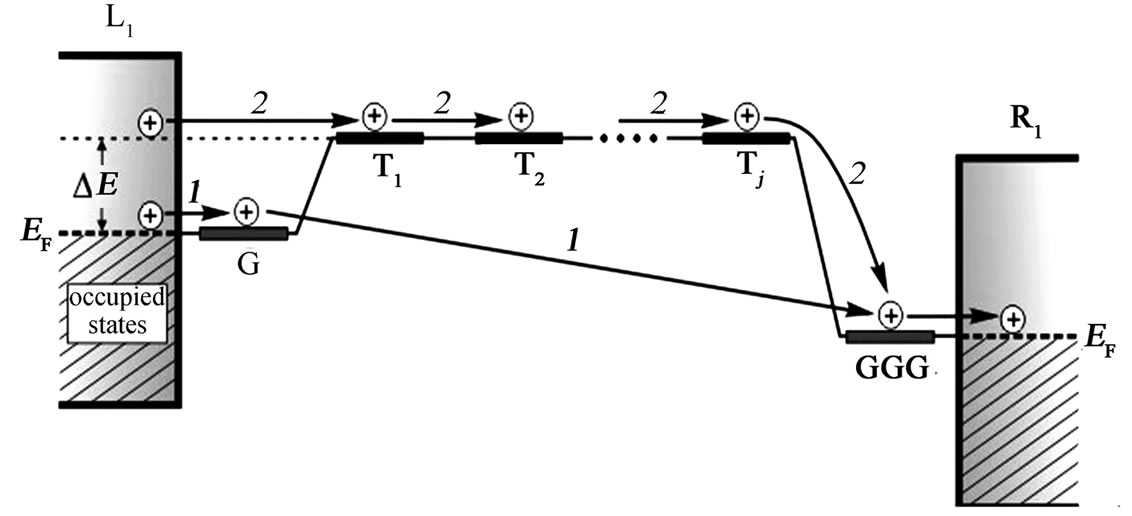
Figure 3. An approximate energetic scheme for hole transport in the 5'-G-(T)j-GGG-3' DNA strand: 1 – under-barrier tunneling mechanism between G-base and GGG-triplet connected to the metal leads; 2 – free over-barrier propagation along the T-bridge with a hole energy ΔE in the emitter electrode comparable with the energy level of T-nucleotide at finite temperature (see text).
strong overlap between orbitals in the linear T-chain [20]. At finite temperature the holes in the leads exist also at energies, sufficient for their capture by the first T-nucleotide in the bridge, and can coherently tunnel through the extended electronic states of T-bases resulting in distance-independent conductance while the number of the bridging sites rises (Figure 3, mechanism 2). The next curves (5-7) in Figure 2 were obtained for the double-stranded (G:C)-(T:A)j-(G:C)3 molecule at fixed t|| = 0.5 eV along both legs when the interstrand coupling parameter t^ is variable. An increase in the number of periodic nucleotides, which constitute the (T:A) molecular bridge, leads to a formation of an energy gap due to the high degree of their p-orbital overlap and the large interstrand coupling between them. In what follows, the hole migration through the set of (T:A) base pairs has a band-like behavior with preservation of hole energy during the transport process in the DNA. A crossover region is characterized by an appearance of oscillations for larger number of (T:A) dinucleotides when the overbarrier charge propagation through the (T:A) bridge preponderates over the under-barrier tunneling mechanism from the donor G-site to the acceptor GGG and shifts toward smaller j with increasing t^. However, it has a smoother behavior in comparison with the result of the work [27] and the experimental findings [28,38,39]. We suppose that these oscillations are to be a consequence of neglecting the environmental effects and the electronphonon interaction in the system, and smaller for the dependence of g(E) on j due to temperature effect.
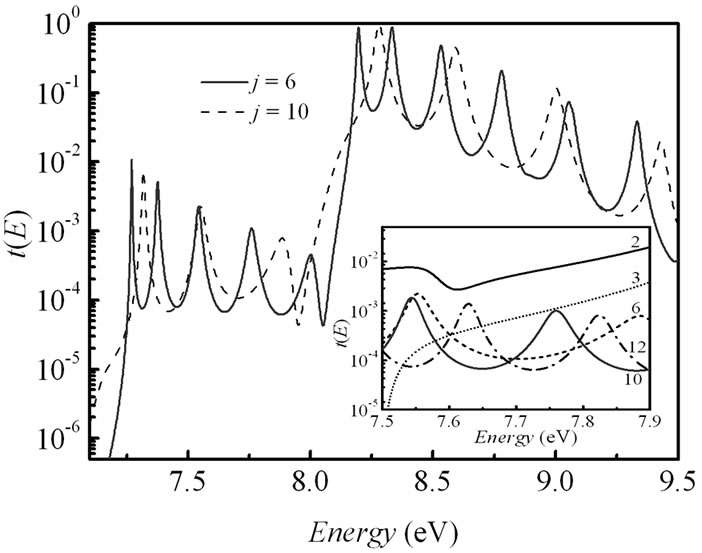
To reach a clearer understanding of the transport mechanisms we investigated the energy dependent transmission t(E) for different length of the molecular bridge in the 5'-G-(T)j-GGG-3' DNA configuration and its (G:C)-(T:A)j-(G:C)3 duplex. The results are displayed in Figure 4. The transmission spectrum consists of peaks related to the energetic states of the corresponding nucleobases, which take part in the hole transport. At the same time in the strong coupling regime (in this case t|| = 0.5 eV) some peaks are merged due to an increase in the hybridization between the p orbitals of the bases giving a broadening of the electronic manifolds. An inset in Figure 4(a) used to show the transmission t(E) around the Fermi energy for j = 2, 3, 6, 10 and 12. An extension of the bridge does not modify the charge propagation mechanism by the primary unistep model and just leads to exponential decreasing of the charge transfer rate in the 5'-G-(T)j-GGG-3' DNA strand. However, in the analogical inset of Figure 4(b) we observe that the t(E) picks have a steady spectral level for the (G:C)-(T:A)j-(G:C)3 duplex when j is more than 6, in that way demonstrating the crossover from the unistep transport through guanine sites to the coherent tunneling through the long (T:A) bridge. The obtained displacement of the picks can be explained by an energy alteration of the charge transport pathways [27].
In view of the fact that the hopping integral between the bases along the DNA chain has been determined not to exceed 0.4 eV [23,35,40], we calculated the transmission coefficient t(EF) (Figure 5) and the conductance g(E) (inset in Figure 5) for the (G:C)-(T:A)j-(G:C)3 DNA molecule with increased bridge length in a weak-coupling regime, viz. at t|| = 0.1 eV. As in Figure 2(b), the intrastrand hopping integral is variable. It was obtained that the magnitude of the g(E) at these conditions is noticeably less than the one in case of the stronger hopping coupling between the nucleobases, while the conductance in DNA molecules has been found to be higher in the recent stud-
 (a)
(a)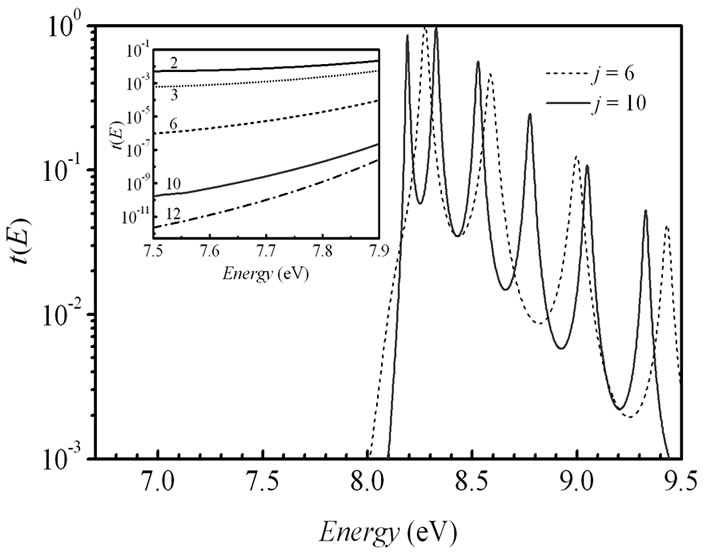 (b)
(b)
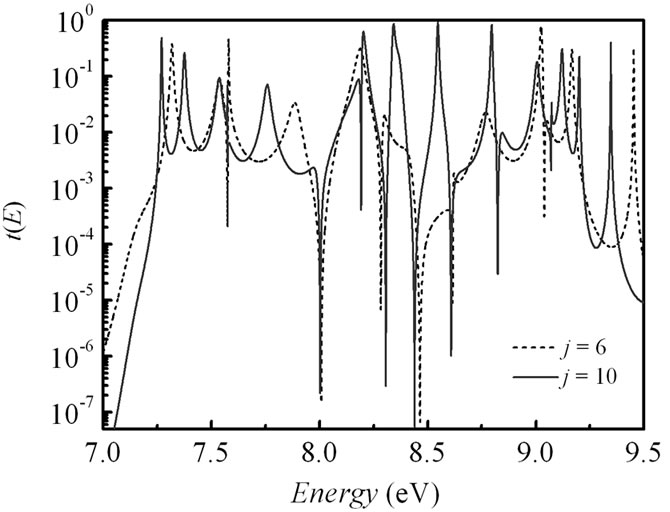
Figure 4. (a) Electronic transmission t(E) as a function of the energy E for j = 6 and j = 10 in 5'-G-(T)j-GGG-3' sequence of the DNA molecule. Inset: blow-up of the transmission in an energy window around the Fermi energy EF = 7.75 eV for j = 2 (solid line), 3 (shot dotted line), 6 (dashed line), 10 (solid red line), and 12 (dot-dashed line). Parameters: t|| = 0.5 eV, γL1= γL2= γR1= γR2= 1.5 eV. (b) Same as in (a) for the DNA molecule with (G:C)-(T:A)j-(G:C)3 configuration at t^ = 0.07 eV.
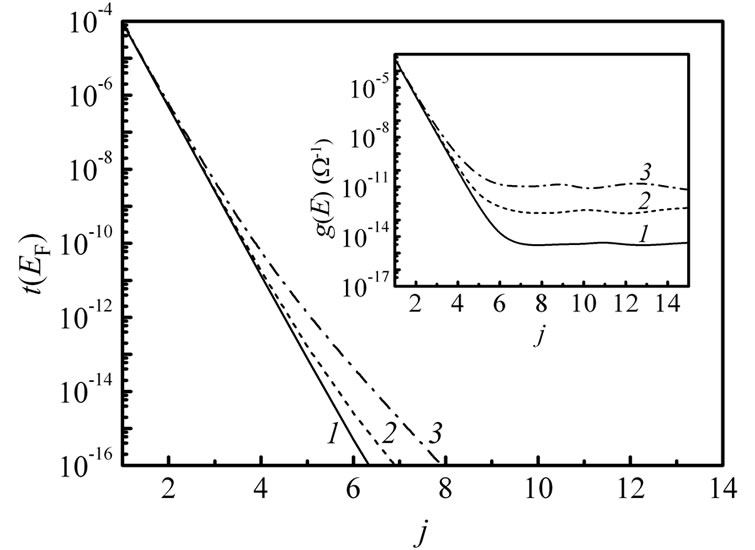
Figure 5. Transmission coefficient t(EF) vs. number of j (T:A) base pairs for different interstrand hopping integrals t^ (1 – 0.01 eV, 2 – 0.03 eV, 3 – 0.07 eV) at fixed t|| = 0.1 eV in the double-stranded (G:C)-(T:A)j-(G:C)3 molecule, when γL1= γL2= γR1= γR2= 1.5 eV. Inset: the corresponding conductance at T = 300 K at the same parameters.
ies [41-43]. Nevertheless, the crossover point is shifted to 6 (T:A) base pairs in that regime that is closer to the experimental results [28,41].
We find that reducing the parameter t|| for the two-leg DNA leads to a shift and change in width of the transmission resonances (Figure 6). Energetically, the transmission window moves towards larger energies with exponentially decreasing in the peak intensities. There is no much distinction in the transmission spectrum nearby the Fermi energy, where just two principal peaks are seen for j = 1 as well as for j = 10 (inset in Figure 6). Consequently, when the number of the (T:A) di-nucleotides is not more than 6, the single-step coherent tunneling through G-nucleotides in the duplex is the dominating
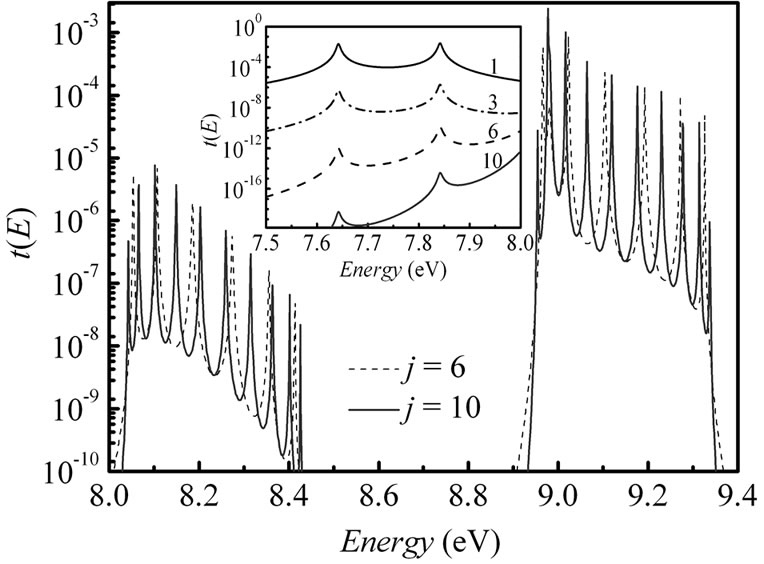
Figure 6. Energy dependent electronic transmission for j = 6 and j = 10 in the (G:C)-(T:A)j-(G:C)3 ladder structure of DNA. Inset: blow-up of the transmission in an energy window around the Fermi energy EF = 7.75 eV for j = 2 (solid line), 3 (shot dotted line), 6 (dashed line), and 10 (solid red line). Parameters: t|| = 0.1 eV, t^= 0.07 eV, γL1= γL2= γR1= γR2 = 1.5 eV.
transport mechanism. However, we suppose that the derived transition in the distance dependence of CTE can be distinctly modified by taking into account dynamic effects in the transmission [44] and environment. These factors and also the equalization of the Fermi energy of the DNA and the electrodes have an impact on the charge propagation pathways through DNA bridges changing significantly the energy levels of the bridging states [16].
We now consider the charge transport in the (G:C)- (T:A)j-(G:C)3 ladder that is coupled asymmetrically to the electrodes, when only the 5'-ends of the duplex are contacted. Figure 7 shows the dependence of the transmission function and conductance on the number of j (T:A)
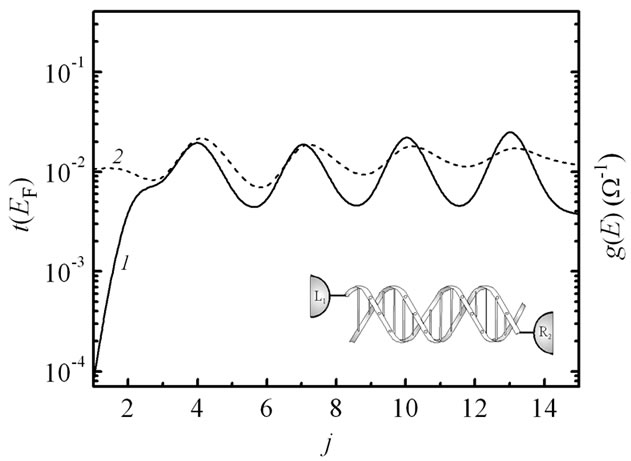
Figure 7. Transmission function t(EF) (curve 1) and conductance g(E) (curve 2) vs. number of j (T:A) base pair at T = 300 K and t|| = 0.5 eV, t^= 0.07 eV for the case of asymmetric coupling of the (G:C)-(T:A)j-(G:C)3 ladder to the electrodes: γL1 = γR2 = 1.5 eV, γL2 = γR1 = 0.
pairs for this contact geometry (γL1 = γR2 = 1.5 eV, γL2 = γR1 = 0). Now there is a single pathway for an electronic tunneling from the electrodes into the ladder. That results in increasing the delocalization of the electronic states in the DNA chains leading to a drastic effect on the transport characteristics. This effect is evidently displayed in Figure 8(a), where new transmission peaks appear irrespective of the number of the bridging sites and the spectrum becomes more fragmented. Thus the transmission and conductance of the duplex are characterized by large oscillations and do not practically depend on the bridge length. In Figure 8(b) the spectrum of the electronic transmission around the Fermi energy gives an apt illustration that the holes propagate through the set of the nucleobases choosing optimal pathways to move in the structure not following the well-defined transport mechanism. Meanwhile, in a case of the asymmetric contact coupling to the 3'-end molecular sites the conductance properties do not undergo any changes, while in [45], where electron-vibron coupling has been taken into account, the absolute values of the current considerably depends on the way the two strands are contacted to the electrodes. So, the model of the charge transport in the DNA duplex, proposed in [27] and extended here, is very sensitive to the DNA-metal contact topology.
The transport mechanism based on a fully coherent picture of charge propagation through the bridge seems to be questionable in the case of DNA. For instance, the inclusion of dynamical effects into our model may qualitatively modify the calculated length dependence. Furthermore, temperature effects in this model are only associated with the electrode Fermi functions; more involved temperature dependencies – observed in the case where charge motion is coupled to dynamical degrees of freedom [20] – may therefore not be adequately described.
Thus different mechanisms of charge motion in DNA cannot be distinguished only on the basis of the mea-
 (a)
(a)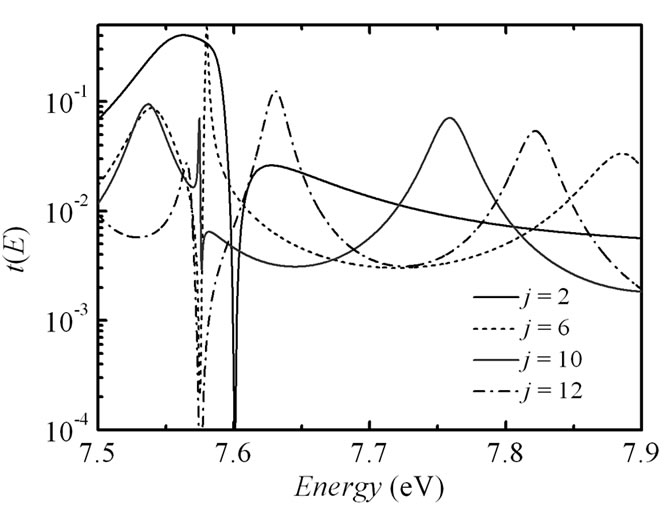 (b)
(b)
Figure 8. (a) Electronic transmission vs. energy E for j = 6 and j = 10 in (G:C)-(T:A)j-(G:C)3 sequence of the DNA molecule attached asymmetrically to the electrodes: γL1= γR2= 1.5 eV, γL2= γR1= 0, (t|| = 0.5 eV, t^= 0.07 eV); (b) Electronic transmission in an energy window around the Fermi energy EF = 7.75 eV at the same parameters.
sured length dependence of CTE. To decide what particular mechanism dominates under given experimental conditions, the whole range of experimental data including information on the hole mobility and diffusion coefficients should be considered.
4. CONCLUSIONS
We reproduced and extended the simulation results, obtained in [27], applying a tight-binding model to a system “electrode-DNA-electrode”, with standard transfer matrix formalism. It has been shown that the almost zero distance dependence of the charge transport cannot be a result of solely the interstrand coupling in DNA, but it is a consequence of a transition from under-barrier tunneling mechanism to over-barrier propagation when the nearest neighbor hopping is chosen large enough. It was demonstrated that the crossover from the strong to the weak distance-dependent charge transport in DNA can be controlled by the electronic coupling between the complementary nucleobases within a chain as well as by their coupling between two strands. Within the proposed model, we establish that the DNA-electrode coupling governs the conductance in the molecule. An asymmetry in the contact of the DNA duplex with the electrodes has a drastic effect on the conducting properties of the system, leading to a lessening in the localization length of the positive charges along the structure.
REFERENCES
- Arkin, M.R., Stemp, E.D.A., Holmlin, R.E., Barton, J.K., Hörmann, A., Olson, E.J.C. and Barbara, P.F. (1996) Rates of DNA-mediated electron transfer between metallointercalators. Science, 273(5274), 475-480.
- Dandliker, P.J., Núñez, M.E. and Barton, J.K. (1998) Oxidative charge transfer to repair thymine dimers and damage guanine bases in DNA assemblies containing tethered metallointercalators. Biochemistry, 37(18), 6491- 6502.
- Rajski, S.R., Jackson, B.A. and Barton, J.K. (2000) DNA repair: models for damage and mismatch recognition. Mutation Research, 447(1), 49-72.
- Grib, N.V., Berashevich, J.A. and Borisenko, V.E. (2006) Equivalent electrical network of the DNA molecule. Russian Microelectronics, 35(6), 398-404.
- Bhalla, V., Bajpai, R.P. and Bharadwaj, L.M. (2003) DNA electronics. European Molecular Biology Organization Reports, 4(5), 442-445.
- Niemeyer, C.M. and Adler, M. (2002) Nanomechanical devices based on DNA. Angewandte Chemie International Edition, 41(20), 3779-3783.
- Parker, J. (2003) Computing with DNA. European Molecular Biology Organization Reports, 4(1), 7-10.
- Rauhe, H., Vopper, G., Feldkamp, U., Banzhaf, W. and Howard, J.C. (2000) Digital DNA molecules. Proceedings of 6th DIMACS Workshop on DNA Based Computers, Leiden, 13-17 June 2000, 1-10.
- Jortner, J., Bixon, M., Langenbacher, T. and MichelBeyerle, M.E. (1998) Charge transfer and transport in DNA. Proceedings of the National Academy of Sciences of the United States of America, 95(22), 12759-12765.
- Berlin, Y.A., Burin, A.L. and Ranter, M.A. (2000) On the long-range charge transfer in DNA. Journal of Physical Chemistry A, 104(3), 443-445.
- Lewis, F.D., Wu, T., Zhang, Y., Letsinger, R.L., Greenfield, S.R. and Wasielewski, M.R. (1997) Distancedependent electron transfer in DNA hairpins. Science, 277(5326), 673-676.
- Porath, D., Cuniberti, G. and Di Felice, R. (2004) Charge transport in DNA-based devices. Topics in Current Chemistry, 237, 183-227.
- Gutierrez, R., Porath, D., Cuniberti, G. and Baranowski, S. (2006) Charge transport in disordered solids with applications in electronics. John Wiley & Sons Inc., Hoboken.
- Porath, D., Bezryadin, A., De Vries, S. and Dekker, C. (2000) Direct measurement of electrical transport through DNA molecules. Nature, 403(6670), 635-638.
- Xu, B., Zhang, P., Li, X. and Tao, N. (2004) Direct conductance measurement of single DNA molecules in aqueous solution. Nano Letters, 4(6), 1105-1108.
- Cohen, H., Nogues, C., Naaman, R. and Porath, D. (2005) Direct measurement of electrical transport through single DNA molecules of complex sequence. Proceedings of the National Academy of Sciences of the United States of America, 102(33), 11589-11593.
- Cohen, H., Nogues, C., Ullien, D., Daube, S., Naaman, R. and Porath, D. (2006) Electrical characterization of selfassembled singleand double-stranded DNA monolayers using conductive AFM. Faraday Discussions, 131, 367- 376.
- Shapir, E., Cohen, H., Calzolari, A., Cavazzoni, C., Ryndyk, D.A., Cuniberti, G., Kotlyar, A.B., Di Felice, R. and Porath, D. (2008) Electronic structure of single DNA molecules resolved by transverse scanning tunnelling spectroscopy. Nature Materials, 7(1), 68-74.
- Yu, Z.G. and Song, X. (2001) Variable range hopping and electrical conductivity along the DNA double helix. Physical Review Letters, 86(26), 6018-6021.
- Renger, T. and Marcus, R.A. (2003) Variable-range hopping electron transfer through disordered bridge states: Application to DNA. Journal of Physical Chemistry A, 107(41), 8404-8419.
- Cuniberti, G., Craco, L., Porath, D. and Dekker, C. (2002) Backbone-induced semiconducting behavior in short DNA wires. Physical Review B, 65(24), 241314.
- Roche, S. (2003) Sequence dependent DNA-mediated conduction. Physical Review Letters, 91(10), 108101.
- Wang, H., Lewis, J.P. and Sankey, O.F. (2004) Band-gap tunneling states in DNA. Physical Review Letters, 93(1), 016401.
- Zhang, W. and Ulloa, S.E. (2004) Extended states in disordered systems: Role of off-diagonal correlations. Physical Review B, 69(15), 153203.
- Cuenda, S. and Sanchez, A. (2004) Disorder and fluctuations in nonlinear excitations in DNA. Fluctuation and Noise Letters, 4(3), L491-L504.
- Fialko, N.S. and Lakhno, V.D. (2000) Nonlinear dynamics of excitations in DNA. Physical Letters A, 278(2), 108-111.
- Wang, X.F. and Chakraborty, T. (2006) Charge transfer via a two-strand superexchange bridge in DNA. Physical Review Letters, 97(10), 106602.
- Giese, B., Amaudrut, J., Köhler, A.-K., Spormann, M. and Wessely, S. (2001) Direct observation of hole transfer through DNA by hopping between adenine bases and by tunnelling. Nature, 412(6844), 318-320.
- Lewis, F.D., Zhu, H., Daublain, P., Cohen, B. and Wasielewski, M.R. (2006) Hole mobility in DNA A tracts. Angewandte Chemie International Edition, 45(47), 7982- 7985.
- Treadway, C.R., Hill, M.G. and Barton, J.K. (2002) Charge transport through a molecular π-stack: Double helical DNA. Chemical Physics, 281(2-3), 409-428.
- Seidel, A.M., Schulz, A. and Sauer, H.M. (1996) Nucleobase-specific quenching of fluorescent dyes. 1. Nucleobase one-electron redox potentials and their correlation with static and dynamic quenching efficiencies. Journal of Physical Chemistry, 100(13), 5541-5553.
- Gutiérrez, R., Mandal, S. and Cuniberti, G. (2005) Dissipative effects in the electronic transport through DNA molecular wires. Physical Review B, 71(23), 235116.
- Sugiyama, H. and Saito, I. (1996) Theoretical studies of GG-specific photocleavage of DNA via electron transfer:
 significant lowering of ionization potential and 5’-localization of HOMO of stacked GG bases in B-form DNA. Journal of the American Chemical Society, 118(30), 7063- 7068.
significant lowering of ionization potential and 5’-localization of HOMO of stacked GG bases in B-form DNA. Journal of the American Chemical Society, 118(30), 7063- 7068. - Endres, R.G., Cox, D.L. and Singh, R.R.P. (2004) Colloquium: The quest for high-conductance DNA. Reviews of Modern Physics, 76(1), 195-214.
- Senthilkumar, K., Grozema, F.C., Guerra, C.F, Bickelhaupt, F.M., Lewis, F.D., Berlin, Y.A., Ratner, M.A. and Siebbeles, L.D.A. (2005) Absolute rates of hole transfer in DNA. Journal of the American Chemical Society, 127(42), 14894-14903.
- Nitzan, A., Jortner, J., Wilkie, J., Burin, A.L. and Ratner, M.A. (2000) Tunneling time for electron transfer reactions. Journal of Physical Chemistry B, 104(24), 5661-5665.
- Macia, E., Triozon, F. and Roche, S. (2005) Contactdependent effects and tunneling currents in DNA molecules. Physical Review B, 71(11), 113106.
- Williams, T.T., Odon, D.T. and Barton, J.K. (2000) Variations in DNA charge transport with nucleotide composition and sequence. Journal of the American Chemical Society, 122(37), 9048-9049.
- Sartor, V., Boone, E. and Schuster, G.B. (2001) Longdistance radical cation migration through A/T base pairs in DNA: An experimental test of theory. Journal of Physical Chemistry B, 105(45), 11057-11059.
- De Pablo, P.J., Moreno-Herrero, F., Colchero, J., GomezHerrero, J., Herrero, P., Baro, A.M., Ordejon, P., Soler, J.M. and Artacho, E. (2000) Absence of dc-conductivity in λ-DNA. Physical Review Letters, 85(23), 4992-4995.
- Segal, D., Nitzan, A., Davis, W.B., Wasielewski, M.R. and Ratner, M.A. (2000) Electron transfer rates in bridged molecular systems 2. A steady-state analysis of coherent tunneling and thermal transitions. Journal of Physical Chemistry B, 104(16), 3817-3829.
- Valis, L., Wang, Q., Raytchev, M., Buchvarov, I., Wagenknecth, H.A. and Fiebig, T. (2006) Base pair motions control the rates and distance dependencies of reductive and oxidative DNA charge transfer. Proceedings of the National Academy of Sciences of the United States of America, 103(27), 10192-10195.
- Voityuk, A.A. (2005) Charge transfer in DNA: Hole charge is confined to a single base pair due to solvation effects. Journal of Chemical Physics, 122(20), 204904.
- Davies, O.R. and Inglesfield, J.E. (2004) Embedding method for conductance of DNA. Physical Review B, 69, 195110.
- Gutierrez, R., Mohapatra, S., Cohen, H., Porath, D. and Cuniberti, G. (2006) Inelastic quantum transport in a ladder model: Implications for DNA conduction and comparison to experiments on suspended DNA oligomers. Physical Review B, 74(23), 235105.
NOTES
*This work was funded by the Deutsche Forschungsgemeinschaft (SPP 1243) and Collaborative Research Center SFB 689.