American Journal of Plant Sciences
Vol.05 No.20(2014), Article ID:50181,5 pages
10.4236/ajps.2014.520322
Effects of Cu2+ on Wheat Seedlings Exposed to Enhanced Ultraviolet-B Radiation
Liyan Yang1, Zhaoqing Wang1, Yuqi Hou1, Rong Han1, Yi Sun2,3*
1Shanxi Normal University, Linfen, China
2Biotechnology Research Centre, Shanxi Academy of Agricultural Sciences, Taiyuan, China
3Key Laboratory of Crop Gene Resources and Germplasm Enhancement on Loess Plateau, Ministry of Agriculture, Taiyuan, China
Email: *sunyi692003@163.com
Copyright © 2014 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 20 July 2014; revised 21 August 2014; accepted 12 September 2014
ABSTRACT
To explore the wheat seedling development and physiological responses under copper contamination and enhanced ultraviolet-B (UV-B) irradiation, 10 mg·L−1 CuCl2 solution was irrigated to Triticum aestivum L. cv. Linyuan 2069 one day after germination with or without ultraviolet-B (10.08 kJ m−2·d−1) light exposure, respectively. The results showed that Cu2+ and UV-B caused various adverse effects on wheat seedling development. Cu2+ hindered root development by significantly reducing root number, while UV-B dwarfed seedling height and decreased the leaf length. Chlorophyll content and activity of ATPase in thylakoid membrane of wheat leaves dropped significantly under enhanced UV-B while the activity of ATPase in plasma membrane of seedling root was significantly decreased in Cu2+ group. Relative electric conductivity of leaves significantly increased in both Cu2+ and UV-B groups, so did the biomass. We also observed that combined Cu2+ and UV-B showed more adverse effects on wheat seedlings than either of them alone except for root growth.
Keywords:
Cu2+, seedling development, UV-B radiation, wheat

1. Introduction
Large areas of soil have been contaminated by heavy metals that are deleterious to the existence, reproduction and development of living organisms including plants, animals and microorganisms. It has even threatened the health of ecosystem and also human beings [1] . Soil may contain elevated levels of copper due to prolonged use of organic fertilizers composed of sewage sludge and poultry manure and some copper containing fungicides, bactericides and pesticide to control diseases [2] . Copper (Cu) is an essential element at low concentration that is involved in a number of physiological process, and also a part of the prosthetic group of many enzymes including ATP synthesis. At the level of whole plants, however, excessive copper reduces iron accumulation, decreases chlorophyll and carotenoids content of Elsholtzia splendens [3] .
Ultraviolet-B (UV-B) light (280 - 320 nm) is a natural component of solar radiation. However, with depletion of the stratospheric ozone layer, enhanced levels of UV-B radiation are now reaching the earth’s surface [4] . Plants show various responses to these higher levels of UV-B irradiation such as photosynthesis, plant height and total mass production reduction [5] [6] .
Wheat is one of the major cereal crops widely cultivated throughout the world, and its yield is very important for world food security. The effects of enhanced UV-B on wheat growth, physiology, biomass as well as trace element accumulation have been investigated [7] - [9] . Previous studies showed that excessive Cu inhibited root length of wheat and caused a decrease in the lipid to protein ratio as well as a change in the phospholipid composition [10] ; it also reduced plant growth, enhanced electrolyte leakage from cells as well as increased lipid peroxidation and protein carbonylation [11] . Proteome analysis showed that copper stress decreased wheat seedling protein expression which is involved in carbohydrate metabolism, protein metabolism, and photosynthesis [12] . Combined UV-B and copper on growth, oxidative stress and ascorbate-glutathione cycle in two cyanobacteria have been studied [13] . However, studies on the effects of combined UV-B and Cu on wheat development were limited. At the present study, we explored effects of combined Cu and enhanced UV-B radiation on the development of winter wheat seedlings; we also investigated their chlorophyll content, relative electric conductivity and ATP synthase (ATPase) activity.
2. Materials and methods
2.1. Plant material and treatments
Winter wheat (Triticum aestivum L. cv. Linyuan 2069) seeds were provided by the Wheat Research Institute, Shanxi Academy of Agricultural Sciences (SAAS), People’s Republic of China. They were selected for uniform size and sterilized for 10 min with 0.1% HgCl2 and washed for 50 min with running water. Thirty seeds were cultured on wet filter paper in each petri dish (diameter
The UV-B radiation intensity was 10.08 kJ m−2·d−1. The spectral irradiance from the lamps was determined with an Optronics (Model 742 Optronics Laboratories, Orlando, FL, USA) spectroradiometer. The UV-B radiation was generated by a filtered lamp (30 W, 297 nm, Qin Brand, Baoji Lamp Factory, Baoji City, China). The lamps were hung on the top of the petri dishes and the desired irradiation was obtained by adjusting the distance between the lamps and the petri dishes.
One day later, just as seeds were germinating, Cu2+ was applied in the form of CuCl2 with 10 mg/l solution for C and CB group, and the same amount of water was applied for B and CK group (Table 1).
2.2. Leaf length, root number and biomass measurement
On the eighth day of the investigation, twenty seedlings per replication were randomly chosen from each treatment, and seedling leaf length, root number, fresh weight, and dry weight were recorded.

Table 1. Light/dark period of irradiation treatments.
CK: distilled water only; C: CuCl2 application only; B: distilled water with ultraviolet-B radiation; CB: CuCl2 with ultraviolet-B radiation.
2.3. Relative electric conductivity and Chlorophyll content measurement
On the eighth day of the investigation, relative electric conductivity and chlorophyll content of leaves were measured according to Lee [14] . Leaf discs from each treatment were placed into a glass tube containing distilled water, vacuumed then shaken to measure the initial electric conductivity (S1). Subsequently, the glass tube with leaf discs was heated in boiling water and cooled to room temperature to determine the final electric conductivity (S2). The distilled water was used as the blank (S0). The relative electrical conductivity (REC) was evaluated as REC(%) = (S1 − S0)/(S2 − S0) × 100.
Chlorophyll a and b (chl a and chl b) contents were measured using spectrophotometric method using the following equations after correction with the value measured: chl a = 12.7A663 − 2.69A645; chl b = 22.9A645 − 4.68A663; chl a + chl b = 8.02A663 + 20.21A645.
2.4. ATPase activity measurement
On the eighth day of the investigation, activity of ATPase in thylakoid membrane of leaves as well as in plasma membrane of root cell was measured by amount of liberated inorganic phosphate (Pi) according to Chen [15] . The ATPase activity was expressed in ug Pi per mg protein in 1 min. All experiments were performed in triplicate.
2.5. Statistical analysis
Statistical significance was estimated at p < 0.05 according to Duncan’s multiple range test. All data give mean ± SD.
3. Results
3.1. The effects of Cu2+ on leaf length, root number and biomass of seedlings under enhanced UV-B
Cu2+ and UV-B showed varied effects on wheat seedlings. Cu2+ solely inhibited the root development, while UV-B dwarfed the shoot and diminished the leaf area. In C group, root number was the least among the 4 groups (p < 0.05), so was the root length (Figure 1), while under combined UV-B and Cu2+, no significant difference was observed compared to the control group (p > 0.05). Leaf length was the highest in the control group, it was significantly lower in B group than C group (p < 0.05), and it was the shortest under combined UV-B and Cu2+. Both UV-B and Cu2+ decreased wheat seedling biomass (both fresh weight and dry weight) compared to the control (p < 0.05) and the lowest biomass appeared in CB group. No significant difference was observed among B, C, and CB groups, though the fresh weight and dry weight were 3.1% and 9.5% lower in B group than C group, respectively (Table 2).
3.2. The effect of Cu2+ on relative electric conductivity and chlorophyll content in seedling leaves under enhanced UV-B
As shown in Table 3, compared with the control group, relative electric conductivity of leaves significantly increased in both C and B groups (p < 0.05), and it was 4.1% higher in C group than B group. In CB group, relative electric conductivity was the highest. Chlorophyll content was significantly reduced in B group (p < 0.05) than the control group, and no significant difference was observed between CK and C group. Chlorophyll content was the lowest in CB group.
3.3. The effect of Cu2+ on ATPase activity in seedlings under enhanced UV-B
The effect of Cu2+ and UV-B on ATPase activity varied with their location. In C group, the activity of ATPase located in plasma membrane of seedling root was significantly lower than the control group (p < 0.05), and it was not significantly different between B and control group (p > 0.05). In CB group, the activity of ATPase was the lowest, which was 26.8% lower than C group, and significantly lower than the control and B group (p < 0.05). As for ATPase located in thylakoid membrane, their activities significantly decreased in B group (p < 0.05), and no significant difference appeared between C group and control (p > 0.05). The ATPase in CB group
Figure 1. Seedling development in various treatments. CK: dis- tilled water only; B: distilled water with ultraviolet-B radiation; C: Cu2+ application only; CB: Cu2+ with ultraviolet-B radiation.
Table 2. Leaf length, root number and biomass in various treatments.
CK: distilled water only; B: distilled water with ultraviolet-B radiation; C: Cu2+ application only; CB: Cu2+ with ultraviolet-B radiation. Values are means ± SD (n = 60), and values in the same column followed by various letters denote significant difference at p < 0.05.
Table 3. Effects of UV-B and Cu2+ on relative electric conductivity and chlorophyll content in seedling l eaves under enhanced UV-B.
CK: distilled water only; B: distilled water with ultraviolet-B radiation; C: Cu2+ application only; CB: Cu2+ with ultraviolet-B radiation. Values are means ± SD (n = 3), and values in the same column followed by various letters denote significant difference at p < 0.05.
were the lowest which was 48.8%, 45.2% and 29.3% lower than the control, C and B groups, respectively (Figure 2).
4. Discussion
During the long evolution process, plants have evolved inducible mechanisms to defend themselves from environmental concerns, including metabolic and morphological changes. The present study showed enhanced UV-B and Cu2+ appeared to have adverse effects on wheat seedlings. However, the mechanisms underlying varied. Dwarfed wheat seedling height and decreased leaf elongation might well serve to diminish the enhanced UV-B irradiation, which might influence photosynthesis and reduce photosynthetic accumulation; while Cu2+ enters plants mainly through the root system, so the significantly decreased root number and root length might be the main adaptive ways to excessive Cu2+. Under field conditions, under-ground parts, such as seeds and roots are protected from UV-B irradiation, consequently, damage from ultraviolet light is minimal. In our study, enhanced UV-B hindered the root development; we owed it to the root exposure to UV-B. However, the root
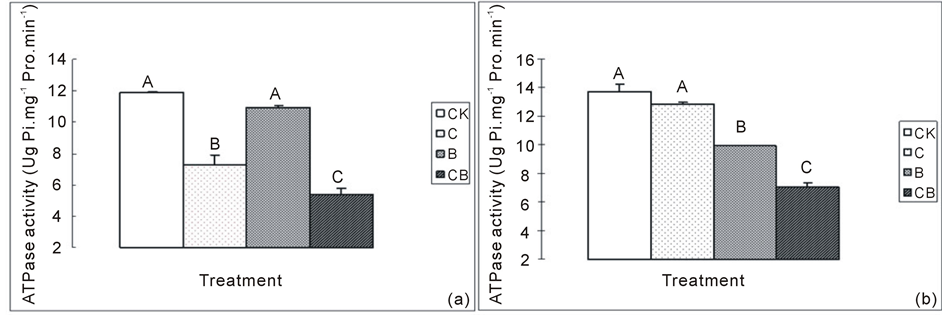
Figure 2. ATPase activity in various treatments. (a) Activity of ATPase in plasma membrane of seedling root; (b) Activity of ATPase in thylakoid membrane of wheat leaves. CK: distilled water only; B: distilled water with ultraviolet-B radiation; C: Cu2+ application only; CB: Cu2+ with ultraviolet-B radiation. Each bar is the mean ± SD (n = 3) for each treatment. Bars with various letters denote significant difference at p < 0.05.
development recovered under CB group, we speculated that UV-B and Cu2+ might have antagonistic effect on root growth.
Under enhanced UV-B, yellow spots or streak appeared in the treated leaves, the biomacromolecules such as certain proteins can absorb ultraviolet-B, but when the UV-B is too strong and beyond the plant tolerant capacity, the excessive UV-B light might injure the chloroplast and cause the reduction of chlorophyll content [16] , while it was not the case in Cu2+ group where no significant chlorophyll content difference appeared compared to the control group.
The target of UV-B radiation is the membrane [17] , which can be damaged by increased reactive oxygen species (ROS) in the plant cell [18] [19] . In our investigation, membrane permeability was enhanced in both B and C groups. We speculate that the membrane can be damaged in various pathways. Among them, UV-B light beyond the cellar proteins’ absorption capacity damaged the cell membrane in B group, or Cu2+ absorption by root cells and then partially transported to leaves in C group. Previous study demonstrated that excessive Cu2+ leads to root cell membrane oxidative system damage [20] , which can explain the significant reduction of ATPase activity in plasma membrane of seedling root. Enhanced ultraviolet-B resulted in adaptive changes on photosynthetic apparatus such as thylakoid membrane [21] . The ATPase of chloroplasts is an important enzyme that provides energy for the cell to use through the synthesis of ATP which is the most commonly used “energy currency” of cells in most organisms. ATPase activity of chloroplasts was significantly inhibited under UV-B radiation, which implied that the ATP production might be hindered. Furthermore, ATPase in root cell membrane was less sensitive to UV-B, but their activity was declined too. Our results also proved that combined Cu2+ and UV-B caused more adverse effects on wheat seedlings than either of them alone except for root development.
Acknowledgements
This research project was supported by the Fund Program for the Scientific Activities of Selected Returned Overseas Professionals in Shanxi Province (2014-95) and the National Nature Science Foundation of Shanxi Normal University (SMYKZ-30).
References
- Lombardi, L. and Sebastiani, L. (2005) Copper Toxicity in Prunuscerasifera: Growth and Antioxidant Enzymes Responses of in Vitro Grown Plants. Plant Science, 168, 797-802. http://dx.doi.org/10.1016/j.plantsci.2004.10.012
- Alaoui-Sossé, B., Genet, P., Vinit-Dunand, F., Toussaint, M., Epro, D. and Badot, P. (2004) Effect of Copper on Growth in Cucumber Plants (Cucumis sativus) and Its Relationships with Carbohydrate Accumulation and Changes in Ion Contents. Plant Science, 166, 1213-1218. http://dx.doi.org/10.1016/j.plantsci.2003.12.032
- Peng, H., Kroneck, P.M. and Küpper, H. (2013) Toxicity and Deficiency of Copper in Elsholtzia splendens Affect Photosynthesis Biophysics, Pigments and Metal Accumulation. Environmental Science & Technology, 47, 6120-6128.
- Jansen, M.A.K. (2002) Ultraviolet-B Radiation Effects on Plants: Induction of Morphogenic Responses. Physiologia Plantarum, 116, 423-429. http://dx.doi.org/10.1034/j.1399-3054.2002.1160319.x
- Fabón, G., Monforte, L., Tomás-Las-Heras, R., Núñez-Olivera, E. and Martínez-Abaigar, J. (2012) Dynamic Response of UV-Absorbing Compounds, Quantum Yield and the Xanthophyll Cycle to Diel Changes in UV-B and Photosynthetic Radiations in an Aquatic Liverwort. Journal of Plant Physiology, 169, 20-26. http://dx.doi.org/10.1016/j.jplph.2011.08.010
- Majer, P. and Hideg, É. (2011) Developmental Stage Is an Important Factor That Determines the Antioxidant Responses of Young and Old Grapevine Leaves under UV Irradiation in a Greenhouse. Plant Physiology and Biochemistry, 50, 15-23. http://dx.doi.org/10.1016/j.plaphy.2011.09.018
- Yang, S.H., Wang, L.J. and Li, S.H. (2007) Ultraviolet-B Irradiation-Induced Freezing Tolerance in Relation to Antioxidant System in Winter Wheat (Triticum aestivum L.) Leaves. Environmental and Experimental Botany, 60, 300- 307. http://dx.doi.org/10.1016/j.envexpbot.2006.12.003
- Lizana, X., Hess, S. and Calderini, D.F. (2009) Crop Phenology Modifies Wheat Responses to Increased UV-B Radiation. Agricultural and Forest Meteorology, 149, 1964-1974. http://dx.doi.org/10.1016/j.agrformet.2009.07.003
- Agrawal, S.B. and Rathore, D. (2007) Changes in Oxidative Stress Defense System in Wheat (Triticum aestivum L.) and Mung Bean (Vigna radiata L.) Cultivars Grown with and without Mineral Nutrients and Irradiated by Supplemental Ultraviolet-B. Environmental and Experimental Botany, 59, 21-33. http://dx.doi.org/10.1016/j.envexpbot.2005.09.009
- Quartacci, M.F., Cosi, E. and Navari-Izzo, F. (2001) Lipids and NADPH-Dependent Superoxide Production in Plasma Membrane Vesicles from Roots of Wheat Grown under Copper Deficiency or Excess. Journal of Experimental Botany, 52, 77-84. http://dx.doi.org/10.1093/jexbot/52.354.77
- Gajewska, E. and SkŁodowska, M. (2010) Differential Effect of Equal Copper, Cadmium and Nickel Concentration on Biochemical Reactions in Wheat Seedlings. Ecotoxicology and Environmental Safety, 73, 996-1003. http://dx.doi.org/10.1016/j.ecoenv.2010.02.013
- Li, G., Peng, X., Xuan, H., Wei, L., Yang, Y., Guo, T. and Kang, G. (2013) Proteomic Analysis of Leaves and Roots of Common Wheat (Triticum aestivum L.) under Copper-Stress Conditions. Journal of Proteome Research, 12, 4846- 4861. http://dx.doi.org/10.1021/pr4008283
- Singh, V.P., Srivastava, P.K. and Prasad, S.M. (2012) Differential Effect of UV-B Radiation on Growth, Oxidative Stress and Ascorbate-Glutathione Cycle in Two Cyanobacteria under Copper Toxicity. Plant Physiology and Biochemistry, 61, 61-70. http://dx.doi.org/10.1016/j.plaphy.2012.09.005
- Lee, H.S. (2000) Principles and Experimental Techniques of Plant Physiology and Biochemistry. Higher Education Press, Beijing.
- Chen, H.D., Xu, H. and Lian, Y.W. (2005) Experimental Techniques of Modern Plant Physiology. Science Press, Beijing.
- Warrant, J.M., Bassman, J.H. and Eigenbrode, S. (2002) Leaf Chemical Changes Induced in Populus trichocarpa by Enhanced UV-B Radiation and Concomitant Effects on Herbivory by Chrysomela scripta (Coleoptera: Chrysomelidae). Tree Physiology, 22, 1137-1146. http://dx.doi.org/10.1093/treephys/22.15-16.1137
- Zhou, X.M., Xi, Z.M., Jiao, X.L., Zhang, Z.W. and Wu, L.Y. (2007) Relationship between Photosynthesis and Leaf Ages of Grapevine under Enhanced Ultraviolet-B Radiation. Agricultural Research in the Arid Areas, 25, 216-220. (In Chinese)
- Ledford, H.K. and Niyogi, K.K. (2005) Singlet Oxygen and Photo-Oxidative Stress Management in Plants and Algae. Plant, Cell and Environment, 28, 1037-1045. http://dx.doi.org/ 10.1111/j.1365-3040.2005.01374.x
- Asada, K. (2006) Production and Scavenging of Reactive Oxygen Species in Chloroplasts and Their Functions. Plant Physiology, 141, 391-396. http://dx.doi.org/10.1104/pp.106.082040
- Ali, M.B., Singh, N., Shohael, A.M., Hahn, E.J. and Paek, K.Y. (2006) Phenolics Metabolism and Lignin Synthesis in Root Suspension Cultures of Panax ginseng in Response to Copper Stress. Plant Science, 171, 147-154. http://dx.doi.org/10.1016/j.plantsci.2006.03.005
- Yang, L.Y., Han, R. and Sun, Y. (2012) Damage Repair Effect of He-Ne Laser on Wheat Exposed to Enhanced Ultraviolet-B Radiation. Plant Physiology and Biochemistry, 57, 218-221.
NOTES

*Corresponding author.