Applied Mathematics
Vol.5 No.12(2014), Article
ID:47368,8
pages
DOI:10.4236/am.2014.512165
On the Fractal Design in Human Brain and Nervous Tissue
Gabriele A. Losa
Institute of Interdisciplinary Studies [ISSI] the Virtual Fractal Lab, Locarno, Switzerland
Email: glosa@cerfim.ch
Copyright © 2014 by author and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 28 April 2014; revised 28 May 2014; accepted 5 June 2014
ABSTRACT
Digital imaging techniques have enabled to gain insight into complex structure-functional processes involved in the neo-cortex maturation and in brain development, already recognized in anatomical and histological preparations. Despite such a refined technical progress most diagnostic records sound still elusive and unreliable because of use of conventional morphometric approaches based on a unique scale of measure, inadequate for investigating irregular cellular components and structures which shape nervous and brain tissues. Instead, these could be efficiently analyzed by adopting principles and methodologies derived from the Fractal Geometry. Through his masterpiece, The Fractal Geometry of Nature [1], Benoît Mandelbrot has provided a novel epistemological framework for interpreting the real life and the natural world as they are, preventing whatever approximation or subjective sight. Founded upon a body of well-defined laws and coherent principles, the Fractal Geometry is a powerful tool for recognizing and quantitatively describing a good many kinds of complex shapes, living forms, organized patterns, and morphologic features long range correlated with a broad network of functional interactions and metabolic processes that contribute to building up adaptive responses making life sustainable. Scale free dynamics characterized biological systems which develop through the iteration of single generators on different scales thus preserving proper self-similar traits. In the last decades several studies have contributed to showing how relevant may be the recognition of fractal properties for a better understanding of brain and nervous tissues either in healthy conditions or in altered and pathological states.
Keywords:Complexity, Fractal Dimension, Self-Similarity, Healthy and Diseased Brain Tissues

1. Introduction
The original conception of Galeno (Pergamo, 129-216 D.C.) that confined the superior functions of brain within three cerebral cells (spheres), has spanned several centuries up to the Renaissance period culminating with Leonardo da Vinci (1452-1519). As pointed out in an exhaustive review [2] the first outstanding breakthrough in the brain knowledge was accomplished by Andreas Vesalius with his famous work De humani corporis fabrica (1543); cerebral surface convolutions were described without a detailed identification of inner morphological pattern. Nevertheless he conjectured a probable link between brain structures and psychological functions. Relevant investigations were successively performed by Marcello Malpighi (1628-1694) who suggested the existence of a nervous fluid filling in cerebral glands, by Thomas Willis (1621-1675) who evidenced an arterial circuit by anastomosis of internal carotids and vertebral artery, and by Vicq d’Azyr (1746-1796) who revealed convolutions in unidentified areas of the external brain surface. Albrecht von Haller (1708-1777) had also conjectured the existence of a secretive function. Franz Joseph Gall (1758-1828) and Johann Spurzheim (1776- 1828) identified phrenological maps sharing specific brain functions. Later on, Paul Broca (1824-1880) localized cerebral zones with specialized activities such as those for the language arguing that “Nous parlons avec l‘hémisphère gauche”. Carl Wernicke (1848-1905) identified an area in the temporal lobe, whose damage may provoke the selective loss of the capacity of listening words. Within the late 19th and the early 20th centuries, there appeared the outstanding contributions of two coeval scientists, both Nobel Laureates: Camillo Golgi (1843-1926) [3] who postulated the “reticular theory” suggesting that the nervous system is a syncytial system of nervous fibers that forms an intricate complex diffuse network along which the nervous impulse may propagate. On the other side Santiago Ramón y Cajal (1852-1934) [4] developed the “neuron theory” for which the relationship between nerve cells was not one of continuity, but rather of contiguity, accomplished through small membranous spines protruding from neuron’s dendrites that typically receive input from a single synapse of an axon (output). In the last decades the implementation of performant imaging techniques, such as Positron Emission Tomography (PET), Functional Magnetic Nuclear Resonance (fMNR), Computed Axial Tomography (CAT), etc. in concomitance with the increment of the theoretical knowledge provided by the innovative Fractal geometry and modern Mathematics have opened an unprecedented analytical breakthrough. Actually, digital images recovered from body sequential cross-sections according to the Cavalieri’s principle for volume determination enabled to combine the close representation of whole structures with the fractal quantitative evaluation of their anatomical/morphological peculiarities [5] .
2. Main Properties of Fractal Elements
According to Mandelbrot “A fractal set is a set in metric space for which the Hausdorff-Besicovitch dimension D is greater than the topological dimension DT” [6] [7] . In nature, a fractal object is defined by its structural properties, namely by surface roughness, intricacy, irregularity and absence of smoothness, form invariance, geometrical or statistical self-similarity, power law scaling, morpho-functional complexity, iteration of simple generators, expressed by a numerical descriptor i.e. a fractional/non-integer dimension. The Richardson-Mandelbrot equation provides the mathematical basis for understanding geometric and spatial fractal structures, and for measuring and interpreting them, namely:
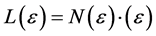 (a)
(a)
where L(ε) represents the contour length (e.g. the perimeter) of the biological component under investigation, (ε) the unit length of measure, and N(ε) the number of unit lengths (ε) needed to cover the contour L(ε). By substituting N(ε) with [loD∙ε−D] into (a), the above equation can be transformed by logarithmic procedure and rewritten as:
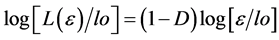 (b)
(b)
which is the equation of a straight line with slope 1 − D, and from which the dimensional exponent D can be calculated to yield the numerical value of the fractal dimension FD. FD is a statistical non-integer quantity that gives a measure of the geometric complexity of an irregular shape. lo is a reference scale without influence on the determination of D. In contrast to deterministic geometric fractals, biological structures and objects observed in Nature are self-similar within a limited range of scales. Only within this scale interval or scaling window can the scale-invariant (fractal) properties of an irregular object of finite size be observed. A real “fractality” exists when the experimental scaling range covers at least two orders of magnitude, although fractality over many orders of magnitude has been observed in various natural fields. The fractal window characterizing biological and natural fractals also called “bi-asymptotic fractals” is graphically represented by the region II in the middle of three typical regions, limited by a lower (εmin) and an upper bound (εmax) of the bi-asymptotic curve, where a straight line can be drawn and the fractal dimension [FD] calculated from its slope as documented in a previous report [8] . Defining a “scaling range” appears an inescapable requisite for assessing the fractality of every biological element (Figure 1). While the practical evaluation of the fractal dimension could be obtained by various quantitative approaches, the most reliable method is by far the box counting easily based on counting of the non-empty boxes N at a variable grid length (ε) [9] . It is obvious that the fractal theory is in opposition to the ancient, conventional vision based on Euclidean geometry and to its widely adopted concepts, such as homeostasis, linearity, smoothness, and thermodynamic reversibility, which stem from a more intuitive but artificially ideal view of reality. In the chapter of his work entitled “Epilog: The Path to Fractals”, Benoit Mandelbrot wrote: “The reader knows well that the probability distribution of fractals is hyperbolic, and that the study of fractals is rife with other power law relationships”. Although Mandelbrot’s famous seminal essay on statistical self-similarity and fractal dimension dates back to 1977 [6] and the first coherent paper on fractal geometry was published even earlier [7] , it is worth here recalling that the “heuristic introduction” of this innovative discipline or, more vividly expressed “the irruption of fractal geometry” [10] into the life sciences such as biology and medicine actually took place in the early eighty years of the last century as documented by the seminal paper of Paumgartner et al. [11] . A critical review of fractal concepts was recently addressed encompassing the definition of dimensional imbalance, the modified capacity dimension and the analytical calculation of its value, the relationship with the scaling exponent, and showing that such a definition satisfies basic demands of physics, before all the dimensional balance in mathematical equations used in applied sciences. Since some concepts in fractal geometry are determined descriptively and/or qualitatively, the paper provides their exact mathematical definitions and explanations, including the Richardson’s coastline method [12] .
3. The Complexity of Human Brain and Nervous Cells
The evolutionary concourse of two major events, “the tremendous expansion and the differentiation of the neocortex”, as reported by De Felipe [13] has contributed to the development of the human brain. Today, modern neurosciences recognize the presence of fractal properties in brain at various levels, i.e., anatomical, functional, pathological, molecular, and epigenetic, but not so long ago there was no analytical method able to objectively
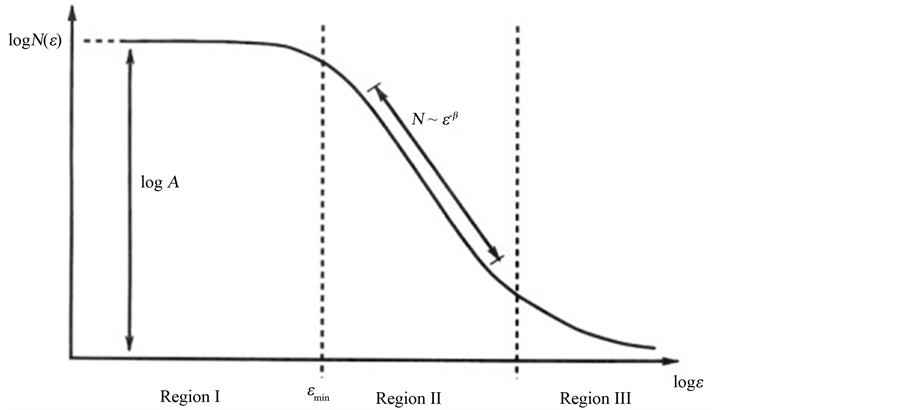
Figure 1. The three typical regions of an asymptotic fractal. Asymptotic natural or biological fractals only show autosimilar scaling properties (fractality) within a fractal window, represented by the Region II, limited by a lower (εmin) and an upper bound (εmax) limit (dashed lines), where a straight line can be drawn and the fractal dimension calculated from its slope, using the logarithmic relation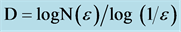 . From: [9] Döllinger, J.W., Metzler, R. and Nonnenmacher, T.F. (1998) Bi-asymptotic fractals: fractals between lower and upper bounds. J Physics A: Math Gen, 31, 3839-3847.
. From: [9] Döllinger, J.W., Metzler, R. and Nonnenmacher, T.F. (1998) Bi-asymptotic fractals: fractals between lower and upper bounds. J Physics A: Math Gen, 31, 3839-3847.
describe the complexity of biological systems such as the brain. The intricacy of mammalian brain folds led Mandelbrot to argue that “A quantitative study of such folding is beyond standard geometry, but fits beautifully in fractal geometry”. At that time however, there was no certainty about the brain’s geometry or about neuron branching. Anatomical-histological evidence that the complexity of the plane-filling maze formed from dendrites of neural Purkinje cells of cerebellum was more reduced in non-mammalian species than in mammals led Mandelbrot to comment: “It would be very nice if this corresponded to a decrease in D (fractal dimension), but the notion that neurons are fractals remains conjectural” affirmed Mandelbrot [1] . Since then, a wealth of investigations have documented the fractal organization of the brain and nervous tissue system, and implication of fractals for neurosciences has been unambiguously affirmed. Among the first applications of fractal analysis to nervous and brain tissue were the pioneering studies of Smith et al. [14] . These authors showed that the fractal dimension is an unbiased measure of the complexity of neuronal borders and branching pattern and of the time course of morphological development and differentiation of spinal cord neurons in culture, increasing from 1.1 for the less differentiated neuron up to 1.5 for the most differentiated cell [15] . Further studies have confirmed that the fractal dimension correlates with the increase in morphological complexity and neuronal maturity [16] [17] . The brain consists of distinct anatomical areas formed by nervous tissue mainly composed of neurons and glial cells of distinct types. Neurons contain the axon (a long cytoplasmic process associated with the cell body, which communicates with target organs), and the dendrites (shorter cytoplasmic processes off the cell body which allow communication between neurons), while glial cells of various types exert a structural and a physioimmunologic role as a net via their branched and unbranched protoplasmic processes [18] . These anatomical, morphological, and physiological properties combine to create the brain’s complexity, which can eventually be modeled by a supercomputer as recently proposed [19] [20] . While three-dimensional digital reconstructions of axonal and dendritic branching are indispensable for exploring neural function, the computational approaches enabled to quantify the intricate relationship between neuronal morphology (structure) and physiology (activity). Although the importance of neuronal morphology has been recognized since the early days of neuroscience, one is obliged to deplore that conventional morphometric approaches inspired to Euclidean geometry were still applied for investigate morphological properties of complex shapes. Suffices it to mention a recent study reporting that cortical neurons and other nervous cells were classified on the basis of their somata identified through an approximated unrealistic shape format, i.e. triangular, round, and oval by conventional geometry [21] . Nowadays, increasing efforts are directed to elucidate the functional roles of axonal and dendritic arbors in synaptic integration, signal transmission, network connectivity, and circuit dynamics which in turn require quantitative analyses of digital three-dimensional reconstructions. Reconstructing complex neuronal branching in digital 3D format may help map brain circuitry with its billions of connections. [22] . One of the most ubiquitous laws in biology is allometric scaling whereby a biological variable Y is related to the mass of the organism by a power law
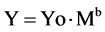
where b is the so called allometric exponent. Empirical observations and recent researches showed that the basal metabolic rate of all organisms is allometric scaled by a factor of 2/3 or 3/4. A recent report on quantitative morphological parameters and cell phenotype showed that the metabolic rate R for all organisms follows exactly the 3/4 power-law of the body mass, i.e., R ~ M3/4. Such a relation holds equally well from the smallest bacterium to the largest animal and remains valid even down to the individual components of a single cell, such as the mitochondrion, and the respiratory complex [23] . Furthermore, the fractal geometry leads to a 4/5 allometric scaling law, endowing human brain with a fifth dimension [24] . In a review article dealing with conceptual implications of fractals in nervous system it has been argued that power-law scaling and other characteristics of fractal and self-similar patterns in space and/or time can be identified at all levels of neural organization [25] .
4. Apoptosis in Nervous Cells
Apoptosis is the principal mechanism by which cells are physiologically eliminated from living tissues and organisms. Those cells that are committed to die undergo morphological changes that are mostly irreversible [26] . During apoptotic death, cells are neatly carved up by caspases and packaged into apoptotic bodies as a mechanism to avoid immune activation [27] . Nevertheless it is difficult to recognize cellular death in solid tissues, first because apoptosis takes place rapidly in vivo and second apoptotic cells are efficiently removed by phagocytic elements so that it reduces to a residual process [28] . Unfortunately, available methods do not adequately identify the early phases of apoptosis when the cells appear overtly normal and healthy. Therefore studies directed to quantitatively describe early events of apoptosis such as changes of transcription factors, rearrangements of chromatin nuclear patterns and membrane ultrastructure are needed, particularly in human nervous cells. In experimentally induced apoptosis of epithelial and other non-lymphoid cells, fine ultrastructure changes were measured in nuclear chromatin, internal perinuclear membrane and in cellular plasmalemma still at the beginning of apoptosis. These changes, i.e. the morphological reorganization of chromatin texture within nuclear domains and other membrane organelles were more pronounced in human SK-BR-3 breast cancer cells treated with an apoptogenic ionophore [29] and measurable by fractal and GLCM analyses at the early stage of apoptosis, well before the onset of conventional cellular markers and enzymatic events. This ultrastructural loss hit most cell and tissue components and was documented by a fractal dimension (FD) lower than in active living elements [30] . Data from human glioma cells indicated that ultrastructural cell complexity determined by fractal dimension and lacunarity significantly decreased after the UV irradiation. The nuclear lacunarity is a particularly sensitive parameter in detecting early apoptosis, while the fractal analysis enables to discover cellular apoptotic changes earlier than conventional flow cytometric analysis [31] . Even in Alzheimer’s disease neuronal apoptosis is difficult to assess because only a limited number of apoptotic neurons can be detected, confirming results of studies mentioned above [32] . Fractal analysis of rat neurons revealed that 24 hours long incubation with a flavonoid quercetin diminished neuronal arborisation in cortical neurons and increased sensitivity to cell death [33] . Neurons of the hippocampal dentate gyrus selectively undergo programmed cell death in patients suffering from bacterial meningitis and in experimental models of pneumococcal meningitis in infant rats [34] . Elucidation of early phases of cell death in neuronal cells will enable to quantify ultrastructural changes most undetectable at a visual and microscopic inspections caused by drugs and other chemotherapeutic agents [29] , and/ or by early onset of neurodegenerative diseases [25] .
5. Healthy and Diseased Brain Tissues
Fractal analysis was applied to anatomical/ histological images and high-resolution magnetic resonance images in order to quantify the developmental complexity of the human cerebral cortex, the alterations in diseased brain with epilepsy, schizophrenia, stroke, multiple sclerosis, amyotrophic lateral sclerosis (ALS), cerebellar degeneration, and the morphological differentiation of the peripheral nervous system [25] . FD values estimated for brain white matter (WM) skeleton, surface and general structure in both controls and ALS patients revealed no significant WM changes between controls and ALS patients and among the AL5 subgroups [35] . A highly significant reduction of the fractal dimension was observed in the cortical ribbon of Alzheimer’s Disease patients with respect to control subjects [36] . The 3-D fractal dimension of the Purkinje neurons decreased from 1.723 to 1.254, indicating a significant reduction of dendritic complexity during cortical development disorder [37] . The fractal analysis has enabled to quantitatively describe the complex morphological forms in which astrocytes occur in brain of ischemic/hemorrhagic stroke and Alzheimer’s disease (AD) [38] . In contrast, fractal dimension values were found higher in the Gray Matter (GM) of Multiple Sclerosis patients (MS) compared to controls and indicated that GM tissue in MS has higher morphological complexity, perhaps due to the presence of the inflammatory component (i.e. microglia activation) and cellular changes in the GM [39] . In the normal human retina, blood vessels or vascular trees exhibited an FD of 1.7, the same fractal dimension found for a diffusionlimited growth process, a finding which may have implications for the understanding of the embryological development of the retinal vascular system. [40] . In large neurons of the human dentate nucleus the fractal dimension [FD] has been found to correlate with the increase in morphological complexity and neuronal maturity [41] . Lastly, a quantitative evaluation of the surface fractal dimension may allow not only to measure the complex geometrical architecture, but also to model the development and growth of tumor neo-vascular systems and explore the morphological variability of vasculatures in nature, in particular the microvasculature of normal and adenomatous pituitary tissue [42] . The fractal analysis was recently applied on patients with cerebral arterial venous malformations (AVM): increased FD values related to structural vascular complexity were due to the increased number of feeding arteries in patients suffering from AVM [43] . A recent study described a method for quantifying cerebral blood flow (CBF) distribution in Alzheimer’s disease (AD) from SPECT images obtained with 99mTc-hexamethylpropyleneamine oxime (HMPAO) by 3-dimensional fractal analysis (3D-FA). The fractal dimension 3D-FA was well correlated with the cognitive impairment, as assessed in neuropsychological tests and could represent a useful method for objectively evaluating the progression of AD [44] .
6. Fractal Design from Genome to Neurons
The relevant question linking fractal and gene design was adumbrated some years ago in these terms: “Do genes contain fractal algorithms?” [45] . It appears as much interlocutory because genes are DNA entities codifying constructive units or templates, while fractal algorithms represent mechanisms (iteration, self-organization, environmental epigenetic constraints, etc.) the nature may eventually adopt in order to assemble self-similar dynamic units into final shapes. Whether genes do or do not determine biological shape directly or by following stochastic and environmental effects, either through a selective activation driven by transcription factors [46] or by fractal mechanisms will be a matter of a controversial debate. Mandelbrot addressed by a fractal model of the guinea pig Purkinje neurons the link between fractality of the genome with the fractality of organelles and neurons [1] . Recently the FractoGene “cause and effect” concept conceived that “fractal genome governs fractal growth of organelles, organs and organisms” [47] . The Principle of this recursive genome function (PRGF) breaks through the double lock of central dogma and junk DNA barriers [48] . Decades of computer modeling of neurons and neuronal networks suggested that the amount of information necessary to build just a tiny fraction of the human body, i.e. just the cerebellum of the nervous system, was a task for which the 1.3% of the information that the genome could contain was just totally insufficient [49] . The hierarchical organization of the brain, seen at multiple scales from genes to molecular networks to building neurons organized in microand macronetworks, has a fractal structure as well, with various modules which are interconnected in a small-world topology [50] .
7. Conclusion
While the complexion of brain and nervous tissue remains largely understood, the present survey provides experimental data confirming that biological processes including growth, proliferation, apoptosis, epigenetic and genetic mechanism, morphologic/ultrastructural and functional organization occurring in living shaped elements and complex structured tissues may follow fractal rules. The large agreement with the fractal nature of the brain and nervous cell system sustained by theoretical, experimental and heuristic foundations is nowadays consolidated and intervenes more than thirty years after the publication of the Fractal Geometry of Nature, in which Mandelbrot recognized that “the notion that neurons are fractals remains conjectural”. Its relevance and contribution to the cultural development of mankind (as comprehensive of humanistic and scientific thinking) is keen underlined by the observation of some years ago arguing that the fractal geometry could be considered as a biological design principle for living organisms [45] . Among the main fractal peculiarities worth noticing is the process of iteration, whose powerful dynamics allows specific generators to be properly iterated at different scales (small and large) without an a priori choice, by linking efficient genetic programming in order to achieve the formation of viable biological forms and living objects [51] .
References
- Mandelbrot, B. (1983) The Fractal Geometry of Nature. Freeman, San Francisco.
- Piccolino, M. (2008) Neuroscienze Controverse: Da Aristotele alla moderna scienza del linguaggio. Bollati Boringhieri, Torino.
- Golgi, C. (1903) Sulla fine anatomia degli organi centrali del sistema nervoso. Opera Omnia, I, Hoepli Milano.
- Ramón and Cajal, S. (1888) Sobre las fibras nerviosa de la capa molecular del cerebelo. Rev. Trim. Histol. Normal. Patology, 2.
- Losa, G.A. (2012) Fractals in Biology and Medicine. In: Meyers, R., Ed., Encyclopedia of Molecular Cell Biology and Molecular Medicine, Wiley-VCH Verlag, Berlin, 1-25.
- Fractals, M.B. (1977) Form, Chance and Dimension. W.H. Freeman & Company, San Francisco.
- Mandelbrot, B. (1967) How Long Is the Coast of Britain? Statistical Self-Similarity and Fractional Dimension. Science, 155, 636-640. http://dx.doi.org/10.1126/science.156.3775.636
- Losa, G.A. and Nonnenmacher, T.F. (1996) Self-Similarity and Fractal Irregularity in Pathologic Tissues. Modern Pathology, 9, 174-186.
- Döllinger, J.W., Metzler, R. and Nonnenmacher, T.F. (1998) Bi-Asymptotic Fractals: Fractals between Lower and Upper Bounds. Journal of Physics A: Mathematical and General, 31, 3839-3847. http://dx.doi.org/10.1088/0305-4470/31/16/012
- Belaubre, G. (2006) L’irruption des Géométries Fractales dans les Sciences. Editions Académie Européenne Interdisciplinaire des Sciences (AEIS), Paris.
- Paumgartner, D., Losa, G.A. and Weibel, E.R. (1981) Resolution Effect on the Stereological Estimation of Surface and Volume and Its Interpretation in Terms of Fractal Dimensions. Journal of Microscopy, 121, 51-63. http://dx.doi.org/10.1111/j.1365-2818.1981.tb01198.x
- Ristanovic, D. and Losa, G.A. (2013) A Contribution to Definitions of Some Fractal Concepts. The Fractal Laboratory Journal, 2, 2-14.
- De Felipe, J. (2011) The Evolution of the Brain, the Human Nature of Cortical Circuits, and Intellectual Creativity. Frontiers iNeuronatomy, 5, 1-17.
- Smith, T.G., Marks, W.B. and Lange, C.D. (1989) A Fractal Analysis of Cell Images. Journal of Neuroscience Methods, 7, 173-180. http://dx.doi.org/10.1016/0165-0270(89)90100-3
- Smith, T.G. (1994) A Fractal Analysis of Morphological Differentiation of Spinal Cord Neurons in Cell Culture. In: Losa, et al., Eds., Fractals in Biology and Medicine, Birkhäuser Press, Basel, 20-25. http://dx.doi.org/10.1007/978-3-0348-8501-0_18
- Milosevic, N.T. and Ristanovich, D. (2006) Fractality of Dendritic Arborization of Spinal Cord Neurons. Neuroscience Letters, 396, 172-176. http://dx.doi.org/10.1016/j.neulet.2005.11.031
- Bernard, F., Bossu, J.L. and Gaillard, S. (2001) Identification of Living Oligodendrocyte Developmental Stages by Fractal Analysis of Cell Morphology. Journal of Neuroscience Research, 65, 439-445. http://dx.doi.org/10.1002/jnr.1172
- Losa, G.A. (2012) Fractals and Their Contribution to Biology and Medicine. Medicographia, 34, 365-374.
- Markram, H. (2006) The Blue Brain Project. Nature Reviews Neuroscience, 7, 153-160. http://dx.doi.org/10.1038/nrn1848
- De Felipe, J. (2011) The Neocortical Column. Frontiers in Neuroanatomy, 5, 1-16.
- Romand, S., Wang, Y., Toledo-Rodriguez, M. and Markram, H. (2011) Morphological Development of Thick-Tufted Layer V Pyramidal Cells in the Rat Somatosensory Cortex. Frontiers in Neuroanatomy, 45, 1-25.
- Halavi, M., Hamilton, K.A., Ruchi, P. and Ascoli, G.A. (2012) Digital Reconstructions of Neuronal Morphology: Three Decades of Research Trends. Frontiers in Neuroscience, 6, 1-11. http://dx.doi.org/10.3389/fnins.2012.00049
- Bizzarri, M., Pasqualato, A., Cucina, A. and Pasta, V. (2013) Physical Forces and Non-Linear Dynamics Mould Fractal Cell Shape. Quantitative Morphological Parameters and Cell Phenotype. Histology and Histopathology, 28, 155-174.
- He, J.H. and Zhang, J. (2004) Fifth Dimension of Life and the 4/5 Allometric Scaling Law for Human Brain. Cell Biology International, 28, 809-815.
- Werner, G. (2010) Fractals in the Nervous System: Conceptual Implications for Theoretical Neuroscience. Frontiers in Physiology, 1, 15-30.
- Wyllie, A.H., Kerr, J.F.R. and Currie, A.R. (1980) Cell Death: The Significance of Apoptosis. International Review of Cytology, 68, 251-306. http://dx.doi.org/10.1016/S0074-7696(08)62312-8
- Edinger, A.L. and Thompson, C.B. (2004) Death by Design: Apoptosis, Necrosis and Autophagy. Current Opinion in Cell Biology, 16, 663-669. http://dx.doi.org/10.1016/j.ceb.2004.09.011
- Losa, G.A. and Graber, R. (1998) Apoptotic Cell Death and the Proliferative Capacity of Human Breast Cancer. Analytical Cellular Pathology, 16, 1-10.
- Castelli, C., Nonnenmacher, T.F. and Losa, G.A. (2002) Morphofractal Reorganization of Plasma Membrane and Nuclear Components during the Apoptosis of Breast Cancer Cells. In: Losa, G.A., Ed., Fractals in Biology and Medicine, Mathematics and Biosciences in Interaction, Birkhäuser, Basel, 67-76.
- Losa, G.A. and Castelli, C. (2005) Nuclear Patterns of Human Breast Cancer Cells during Apoptosis: Characterization by Fractal Dimension and Co-Occurrence Matrix Statistics. Cell and Tissue Research, 322, 257-267. http://dx.doi.org/10.1007/s00441-005-0030-2
- Pantic, I., Harhaji-Trajkovic, L., Pantovic, A., Milosevic, N.T. and Trajkovic, V. (2012) Changes in Fractal Dimension and Lacunarity as Early Markers of UV-Induced Apoptosis. Journal of Theoretical Biology, 303, 87-92. http://dx.doi.org/10.1016/j.jtbi.2012.03.013
- Yuan, J. and Yankner, B.A. (2000) Apoptosis in the Nervous System. Nature, 407, 802-809. http://dx.doi.org/10.1038/35037739
- Jakubowicz-Gil1, J., Rzeski, W., Zdzisinska, B., Dobrowolski, L. and Gawron, P.A. (2008) Cell Death and Neuronal Arborization upon Quercetin Treatment in Rat Neurons. Acta Neurobiologiae Experimentalis, 68, 139-146.
- Gianinazzi, C., Grandgirard, D., Simon, F., Imboden, H., Joss, P., Täuber, M. and Leib, S.L. (2004) Apoptosis of Hippocampal Neurons in Organotypic Slice Culture Models: Direct Effect of Bacteria Revisited. Journal of Neuropathology & Experimental Neurology, 63, 610-617.
- Rajagoplan, V., Liu, Z., Allexandre, D., et al. (2013) Brain White Matter Shape Changes in Amyotrophic Lateral Sclerosis (ALS): A Fractal Dimension Study. PLoS ONE, 8, Article ID: e73614.
- King, R.D., Brown, B., Hwang, M., Jeon, T. and George, A.T. (2010) Fractal Dimension Analysis of the Cortical Ribbon in Mild Alzheimer’s Disease. NeuroImage, 53, 471-479. http://dx.doi.org/10.1016/j.neuroimage.2010.06.050
- Kim, J., Kwon, N., Chang, S., Kim, K.T., Lee, D., Kim, S., Yun, S.J., Hwang, D., Kim, J.W., Hwu, Y., Margaritondo, G., Je, J.H. and Rhyu, I.J. (2011) Altered Branching Patterns of Purkinje Cells in Mouse Model for Cortical Development Disorder. Scientific Reports, 1, 122-128.
- Pirici, D., Mogoanta, L., Margaritescu, O., et al. (2009) Fractal Analysis of Astrocytes in Stroke and Dementia. Romanian Journal of Morphology and Embryology, 50, 381-390.
- Esteban, J., Sepulcre, J., de Miras, J.R., Navas, J., de Mendizábal, N.V., Goñi, J., Ma Quesada, J., Bejarano, B. and Villoslada, P. (2009) Fractal Dimension Analysis of Grey Matter in Multiple Sclerosis. Journal of the Neurological Sciences, 282, 67-71. http://dx.doi.org/10.1016/j.jns.2008.12.023
- Masters, B.R. (2004) Fractal Analysis of the Vascular Tree in the Human Retina. Annual Review of Biomedical Engineering, 6, 427-452. http://dx.doi.org/10.1146/annurev.bioeng.6.040803.140100
- Milosevic, N.T., Ristanovic, D., Jelinek, H.F. and Rajkovic, K. (2010) Morphology and Cell Classification of Large Neurons in the Adult Human Dentate Nucleus: A Quantitative Study. Neuroscience Letters, 468, 59-63. http://dx.doi.org/10.1016/j.neulet.2009.10.063
- Di Ieva, A. (2010) Angioarchitectural Morphometrics of Brain Tumors: Are There Any Potential Histopathological Biomarkers. Microvascolar Research, 80, 522-533. http://dx.doi.org/10.1016/j.mvr.2010.09.005
- Reishofer, G., Koschutnig, K., Enzinger, C., Ebner, C. and Ahammer, H. (2012) Fractal Dimension and Vessel Complexity in Patients with Cerebral Arteriovenous Malformations. PLoS ONE, 7, Article ID: e41148. http://dx.doi.org/10.1371/journal.pone.0041148
- Nagao, M., Murase, K., Kikuchi, T., Ikeda, M., Nebu, A., Fukuhara, R., Sugawara, Y., Miki, H. and Ikezoe, J. (2001) Fractal Analysis of Cerebral Blood Flow Distribution in Alzheimer’s Disease. Journal of Nuclear Medicine, 42, 1446-1450.
- Weibel, E.R. (1991) Fractal Geometry: A Design Principle for Living Organisms. American Journal of Physiology, 261, 361-369.
- Honda, H. (1999) Genes Do Not Determine Biological Shape Directly. Forma, 14, 287-293.
- Pellionisz, A.J. (2012) The Decade of FractoGene: From Discovery to Utility-Proofs of Concept Open Genome-Based Clinical Applications. International Journal of Systemics, Cybernetics and Informatics, 17-28.
- Pellionisz, A. (1989) Neural Geometry: Towards a Fractal Model of Neurons. Cambridge University Press, Cambridge.
- Pellionisz, A. (2008) The Principle of Recursive Genome Function. Cerebellum, 7, 348-359. http://dx.doi.org/10.1007/s12311-008-0035-y
- Agnati, L.F., Guidolin, D., Carone, C., Dam, M., Genedani, S. and Fuxe, K. (2008) Understanding Neuronal Molecular Networks Builds on Neuronal Cellular Network Architecture. Brain Research Reviews, 58, 379-399. http://dx.doi.org/10.1016/j.brainresrev.2007.11.002
- Di Ieva, A., Grizzi, F., Jelinek, H., Pellionisz, A.J. and Losa, G.A. (2013) Fractals in the Neurosciences, Part I: General Principles and Basic Neurosciences. The Neuroscientist. [Epub ahead of print]