M. J. Duechs et al. / Open Journal of Immunology 2 (2012) 17-24
Copyright © 2012 SciRes.
23
airway hyperresponsiveness and allergic airway inflam-
mation [22]. Why did we not see these effects? We can
currently not explain the divergent results, however these
might be due to the difference in concentrations applied,
as Sel et al. use a 10 times higher dose of poly(I:C) and a
2.5 times higher dose of R848. Our finding that CPG-
ODN are very potent inhibitors of the development of
allergen-specific Th2 responses is in line with previously
published reports [16-19].
OPEN A CCESS
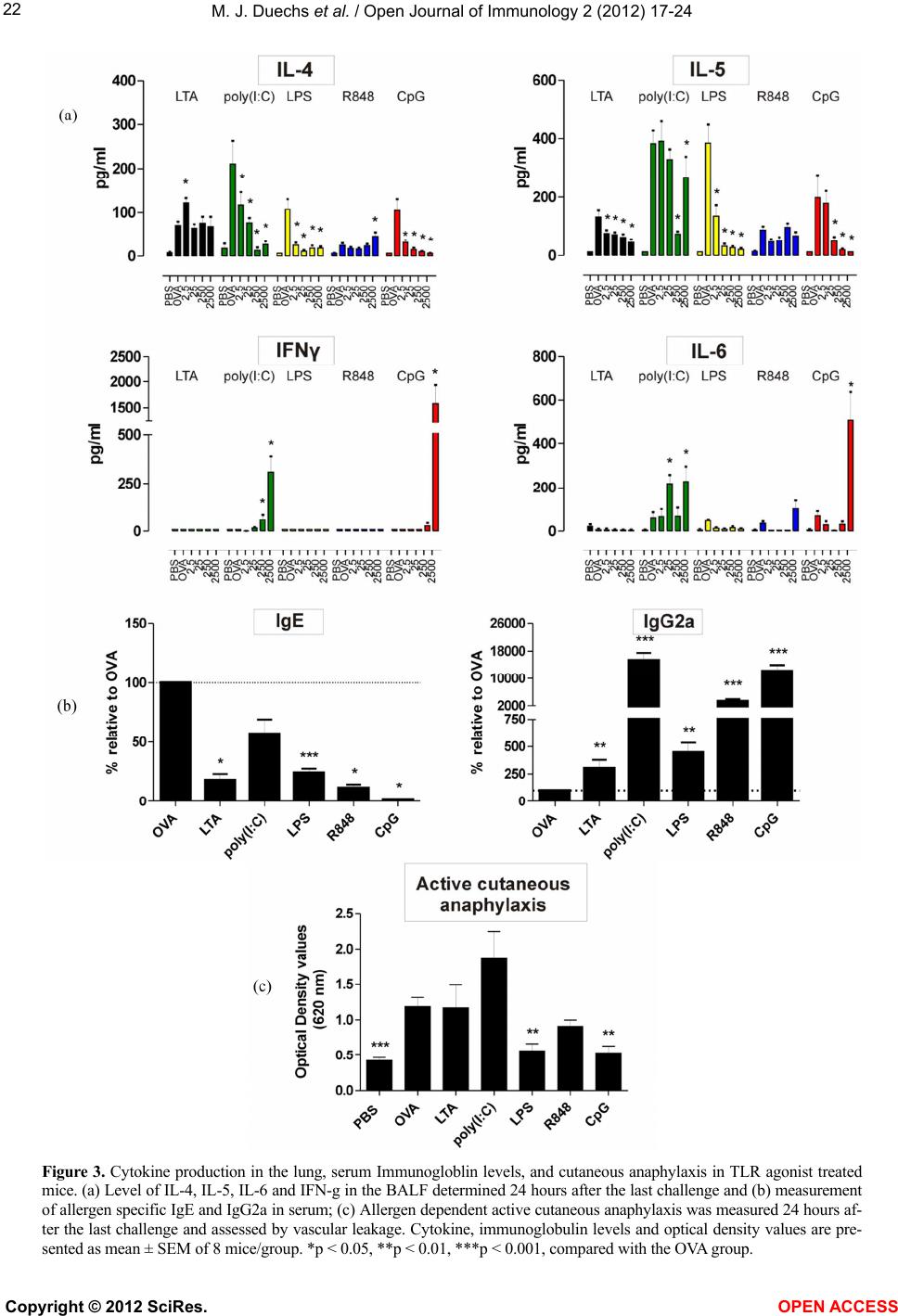
A very interesting finding of our study was that the
induction of strong allergen-specific Th1 responses did
not always correlate with a strong reduction in Th2 re-
sponses. Poly(I:C) had the second strongest Th1-induc-
ing effect, but did not reduce OVA-specific IgE levels in
the serum, goblet cell metaplasia in the lung or cutaneous
anaphylaxis. Another surprising effect was that only LPS
and CpG were able to significantly reduce all Th2 pa-
rameters measured. This clearly suggests that only parts
of the allergen-specific Th2 response could be modulated
by R848, poly(I:C) and LTA and that the effect varied
from agonist to agonist. For example, LTA reduced air-
way eosinophilia, IL-5, and IgE levels but not IL-4, gob-
let cell metaplasia or cutaneous anaphylaxis. In contrast
R848 did not reduce airway eosinophilia, IL-4 or IL-5
levels but reduced IgE levels. We have no explanation
why some parameters are affected and some not. How-
ever, it is clear that it depends on the TLR agonist used.
Taken together our results clearly show that LPS and
CpG have the strongest suppressive effects on the de-
velopment of numerous allergen-specific Th2 responses
in mice. TLR9 and TLR4 agonists have already been
used as adjuvants in clinical SIT trials [9,23] and our
results support the further clinical testing of these ago-
nists. Interestingly, the strong induction of allergen-spe-
cific Th1 responses did not always correlate with a strong
reduction in the allergic response, suggesting that this
measure of an effective SIT response may not translate in
a reduction in the allergic response.
REFERENCES
[1] Barnes, P.J. (2010) New therapies for asthma: Is there any
progress? Trends in Pharmacological Sciences, 31, 335-
343. doi:10.1016/j.tips.2010.04.009
[2] Walsh, G.M. (2006) Targeting airway inflammation: Novel
therapies for the treatment of asthma. Current Medicinal
Chemistry, 13, 3105-3111.
doi:10.2174/092986706778521779
[3] Holgate, S.T. and Polosa, R. (2008) Treatment strategies
for allergy and asthma. Nature Reviews Immunology, 8,
218-230. doi:10.1038/nri2262
[4] Durham, S.R., Walker, S.M., Varga, E.M., Jacobson, M.R.,
O’Brien, F., Noble, W., Till, S.J., Hamid, Q.A. and Nouri-
Aria, K.T. (1999) Long-term clinical efficacy of grass-
pollen immunotherapy. New England Journal of Medi-
cine, 341, 468-475. doi:10.1056/NEJM199908123410702
[5] Eifan, A.O., Akkoc, T., Yildiz, A., Keles. S., Ozdemir, C.,
Bahceciler, N.N. and Barlan, I.B. (2010) Clinical efficacy
and immunological mechanisms of sublingual and sub-
cutaneous immunotherapy in asthmatic/rhinitis children
sensitized to house dust mite: An open randomized con-
trolled trial. Clinical & Experimental Allergy, 40, 922-
932. doi:10.1111/j.1365-2222.2009.03448.x
[6] Moingeon, P., Batard, T., Fadel, R., Frati, F., Sieber, J.
and Overtvelt, L. (2006) Immune mechanisms of allergen
specific sublingual immunotherapy. Allergy, 61, 151-165.
doi:10.1111/j.1398-9995.2006.01002.x
[7] Tversky, J.R., Bieneman, A.P., Chichester, K.L., Hamilton,
R.G. and Schroeder, J.T. (2010) Subcutaneous allergen
immunotherapy restores human dendritic cell innate im-
mune function. Clinical & Experimental Allergy, 40, 94-
102.
[8] Calamita, Z., Saconato, H., Pela, A.B. and Atallah, N.
(2006) Efficacy of sublingual immunotherapy in asthma:
systematic review of randomized clinical trials using the
Cochrane Collaboration method. Allergy, 61, 1162-1172.
doi:10.1111/j.1398-9995.2006.01205.x
[9] Senti, G., Johansen, P., Haug, S., Bull, C., Gottschaller, C.,
Müller, P., Pfister, T., Maurer, P., Bachmann, M.F. and
Graf, N. (2009) Use of A-type CpG oligodeoxynucleo-
tides as an adjuvant in allergen specific immunotherapy
in humans: A phase I/IIa clinical trial. Clinical & Expe-
rimental Allergy, 39, 562-570.
doi:10.1111/j.1365-2222.2008.03191.x
[10] Kündig, T.M., Senti, G., Schnetzler, G., Wolf, C., Prinz,
Vavricka, B.M., Fulurija, A., Hennecke, F., Sladko, K.,
Jennings, G.T. and Bachmann, M.F. (2006) Der p 1 pep-
tide on virus-like particles is safe and highly immuno-
genic in healthy adults. Journal of Allergy and Clinical
Immunology, 117, 1470-1476.
doi:10.1016/j.jaci.2006.01.040
[11] Trujillo-Vargas, C.M., Werner-Klein, M., Wohlleben, G.,
Polte T., Hansen, G., Ehlers, S., Erb, K.J. (2007) Helminth-
derived products inhibit the development of allergic re-
sponses in mice. American Journal of Respiratory and
Critical Care Medicine, 175, 336-344.
doi:10.1164/rccm.200601-054OC
[12] Racila, D.M. and Kline, J.N. (2005) Perspectives in asthma:
Molecular use of microbial products in asthma prevention
and treatment. The Journal of Allergy and Clinical Im-
munology, 116, 1202-1205.
doi:10.1016/j.jaci.2005.08.050
[13] Rolland, J.M., Gardne, L.M. and O’Hehir, R.E. (2009)
Allergen-related approaches to immunotherapy. Pharma-
cology & Therapeutics, 121, 273-284.
doi:10.1016/j.pharmthera.2008.11.007
[14] Wohlleben, G. and Erb, K.J. (2006) Immune stimulatory
strategies for the prevention and treatment of asthma.
Current Pharmaceutical Design, 12, 3281-3292.
doi:10.2174/138161206778194114
[15] Trujillo-Vargas, C.M., Mayer, K.D., Bickert, T., Palmet-
shofer, A., Grunewald, S., Ramirez-Pineda, J.R., Polte, T.,
Hansen, G., Wohlleben, G. and Erb, K.J. (2005) Vaccina-
tions with T-helper type 1 directing adjuvants have dif-