Neuroscience & Medicine
Vol.2 No.3(2011), Article ID:7398,17 pages DOI:10.4236/nm.2011.23023
Cellular Perception: When the Cell Model Includes a Sense Order which Ensues from a Philosophy of Nature, the Signaling and Epigenetics Effects which Can Result from Exposure to Magnetic Fields Are Described Better
![]()
Algology Department, Soissons General Hospital, Soissons, France.
Email: pierre.lechapellier@ch-soissons.fr
Received February 9th, 2011; revised May 11th, 2011; accepted July 20th, 2011.
Keywords: Philosophy of Nature, Aporia, Organicism, Sense Order, Causal Order, Cell, Epigenetics, Static Magnetic Fields, SMF, Electromagnetic Fields, EMF, Topological Thermodynamics, Extracellular Aqueous Medium
ABSTRACT
Academic biology-medicine refers to a couple of philosophies, Organicism and Mechanism, which translates into an association of Cybernetic diagrams and molecular Reductionism. This association presents logical difficulties which make it unsuitable to describe correctly biological effects of electromagnetic fields, EMF. But these logical difficulties may be overcome when renewing the organic cell idea by means of a Philosophy of Nature which juxtaposes causality order and sense order in the cell. The signalsome, the set of descriptive components resulting from the genome, is constantly reorganized. This remodeling may become epigenetic when the phenotype becomes transformed by experience of perceptions in a given medium, because the perception of overall information coming from the extracellular medium becomes functional within the system. In that cellular perception, it is stated that the significance base which contributes to the sense order results from the qualitative topological structure of the extracellular medium. Therefore the EMF interactions target is not only the membrane and its molecules; it is also the structure of the extracellular medium which bathes the membrane. Knowing that the sense order modulation constitutes the global soil of the (localized) causality order, it is possible to obtain a same EMF bioeffect on a membrane molecule by treating a culture of cells in its bath or by treating only the extracellular aqueous medium. Consequently, the double bioeffect resulting from EMF exposure is described simply, because the sense order, such as it results from the qualitative structuring of the medium, forms the significance base which directs the causal mechanics of the cellular answer.
1. The Two Aporias of the Biology-Medicine Usual Model
1.1. Static Magnetic Fields Induce Bioeffects in Living Cells, which Must be Explained
Healthy cells in living organisms are widely transparent to Static Magnetic Fields, SMF: Reports indicated transient but not harmful effect under 4T1 [1]. But cell shape and plasma membrane may be altered, generally in a way able to restore the cell life, after exposure of chemically hit cells to SMF, 6 mT [2]. Moreover, the growth speed of phytopathogen microscopic fungi, depending of the kind of fungi, may vary around +/– 10% under exposure to 0.1 - 1 mT [3]. How a magnetic field exposure can induce such qualitative bioeffects in living cells? Pulsed electromagnetic fields, PEMF, induce weak electric currents which are supposed to mediate the healing processes. But these currents are absent under a SMF exposure bringing similar bioeffects. Infrared spectroscopy showed recently that that the largest changes in the cells exposed to SMF were found in hydrogen bonds, H-bonds, and more commonly in water [4]. How could some changes in Hbonds of water outer the cell, going to fleeting relaxation times and being under thermal agitation, produce bioeffects?
1.2. Reductionism and Cybernetics
We know that the two philosophies of the biological nature of life which are used by science are Organicism and Mechanism. Usual Organicism gave birth to Cybernetic descriptions of interactions within biosystems and to a doctrine called Holism. Mechanism led to deriving the organism vitality and conscience from relations between its constitutive molecules. This mechanism approach thus translated into Reductionism, since life analysis is reduced to mechanical descriptions of biomolecules and their interactions. Reductionism doctrine which is dominant in biology-medicine, is thus unaware of the role of the sense order and it doesn’t take account about event’s significance in an irreducible living being. That is the reason why Reductionism poorly understands the biological effects of electromagnetic fields, EMF, when its descriptions are limited to movements of electro-charged molecules, i.e. to current density induced by applied EMF, (Liboff, 2004, [5]). Moreover, as Reductionism doctrine is based on Cartesian dualism which operated a rupture between a real objective part and a phenomenal subjective part, Reductionism doctrine presents an aporia2. And the Cybernetic model which supplements it and which allows the explanatory diagrams, also presents an aporia3.
In spite of these two aporias, Reductionism approach got a great success and produces a very large knowledge of the molecular components of the biosystems. Also known as Atomism, Reductionism has not got any reference to Atomist philosophy of Democritus. However in Democritus theory, physical vacuum played an essential part, in relation to the varieties of forms and changes. So the role of the bioelectromagnetic interactions space, i.e. the role of the physical vacuum located between biomolecules, does not have any place in the Reductionist doctrine, especially in relation to the varieties of forms and changes. Can there be a relationship between the physical vacuum properties and the molecules forms and changes in the matter of a living organism? For example, we know hydrophobic forces fields present in the aqueous physical vacuum4 may affect the organic molecules form and curvatures directly. Reversely cointegration of biomolecules on the cell scale, then on the whole organism scale, is not entirely understood, particularly for genes and their products during embryo development, as Jacobs indicates it, (2003, [6]).
Such a biological understanding could be presumably facilitated if the two aporias of the usual Reductionist-Cybernetic model were overcome. That can be done by means of a new epistemological contribution resulting from the Philosophy of Nature. The naturalism of such a contribution can be useful not only for the bioelectromagnetic knowledge, but also for a biological comprehension leading to genesis of new therapies.
2. The Naturalism which Arises from the Philosophy of Nature
2.1. A Topological Immersion of the Reductionism in Biology
The living system functioning is not exhausted by a Reductionist description based on Cartesian dualism. However Cartesian dualism affirms that the objective reality of any material substance could appear in the space extension. The mathematician Rene Thom, (1980, [7]), thus proposed an immersion of Reductionism in its space extension, i.e., according to Thom, immersed in the morphology of the living systems. This space extension gives sense to the biochemical reaction which results in a conformation change, for a protein receptor as for a cell. And mathematics could abstract such a sense by means of topology5.
2.2. Topology as Situation Analysis
For the mathematician Henri Poincaré, (1895, [8]), the principles of topology correspond to a situation analysis, (Analysis Situs.) That leads to the following idea: If a sense order is inherent to a biological phenomenon constrained by morphology, it could be formalized by a combination between biochemistry and topology, (analysis of the situation constraints,) so that the aporia of the cybernetic diagram be overcome. That combination extends the reductionist model of biology, in which sense order and morphological significance are largely ignored. Organism is indeed described by Reductionism as being an interactive and integrated network of biochemical genes, proteins and reactions which gives birth to life6, as Chautard, (2009, [9]), indicates it. This interactive network is described by a topology of nodes and connections, without any morphological significance.
2.3. The Scientific Risk of a Naive Ontology
Insofar as the morphological idea of Thom results from its mathematical work relating to Cobordism, (i.e. to a tool for classifying differentiable manifolds, leading Thom to the idea of properties of form edges of topological varieties [11]), it has a major defect which is due to its philosophical weakness: The idea that forms edges could have a concrete dynamism corresponds to a naïve ontology7, which is naïve because contrary to Cartesian dualism. So on the one side, the idea of Thom had been rejected by reductionist philosophy. On the other side, it had not been integrated by philosophy of Nature, because it offers link neither with the existence in the world, nor with the irreducible principle which, according to the philosopher Maurice Merleau-Ponty, (1945, [12]), ensures the existence in the world of the living being. Since Thom topological approach does not describe form as a totality, it is not very useful for Holism8. That’s because, in biology-medicine, Holism is a philosophy of life which is based on totalities, to oppose, or supplement, to Reductionism, (Largeault, 2006, [13], Munson, 2007, [14]).
2.4. The Naturalist Theory of Cellular Perception
A topological idea has appeared in the qualitative theory of the differential equations, proposed by the mathematician Henri Poincaré, in 1890, [15,16]. In order to solve differential equations which were not integrable, Poincaré combined a local analytical information, observed in the vector field vicinity of the points of interest, with topological properties which depend on the overall structure. Such a combination idea can be taken up into biology of Cellular Perception:
The naturalist theory of cellular perception regards causal analytical information as being combined with topological properties. Analytical information corresponds to the signal transcription at the membrane level and its cascade transmission within the cell. The topological properties relate to the extracellular medium structure and its modulation by some events, depending on the cell intentionality. Such an approach allows a naturalist insertion of the sense order in cellular perception, through an epistemological reduction of the philosophy of Nature concepts:
2.5. The Essential Naturalist Idea of the Philosophy of Nature
The current philosophy of Nature has almost a century. The need for juxtaposing causal order and perceptive order appeared in the “Concept of Nature” of Whitehead, (1920, [17]) and its following texts [18-20]. It extended with the “Notes sur la Nature” of Merleau-Ponty, (1995, [21]), then with work of the current cross continuators, such as Barbaras, (2000, [22]), Toscano, (2003, [10]), Miquel, (2010, [23]). The whole of these philosophers leads to an essential idea: The existence at world and qualitative perception of a natural organism function through the topological structure of its being-envelope.
2.6. The Philosophy of Natural Organism and Its Being-Envelope
In order to suture logically the Nature wound opened by the Cartesian dualism, (between objective scientific construction and everything else considered as subjective,) Arnold Whitehead, (1925, [18]), stated that any natural being has an organism and that any organism has properties of endurance, stability, and extension.
The extension property destroys the idea of simple space-time localization. It applies to a feeling aspect which is described as an elementary equivalent of perception. It implies that any organism must have a beingenvelope to exist in the world, in which the external world events are structurally and gradually integrated. From the being-envelope in the way it affects the sense order, a constant modulation of the internal relations within the organism is caused. This constant internal modulation is what ensures the organism stability and adaptation to a changing world. It is also what makes possible the organism to be robust and enduring.
The philosophical idea about the being-envelope is concretely illustrated in biology by the Heat Shock Protein, HSP. When the cell feels there is a risky situation, for example a thermal risk, it synthesizes HSP which form model coats for proteins. These concrete being-envelopes of proteins will allow deformed proteins to find back their initial form once the stress has disappeared. The risk of tissues heating under exposure to EM waves leads the cells to synthesize HSP a long time before any consistent variation of the temperature, Blank, (2009, [24]), indicated. There is here an EMF qualitative impact.
2.7. The Concept of Cellular Organism
The application of the natural organism concept at the cell scale overcomes the Cartesian aporia, because it allows a logical association between the quantitative and qualitative signaling roles. In that view, the Cellular Perception associates the mechanical and quantitative role of a local signal with the non-mechanical and qualitative role of a delocalized9 overall information. This overall information starts from external events which gradually affect the cell being-envelope.
2.8. The Sense Order, between External Event and Organism Internal Relations
The epistemological reduction of being-envelope concept leads to the following double idea: The topological structure of the extracellular aqueous medium is modulated by certain worldly events. That topological structure change modulates the cell sense order, which affects and directs the internal relations of the cellular organism, (including epigenetic choices.) The sense order is defined as the qualitative soil where are growing the causal order effects resultant of quantitative signals. The sense order is logically what orients the cell active gene components choices:
3. Epigenetic Modulation of Signalsome
3.1. Signalsome Remodelling
While specializing, a cell selects the gene components being the best adapted to the cell function. It constitutes thus a signalsome, a set of signaling components resulting from genome. According to Berridge, (2008, [25]), each “signalsome” is constantly reorganized. This constant remodelling of the gene signalling components is what ensures the signalsome stability. It uses gene silencing [26], as well as disappearance and appearance of gene expressions in signalling networks. It is first translated by a controlled fluctuations damping, then stability.
Gene remodelling also tends to adjust permanently the signalsome properties so that the cell can face and answer to the request exerted by the external world. For example, Radman, (1999, [27]), observed the enzymes which are expressed according to the evolutionary changes. Radman showed that the three observable gene levels, (the individual genes, the gene populations relating to species, and the relations between the species sharing the same ecological niche,) function in a tangled up interaction. According to Miquel, (2010, [23]), this can be explained only because an overall information is made functional within the system, so complex it is, cell time after cell time. Such overall information can also lead to an epigenetic remodelling of the phenotype.
3.2. The Epigenetic Remodelling Pathway
The epigenetic remodelling pathway was defined by Berger, (2009, [28]). Issued from the organism life in a given world, it leads to a stable and inheritable phenotype by a change effect into a chromosome without alteration in the ADN sequence.
The epigenetic pathway is initiated by an extracellular information which comes from the external environment. It can’t take an elementary repercussion within the cellular organism unless an initiating protein translates the perceived information towards the establishment of a local chromatin context. The set of possible phenotype transformations issued from after the cell real-life is thus strongly constrained, Ho, (2009, [29]) indicated. Nevertheless, in partnership with the constant signalsome modulation, the epigenetic remodelling pathway testifies about a constant interaction between the genome and the environment.
This real interaction between organism-environment questions the reductionist philosophy of biology, as it is illustrated by an aphorism of Jacques Monod, quoted by Baulcombe, (2008, [30]): “What applies to a bacterium applies to an elephant.” The reciprocal aphorism is thus as follows: The fundamental principle of a biological interaction between organism and environment which applies to an elephant is also worth for a bacterium.
Now the Tsunami episode on 26 December 2004 showed that elephants have got a right perception of the forerunners signals of a natural event which could jeopardizes them. Signals started from subtle or abrupt changes in the wave’s environment [31].
This perception generated a salvation behaviour which was shared by most animals: As the vital danger was coming from the seaside, the elephants found enough monopolized power in themselves to break their chains and flee towards the safety of the heights, whereas the fish shoals have fled towards the safety of the open sea. An explanatory diagram according to the philosophy of Nature is represented on Figure 1.
Whatever for a bacterium or an elephant, being-envelope is supposed to have a four dimensions, 4D, qualitative structure. Those dimensions are the three of space
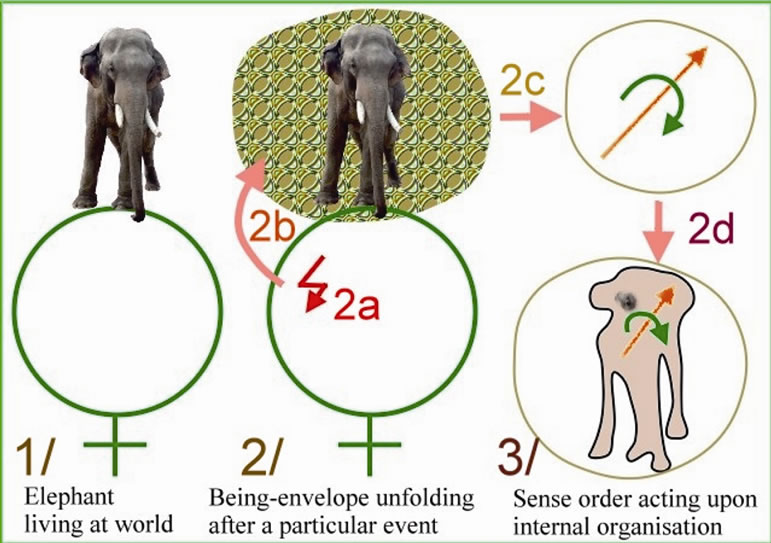
Figure 1. Philosophy of nature: organism at world: shows an explanation accorded to the Philosophy of Nature. In figure part 1, a natural organism as an elephant is living at world. In part 2a, a telluric event happens somewhere in this world, may be two thousand kilometres away. In 2b, because of its qualitative value, this event unfolds the being-envelope of the natural organism. In 2c, this external unfolding modulates the internal sense order. In 2d, modulation of sense order acts upon internal organization. Finally, in part 3, sense order modulation orients the internal activity.
plus the one of time equipped with some time thickness. Being-envelope structure is said to be unfolded by event with its qualitative thickness, including forerunners signals and possible effects.
In case of a sudden danger feeling, brutal unfolding of the being-envelope affects the sense order of the natural organism. It directs internal relations, which could allow monopolizing power in the salvation direction. But here, man as a human being is not a natural organism: He does not have this perceptive instinct depending on the quailtative environment, because the structured language is the house of its being in which he lives, as philosopher Heidegger, (1959, [32]), said. Nevertheless its organism cells are supposed to have, like all living cells, a perception and a joined epigenetic ability. This elementary perception has a lot more amplitude in its free individual cells than in its joined cells having a direct communication by gap junctions, as Ho [29], indicated. This cellular perception may be deduced by a reduction of what it is primarily for an elephant.
3.3. The Causal Order in Cellular Perception
In simple terms, any perception associates a causal order and a sense order.
The causal order in cellular perception is presented by systemic biology through signaling networks which mechanically ensure the detection of the cell micro-environment variations. According to the state and situation of the cell, this detection can be amplified, inhibited or modulated. To do this the majority of the signaling networks molecules may be activated or inhibited according to complex interactions. In that way Tan, (2008, [33]), indicates that gene expressions, located in the context of the biological signaling ways, are associated with the cellular regulation.
A principle of mechanical genes connectivity is illustrated on the left part of the Figure 2. Many gene expressions may be connected to proteins of the signaling networks and to pathophysiological status. For example, in the calcium signaling pathway, which sometimes functions independently of the genetic nucleus, the influx and efflux modulation of calcium ions, Ca2+, is qualitatively consistent with known transcriptional changes of ARN messenger, ARNm, as Wang, (2009, [34]), indicated. But Molecular Reductionism has no explanation about the overall “mechanism” allowing these cointegrated transcriptional changes. Based on philosophy of Nature the assumption is that the permanent modulation of the signalsome, related to the interaction between the cell and its environment, is directed by the sense order in the cellular life.
3.4. The Sense Order in the Cellular Life
According to the Philosophy of Nature presented by Barbaras, (2000, [22]) himself founded on Merleau-Ponty [21], the sense order in the life of a natural being such as a cell refers to a total form. This form which is never completely realized offers here and now the experience of qualitative virtualities which are present in the temporal thickness. The example of the elephant becoming aware of the imminence of a tidal wave which might submerge it shows the determining role of temporal thickness. The event to come causes a virtuality to be anticipated as a form by the natural organism. When modulating the sense order, this form causes an “instincttive” answer.
This idea may be epistemologically reduced from the elephant scale to the cell scale. In that way the cell beingenvelope could correspond to a given architectonic environment. A description of this architectonic environment can be found into qualitative features of the topological structure of the aqueous extracellular medium.
It is assumed that the cell sense order associated with the cell total form results as an oriented set of constraints exerted on the intracellular causal relations. This way, some external events may have a qualitative repercussion upon cell sense order, justifying a possible reorientation of the signalsome modulation, when these events affect qualitatively the topological structure of the extracellular
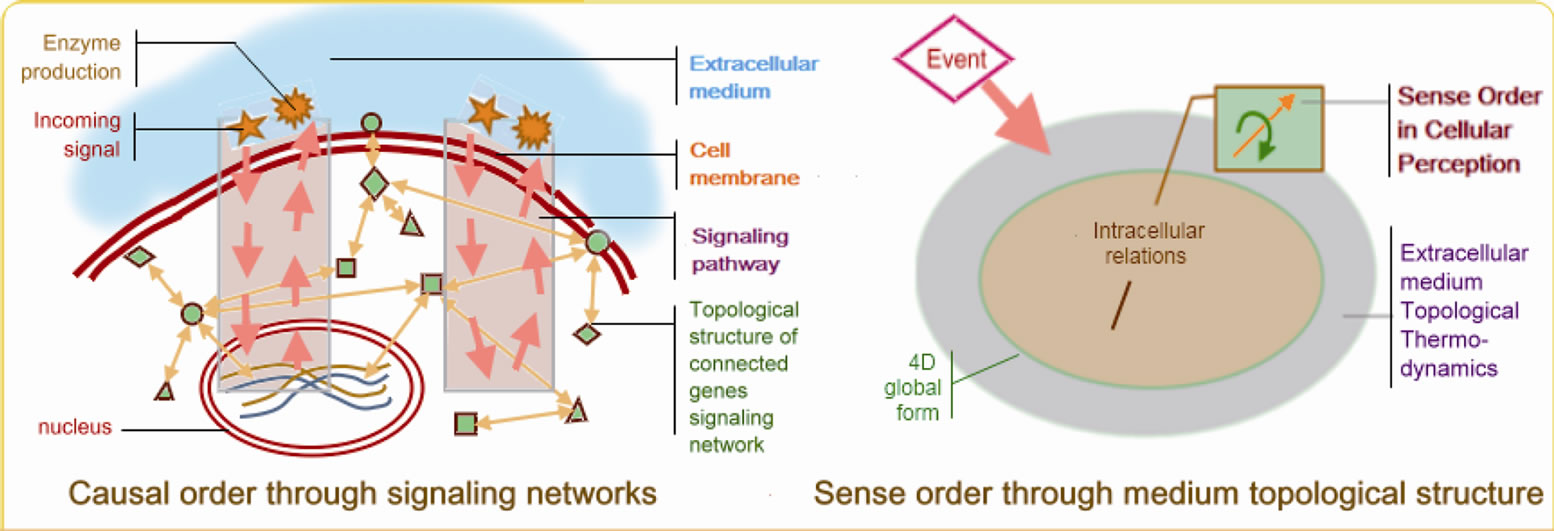
Figure 2. N˚2 Causal order and sense order in cellular perception: shows the two sides of cellular perception, which are causal order and sense order. For causal order, a principle of mechanical genes connectivity is illustrated on the left part of the figure. For example, in the calcium signaling pathway, which sometimes functions independently of the genetic nucleus, the influx and efflux modulation of calcium ions, Ca2+, are qualitatively consistent with known transcriptional changes of ARN messenger, ARNm, as indicated by Wang, (2009, [34]). Many genes expressions may be topologically connected to proteins of the signaling networks and also connected to pathophysiological status. But Molecular Reductionism has no explanation about the overall “mechanism” allowing these cointegrated transcriptional changes. It could be a super-mechanism, based on sense order. The right part of the figure shows that after some events the variations of the extracellular medium topological thermodynamics can modulate sense order, thus modulating intracellular relations. This sense order modulation by a qualitative topology is a holist idea in that it is always associated with the idea of any organism’s total form.
medium. The work of the biologist J von Uexküll, (quoted by Kull, [35]), had anticipated this concept.
EMF are external events which may affect extracellular medium structure, then sense order, then cellular behaviour, depending on the cell pathophysiological and cycle status. The EMF bioeffects are marked when cells are out of balance, such as the injured tissues cells, or cells of embryonic development tissues: When a cell is healthy balanced, its signaling calcium pathway moves temporarily under the influence of an EMF event, then restores, observed Wang [34]. But when a cell is out of balance, it appears to be more receptive with the applied organization models due to the electromagnetic modulation of its medium.
4. The Biological Interaction of Static Magnetic Fields, SMF
4.1. The Primary Target
Reductionism offers a conventional explanation of EMF bioeffects in terms of a current density acting on some molecules movements. EMF bioeffects cannot thus be explained as soon as the induced current density is too low to disturb the atomism of nervous tissues, as Liboff, (2004, [5]) observed. A non-Reductionist model could thus be more realist.
According to the conventional model, the target for the SMF biological interactions is the outer cellular membrane. Ayrapetyan, (2006, [36]), suggested a more general primary target which is the aqueous medium which bathes the membrane and acts on the membrane components. Further forwards, the primary target could be the structure of the interaction field situated in the physical vacuum between the extracellular medium molecules. In the liquid water, between two H2O molecules attracted each other by the power of an electrostatic hydrogen bond, type HHO|||||HOH, the void distance is at least 2,4 Ä. The extracellular medium space thus mainly consists of an Aqueous Physical Vacuum, APV, which is the space of the electromagnetic fields and thermal interactions.
4.2. Topological Approach of the Extracellular Aqueous Medium
A topological model of thermodynamics for non-equilibrium systems was described by Kiehn, (2008, [37]). The living matter answers this model because its thermodynamics is out of balance and its irreversibility implies a topological evolution. The topological model is based on the differential topological expression that the mathematician Cartan gave for the first law of thermodynamics*10. It is assumed that the vector fields whose fluctuations can be connected to certain world events, (thus for the fluctuations of the electric and magnetic energy densities,) can modulate the topology of the aqueous extracellular medium. The reason is that the overall differential topology includes in fact several topologies, (which are presented in appendix.) The first topology applies to the aqueous extracellular medium molecules, the second relates to the physical vacuum where the interaction fields are, and the third applies to bosons, (particles such as are the photons.)
A result of this topological approach is that EMF, SMF, interaction corresponds to a cubic curvature not concerned by forces produced by the E and B fields, but potentials A and of Φ, as well as charge current densities. This Kiehn result crosses perfectly with the quantum approach of Binhi, (2007, [38]). According to Binhi, the state of a molecular target interacting with a Pulsed Magnetic Field, PMF, of a very low frequency does not have to be described by the forces E and B, but by the potentials A and Φ, as in the Aharonov-Bohm effect11. Andocs, (2009, [39]), used this idea to show that when two charged particles create an electrochemical connection, their interaction could be changed by the application of a magnetic vector potential A. The effect would apply to bifurcation of hydrogen bonds in water, while bringing certain changes in the thermodynamic topology of the protons subset. The SMF effect on the water protons subsets which was observed by Lazarekis, (2009, [4]), by means of the infra-red spectroscopy, can also depend on the induction force B of MF.
4.3. The Action of the Magnetic Field on the Extracellular Aqueous Medium
Water is organized in continuous network by means of the hydrogen connections, H-bonds, of which the lifespan is about the picosecond. The co-operative effect of the meshs of the network is impeded by thermal agitation, indicates Mentré, (1995, [41]). In addition there are the impeding effects of the H-bonds defects which are due to the relative acidity of the water. These defects have one more or less H+ proton, (according to whether the hydrated ions are of type H3O+, acidity, or OH–, alkalinity). The H-bond defects by lack of one proton H+ were described by Petrenko, (2004, [42]). They are showed on Figure 3.
At the same time, MF controls the spin states in the subsets of the extracellular medium, by supporting the existence of local configurations there. These fields are primarily those of diamagnetic water, majority variety of known as “para”, and paramagnetic water, minority variety known as “ortho”, according to the work of Tikhonov, 2002, [43]. After an exposure to EMF or MF, these sub-

Figure 3. Rotation of an alkaline, OH–, defect in a water structure: shows that in this case there would be a connection into negative between two O; it forms a pivot for the rotation and the displacement of the bridge failing this OH–. the defects by excess of H+, (i.e. bonds with H3O+), also move while swivelling. The effect of EMF, MF, exposure upon connection defects of water structure, was modelled by (Binhi, 2007, [38]).
sets configurations are preserved beyond the spins relaxation times which are of about one second, as indicates Binhi [38]. A possible dialectic appears on Figure 4.
4.4. Gas|Water or Solid|Water Interface and Magnetic Memory of Water
Even though relaxation phenomena in water last from picosecond to second, effects of EMF or SMF treated water upon matter, alive or inanimate, remain efficient during hours after water treatment, indicated Colic & Morse, (1999, [44]). Among long-term EMF, SMF, effects existing in the water structure and which alter its behaviour there are those which relate to the hydration of ions near surfaces and to the reactivity of interfacial water. The fact that outgassed water cannot be magnetized, implies that gas|water interfaces are necessary for EMF,
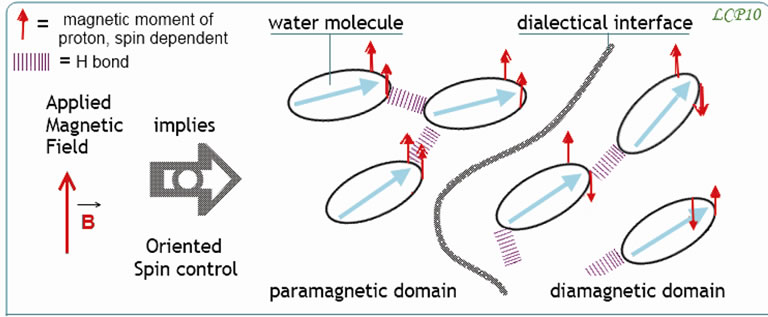
Figure 4. Effect of an applied magnetic field on a possible magnetic dialectic in water: shows that a “dialectical interface” can correspond to the “para” (diamagnetic) and “ortho” (paramagnetic) configuration of water. The fluctuation of this interface could be an event with some repercussion in the sense order of a cellular organism. This could not justify the very much debated theory of water memory; but it returns to the physical nature of the water magnetic memory.
MF, treatment of water, Colic indicated. The fact that EMF, MF, scale removing power is inhibited when pipe is in pure PVC indicates that apolar surfaces of pure PVC have something which is missing, Gabrielli, (2001, [45]) observed. What in both cases is necessary for the EMF, MF action, is the double ionic layer which is constitutive of the gas|water or solid|water interface. This ionic interface is very sensitive to EMF, MF, fluctuations, including natural MF fluctuations, and its changes relax very slowly, as Higashitani, (1995, [46]), Lipus, (2001, [47]), Vallée, (2004, [48]), observed. The double ionic layer interface ensures stability and evolution of the bubbles in water. The fact that bubbles evolution could be stabilized during hours after EMF, MF, exposure constitutes a kind of aqueous magnetic memory. But water organization around non-polar, or hydrophobic, surfaces is the reason of the sensitivity of the gas|liquid interface to MF, EMF, observed Colic.
Most part of time, water has microscopic bubbles of gas and it becomes partially degassed, then structured, by EMF, MF, exposure. This MF degasification affects protons subsets, thus imposes a progressive modification of the water organization and reactivity, when conversely the mechanically outgassed water gets no structured organization and cannot acquire some by MF. This way EMF, MF, treated water acquires a durable and reactive organization whose effects are observable in physics, chemistry and biology:
• In physics of habitat, water rise in wet walls constitutes a problem. The application of the Jurin law indicates that the surface tension12 allows a water rise lower than a height 30 cm in a capillary mass. However moisture coming from the basement easily goes up much higher. It does it because of the rise power of the hydrated positive ions (+) which are present at the solid| water interface. According to Le Chapellier, (2004, [49]), the basement moisture treatment by electrosmosis or by EMF (microwaves) allows to carry out a broad walls draining. The explanation could be that EMF treatment with microwaves creates a kind of treated water, i.e. a reduction of gas bubbles associated to rate of ions (+).
• In chemistry, structural effects of EMF, MF, treated water turned on to be equivalent to those of the direct exposure to same EMF, MF. That is true for the electrodeposition of nickel, indicated Devos, (1998, [50]), for the doping of nanoparticules, indicated Gavrilov-Yusim, (2008, [51]), or for the descaling of calcite to aragonite, indicated Higashitani, (1995, [46]).
• In biology the same equivalence between direct EMF exposure effects and EMF treated water effects can be observed, for example in case of activation of certain membrane channels, indicated Fesenko, (1995, [52]).
4.5. Membrane Surface Forces and Cellular Activity
Surface forces acting in water are both long and short range. As Craig, (1993, [53]) indicated, they include: 1) Short range electrostatics Van der Waals forces, 2) Long range attraction between non-polar, or hydrophobic, surfaces, 3) Oscillatory forces due to liquid structure and leading to aqueous clusters, 4) Forces dues to fluctuations in correlated surface dipoles, 5) Short-range hydration forces between lipids, 6) Ion correlation effect beyond the double layer that give rise to attraction forces, 7) Double layer repulsive forces, 8) Fluctuation forces between bilayers surfaces, 9) Effects of hydration ions size, etc.
As these surface forces are related to surface curvatures, they relate to tensors or also to the differential topology of the cell external medium. In that way surfaces forces can have an impact upon cellular sense order.
The lipid bilayer of a cell membrane has opposite forces involving the polar head group and the hydrophobic hydrocarbons tails. Long range curvature forces compete with hydrophobic matching forces in the membrane, so that the lipids geometry on the proteins conformational energetic and organization. Botelho theory, (2006, [54]), indicated that spontaneous curvature of the lipid bilayer tends to match with the mean curvature of the lipid|water interface adjacent to the protein. The biological activity may be thus regulated by the water structure, because of the membrane lipids whose spontaneous curvature most closely matches the active state of the proteolipid assembly. According to Rosen, (2010, [55]), these proteins deformation effects due to the phospholipids surfaces curvatures could be corrected by SMF exposure, because of a direct SMF action towards the diamagnetic anisotropy of phospholipids. But why applied SMF would be perfectly oriented in each membrane point? It is more logical to admit a more general action of the extracellular magnetic weak and indirect surface forces, giving to the extracellular medium its bathing reactivity. This bathing reactivity can thus produce the effects observed on the intramembrane proteins after exposure to EMF, SMF, as it can be seen on Figure 5.
This mechanical presentation, adapted from the Bothelho theory, is perfectly Cartesian. It explains the clustering of the intramembrane protein distribution after PMF exposure, observed by Marinelli, (1997, [56]) and the channel opening through an indirect action of geo
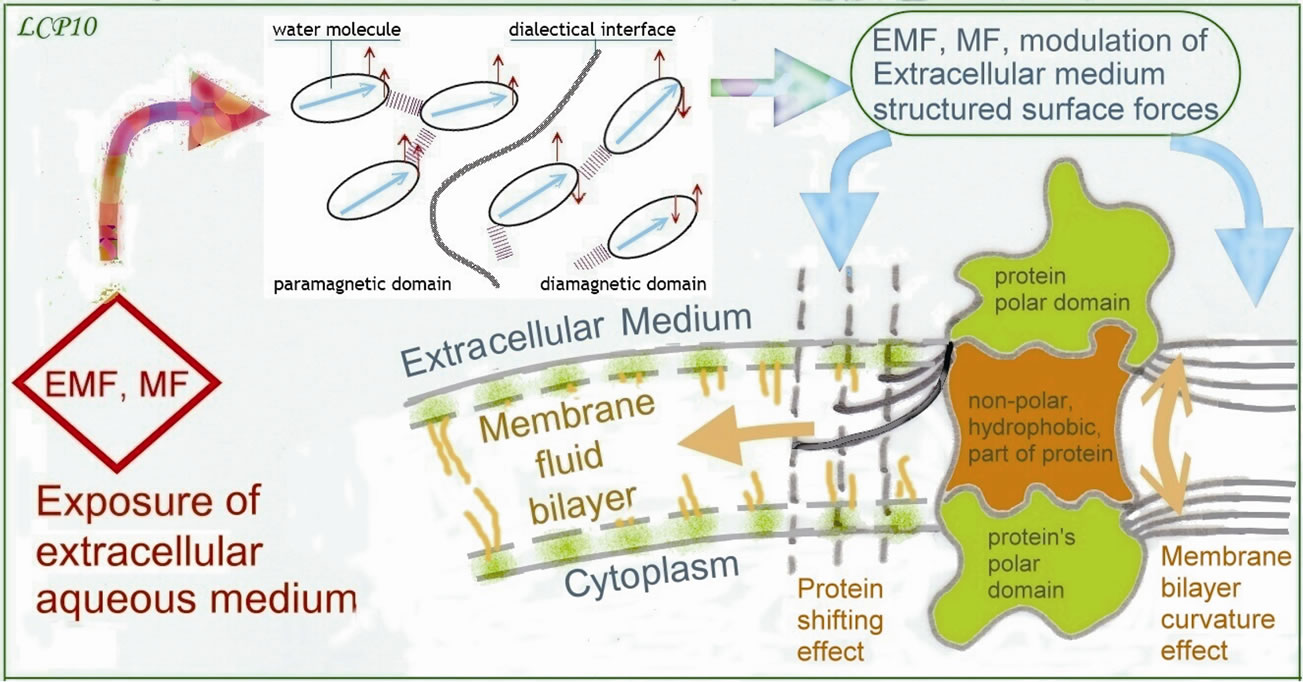
Figure 5. Curvature mismatch of liqid bilayer thickness to the protein receptor due to an EMF, MF treatment of extracellular medium, (from after Botelho theory): shows a possible action of an EMF, MF, treatment of extracellular aqueous medium upon a membrane protein, through a modulation of the structured surface forces. These geometrical forces could be acting on the one hand by some curvature effect, implying a protein reversible denaturation directed towards activation or inhibition, and on the other hand by a shifting effect depending on water domains redundancy. The membrane protein may be inhibited, or activated, so as displaced in the bilayer to form clusters or rafts. These clusters of protein-receptors formed in membrane have an important mechanical role in cellular signaling.
metrical forces upon membrane, after an extracellular medium exposure to EMF microwaves, observed by Fesenko [52]. But it doesn’t explain the observed biphasic effect. This one depends more on the non-Cartesian sense order of the cell, as it proceeds on the medium qualitative topology.
4.6. The Effect of the Magnetic Sensitivity of the Stern and Gouy-Chapman Double Ionic Layer
Modulation of sense order and reorientation of cellular activity after a magnetic exposure of the extracellular medium seem to depend on the CO2 bubbles which are present in biological water. The double ionic layer which interfaces gas|liquid ensures the stability and evolution of the bubbles in water. The fact that bubbles evolution could be stabilized during hours after EMF, MF, exposure constitutes a kind of aqueous magnetic memory.
The double ionic layer implies a Zeta potential, as it can be seen on Figure 6. The exposure of non-degassed water to EMF, MF, brings about a ξ potential variation, mainly decreasing. The reduction in the ξ potential implies ions recombinations in the double layer, Higashitani [46], stated. It results an outgassing of CO2 molecules, Vallée [48], indicated. This corresponds to a progressive reduction of bubbles size and number of hydrated protons, H3O+. That means a medium alkalisation, which had been observed by Higashitani.
But what could be the initial mechanism which causes the CO2 recombination during EMF exposure? Hisgashitani proposed a magnetohydrodynamic explanation which is valuable for moving water only. For calm water, the explanation can be an evolution of aqueous medium thermodynamic topology after a magnetic field exposure:
According to Binhi [38], then Le Chapellier [57], the aqueous medium is structured by hydrated protons H3O+ subsets, while MF, through spin control, affects the H3O+ defects movements. The modulation of the protons hydration and translocation affects the aqueous domains, (Bankura, 2005, [58]), and modulates the activity of hydrated proteins, (Wikström, 2003, [59], Mentré, 2005, [60]) while presenting a topological interaction. It also leads to the recombination of hydrated protons H3O+ with bicarbonate ions HCO3–. This way the water treated by EMF, MF, gradually acquires a state of higher organization durable some hours.
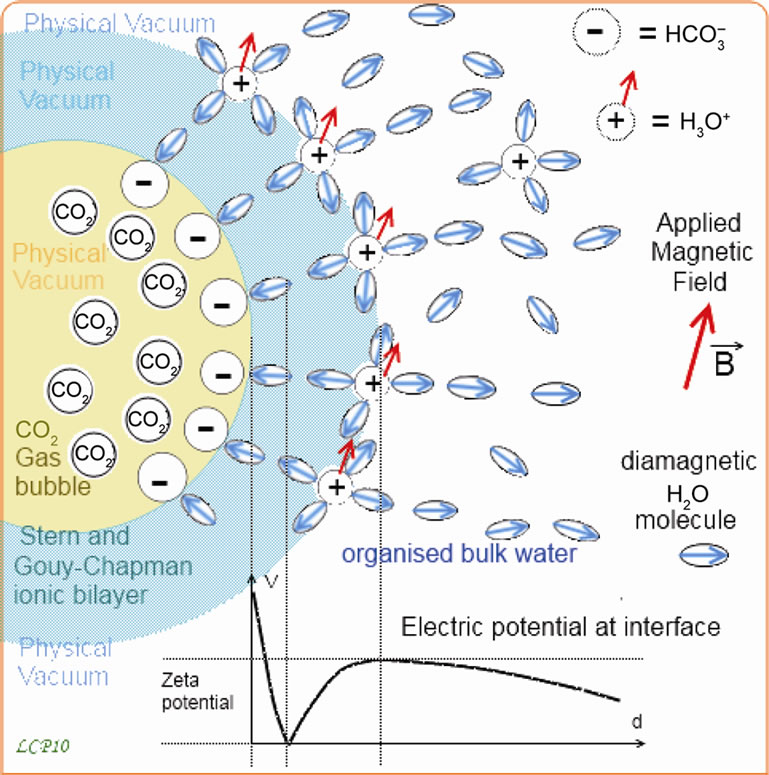
Figure 6. CO2 gas bubble in an aqueous medium: shows the structuring effect of a carbon dioxide bubble in water. This bubble behaves like a colloid, in the sense that its interface includes a double ionic layer whose make up consists of water and CO2 in a dissolved state. The hydrolysis reaction of CO2 is: CO2 + 2O2 ↠ HCO3– + H3O+. The Stern layer, situated on the internal side, is made of hydrated HCO3– ions. The Gouy-Chapman layer, situated on the aqueous medium side, is made of hydrated H3O+ ions. The electric tension between the two layers, called Zeta potential, can reach about –65 mV, as Vallée [48], indicated. The figure shows that the ξ potential is extended in the surrounding water and reduced with the distance. That means the double layer propagates an organization of the water molecules around it, as it can be seen on figure. This water organization which faces thermal agitation and H-bonds connection defects, is translated in the background surface forces fields structure, i.e. the topological structure of the Aqueous Physical Vacuum.
In topological terms, a state of higher organization corresponds, in the background physical vacuum, to a topological structure of bosons13. But in philosophy of Nature terms, the perception that a cell can have of a bosons structure cannot be described, since the sense order is irreducible. Reversely, if one cannot know what is really perceived by an individual sensitivity, epistemological reduction ensures that the sense order is overall modulated by the cell being-envelope, i.e. by the qualitative topology of the extracellular medium.
In the Fesenko example [52], the patch-clamp technique, (which is an electric technique of insulation of a cellular membrane fragment,) enables him to measure the currents through a Ca2+ dependent K+ membrane channel. The exposure of in vitro cultured cell to EMF, µ-waves, 46 GHz, produces a biphasic effect on the observed channel, opening during a few minutes, and then closing. The change of the aqueous medium of a cell not-treated by EMF by a medium similarly treated by EMF, 46 GHz, produces the same biphasic effect on the channel. The bioeffect occurs in both cases, in spite of the electric disturbance locally imposed by the patch-clamp micropipettes. The common cause is the EMF extracellular medium treatment which modulates topological structure. It is an event which affects not only proteins of the channel but also the cellular perception.
4.7. Medium Conditions and Possible EMF Bioeffects
More than simply temporal, the biphasic effect can result in a distinct behavioural answer, as if the cell chose from after its perception. A higher structure of medium surfaces forces is not necessarily a significant fact for a cell: Healthy living tissues in equilibrium are overall transparent with the magnetic field which causes this organization.
It is thus necessary to associate the bosons topological structure with a non-mechanical “sensitive touch” (“un touché sensible”), which gives the cell a sense of its behavior modulation. What can be the physical ingredient? The literature, quoted by De Mattei, (2007, [61]), indicates that the biological effects of the EMF depend on the EMF physical parameters, and in particular depend on the waveform. In addition a significant role of some chemical actors has been shown for the chondrocytes response to EMF.
The foetal calf serum, FCS, role has been shown by Pezetti, (1999, [62]): The presence in the extracellular medium of this serum helps proliferation of cultured human chondrocytes. It stimulates the synthesis of proteoglycanes, PG, by the articular cartilage cells, indicates De Mattei, (2004, [63]). When 10% of fresh FCS are added in the water of the bath, stimulation by EMF causes a proliferation of the chondrocytes cultivated in low density. In the cultures with high density, the effect occurs only temporarily. Admitted that the cell answer to EMF exposure depends on its sense order, then its medium structure perception, it thus depends on several microenvironment factors, as De Mattei, (2001, [64,65]) indicated.
The chondrocytes express collagen II ARNm, which is a specific marker of the chondrocyte phenotype, Pezetti [62], indicated. When chondrocytes are cultured into monolayers, they show phenotype instability; so they can dedifferentiate into fibroblasts, which results in a more flat conformational change and by collagen I synthesis instead of II. The EMF treatment removes this instability and maintains round morphology, De Mattei [63], indicated.
On the other hand, Jahns, (2007, [66]), showed that chondrocytes stripped of their extracellular matrix and cultured in monolayers, take a spindle morphology extremely studded at the bottom. After EMF exposure morphological variations occur, passing from spindle to star, then to the round sphere which allows chondrocytes to leave bottom: The cell behavior evolution after EMF exposure translated into a reduction of 30% of the bottom anchors.
4.8. The Evident Role of Calcium Signaling
The cell behavior evolution can be explained by a coherence contribution in sense order as it is embodied in the calcium signaling events. For example, Cadossi, (1992, [67]), indicated the Ca2+ influx is an early event of the lymphocyte activation process, concomitant to the clustering of some membrane receptors. So the stimulation of lymphocytes with mitogen is a prerequisite for an evident increase of Ca2+ influx after EMF stimulation. In HL-60 cells, Zhou, (2002, [68]), showed that gene transcription changes induced by EMF stimulation are dependent both extracellular and intracellular Ca2+. In chondrocytes, the synergy of EMF stimulation and presence of FCS can be also explained by the calcium signaling. Mac Neil, (1984, [69]), showed the existence in FCS of CaM-like enzymes, with a different molecular weight but a similar activity to the one of calmoduline, CaM. Thereafter, WenQiang, (1997, [70]), showed that four major proteins which bind to CaM activated by Ca2+ were detected in the chicken serum and that growth promoters can also bind to CaM. The WenQiang suggestion, not checked, is that growth promoters as TGF-β1 could be stimulated in the extracellular medium by an activated CaM-like, before being able to effectively address to the specific membrane receptors of chondrocytes. The calcium signaling events associated with the complex Ca2+/ CaM are notably responsible for the changes controlling the actine filaments network within the chondrocytes cytoplasm, (Onuma 1988, [71]), what directs the cell morphological changes, (Jahns, 2007, [66]).
All this suggests that the extracellular medium action on the cellular perception can operate by coupling a nonmechanical qualitative effect, according to sense order, with a mechanical quantitative effect, according to the causal order implying calcium signaling: The membrane receptor proteins denaturization, activation or clustering due to medium structure curvatures, is thus concomitant with a calcium flux modulation depending on cell status.
5. The Double Bioeffect Resulting from Electromagnetic Fields Exposure
The exposure of cells and their medium to Electromagnetic Fields, EMF, may results in a double observable effect. The former relates to the calcium signaling variations, the latter relates to the gene expressions.
5.1. The First Effect
It is well known that the intracellular calcium level, (Ca2+)i, presents a natural fluctuation which constitutes a principle of regulation, (Cho, 2007, [72]). The fluctuation rhythm of (Ca2+)i takes into account of the other active ions, (Shuster, 2001, [73]). Rhythm acts in accordance with the Na+/K+ pump activity, the Na+/Ca+ exchanger, and with the voltage operated channels calcium, VOC, activity. Each concentration peak in calcium ions mechanically supports the graduated activation of sensors such as CaM.
The observations carried out since 1978 showed that exposure to EMF, MF, causes a biphasic variation of calcium, due to a variation of influx or efflux of Ca2+ ions. Its direction depends on experimentation condition, type and state of cells. A similar concentration of the various elements operating the calcium cycle may modify in a different way the response to a stimulus, according to cell situation and type, because of a different modulation of calcium signalsome. The variation of the calcium flow according to EMF, MF exposure, is thus imperfectly reproducible in-vitro, undoubtedly because of the non-mechanical part in cellular perception. The significance for the cell of the structuring of medium surfaces forces after an EMF, MF, exposure cannot be completely explained.
However there is a pain relief after exposure to MF which is frequent and which can be explained through calcium signaling. On the one hand, algogenic substances such as bradykinine modulate up the (Ca2+)i level, (Bathon, 1994, [74], Gelperin, 1994, [75]). In addition, according to Voitenko, (2006, [76]) fluctuation disorders and thus homeostasis of intracellular calcium, when observed in dorsal root ganglion, DRG, neurons as in dorsal horn DH, neurons, form the common denominator of the various types of pains. The contribution of the cell sense order after a MF exposure could be a corrective step of the calcium signaling disorders. Remember it is an indirect MF effect, because Ca2+ ion has no magnetic moment. The modes of MF actions upon Ca2+ ion and calcium signaling are suggested on Figure 7.
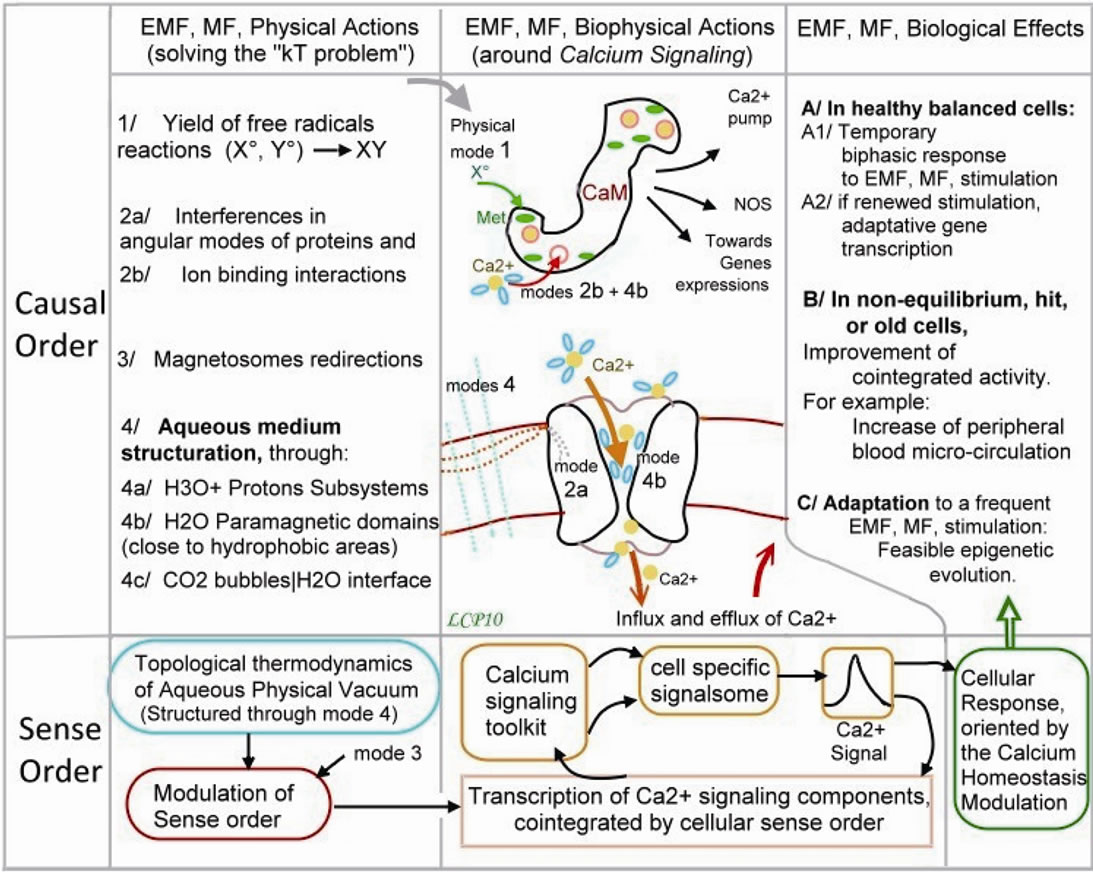
Figure 7. Calcium signalling and cellular bioeffects after EMF, MF, exposure: shows the table by which a magnetic field exposure can act on living cells. Horizontally are the physical then biophysical actions leading to biological effects. Vertically are the causal order relating to mechanisms, situated over the sense order relating to qualitative data. Some physical action modes may solve the kT problem, (relating to thermal agitation forces): 1/ Free radicals, 2a/ Protein angular interferences, 2b/ Ion binding interactions, 3/ Magnetosomes, 4/ Aqueous medium structure, [38]. They can act upon calmoduline status, through free radicals acting on Methionine oxidation and through ion binding interaction. They can act upon channel opening through aqueous structure, and upon Ca2+ pump through calmoduline status, [83]. It is stated that the qualitative topology of the aqueous medium structure modulates the cell sense order. This one is supposed to ensure a cointegrated transcription of the calcium signaling components. It leads to a calcium homeostasis which is crucial for observable biological effects.
5.2. The Second Effect
The second effect of EMF, MF, exposure relates to the modulation of genes connected with the signaling pathways components. Hanada, (2007, [77]) showed that in neuronal cells (of wild worms C Elegans,) genes connected to calcium signaling pathway or to synaptic transmission have expression levels modified after exposure to MF, 5000 Gauss14. Exposure to MF of cells Escherichia coli induces effect on gene expressions and cell multiplication, indicated Potenza, (2005, [78]). The possible migration of cells can be reinforced by exposure to MF, depending on the kinds of cells, which implies a transmission of significant information, indicated Hashimoto, (2007, [79]). Wang, (2009, [34]), showed that the exposure of embryonic cells to SMF modulates genes expressions of more than nine signaling networks associated to the inflammatory cytokine IL-6. Wang adds that magnetic fields of about 2500 Gauss do not have any observable effect in differentiated cells of healthy tissues. Response of cells to EMF, MF, thus depends on their nature, status and situation. The biphasic biological response of cells treated by MF falls over according to the cell conditions, indicated Xu [80], Okano [81], Mayrowitz [82]. Such a selective and directed biological effect EMF, MF, exposure is explained by the naturalist approach of cellular perception, but it is not explained not by the conventional and reductionist cell model.
6. Conclusions: The Naturalism of Cellular Perception
6.1. The Objective Horizon of the Cellular Perception Theory
Cellular perception theory combines intracellular signaling mechanics with extracellular qualitative topology. This combination allows to overstep the Cartesian aporias relative to unjustified cybernetic regulation and to sense order set out of scientific biology.
A justified cybernetic regulation feed-back is shown on Figure 7, with the diagram relating to the calcium signaling regulation, where sense order constitutes a kind of super-mechanism.
A suffering patient always presents a fluctuation defects of the calcium signal in some of its neurons, indicated Voitenko [69]. These fluctuation defects do not have a logical justification in only terms of molecules analysis. The Holistic approach requires a totality to justify the regulation diagram of the calcium signaling. This necessary totality is the one of the sense order where some events of the extracellular micro-environment may resound in structural terms. That is why the biophysical action of a magnetic exposure can offer a pain relief15. The biophysical effect of MF upon cellular calcium which has no magnetic moment must be clearly explained. But MF actions on biological magnetic species, (as magnetosomes,) and paramagnetic species, (as free radicals, paramagnetic water, protons,) are insufficient to explain the MF modulation of the calcium signaling homeostasis. The important point consists in translating into objective terms the possible coherence contribution concerning calcium homeostasis after MF exposure. The naturalism of cellular perception suggests that external qualitative topology terms modulate a sense order whose coherence degree sets a constraint on internal calcium homeostasis.
Cellular perception also includes subjective aspects which cannot be studied by naturalism with objective horizon. Thus for example the sense order in a cellular organism perception also includes probably a psychic side, according to an elementary material unconscious. But its study, at the interface between biology and psychology, is out of objective naturalism.
6.2. Temporal Thickness and Cellular Resistance to EMF Exposure
Detailed descriptions by patients showed the existence of some “cellular resistance” which rises after some sessions to oppose the dictation of the applied PEMF treatment. It also appears during animals’ treatment, as De Mattei (2007, [61]) observed. To stop the genetically irreversible evolution of the articular cartilage in the old Dunkin-Hartley pigs, the chondrocytes stimulation by PEMF must be important in duration, and to be lengthily and patiently repeated, because a cellular resistance opposes the dictating of the treatment:
Insofar as the most elementary psychic material is the feeling, cells have probably a psychic part which is an elementary material unconscious. This way, the evolutionary change under dictation of a repeated event, is not done without a natural attempt to get back to the previous order, because material unconscious is former to the present cell status with its molecules organization, [84, 85]. In that way, topological nuances which integrated the being-envelope during PEMF treatment meet consist of a temporal thickness. Within this temporal thickness the constant modulation of the signalsome ensures all at the same time both the cell endurance and its possible adaptation to the changing world. Temporal thickness is what allows the organism to be enduring, rising all at the same time both a resistance to events dictation and an adaptation faculty.
7. Appendix: Cell Topological Thermodynamics as Connected to Electromagnetic Theory
Topology is the study of properties that remain invariant under deformations through homeomorphisms, (i.e. oneto-one correspondence of the set’s points.) With Gauss, then Riemann, differential geometry applied to metric non-euclidian surfaces leading to the idea of topological spaces [86]. Now, applied topologies are said to be connected if they are all in one piece. With contrast, a qualitative topology may be defined as neither metric, nor connected.
In biological systems, qualitative development involves an alteration in the structural and functional nature of system, as it can be by entropy effect, or by an unusual expression of genes [87]. A qualitative topology of biological systems should thus take account for entropy.
As a Kolmogorov Topology may be non-metric, disconnected and additive for entropy, it could be a candidate to describe a qualitative topology. By example, the topological thermodynamics structure of any system, as described by Kiehn, (2009, [88,89]), is a KolmogorovCartan T0 topology, given the symbol, KCT0. This topology, with sets that are exterior differential forms, is disconnected into two components representing equilibrium systems, (Pfaff dimension <3) and non-equilibrium systems, (Pfaff dimension >2). Living systems are nonequilibrium systems.
In this perspective, a differential 1-form of Action A on a differential variety {x, y, z, t, dx, dy, dz, dt} may encode topological electromagnetism of the thermodynamic living system. The coefficient functions of the 1-form of Action A, per unit charge, are the symbols representing the vector, A, and scalar, Φ, potentials of electromagnetic theory. The exterior differential of A is utilized to produce the 2-form, F = dA, of field intensities B and E, which are the limit sets of the Action relative to the KCT0 structure. Only the rotational components of A act as a magnetic field, with B = curl A. An interaction energy density and the total energy density are defined as:
The interaction energy density = (A·J – ρΦ). (1)
The total energy density = (B·H – D·E) – (A·J – Φ). (2)
In these two formulas, the classic symbol definitions are: Magnetic potential vector A; Electric potential scalar Φ; Excitation vectors: magnetic H = µB; electric D = eE; Current density: J = curl H – ∂D/∂t; Charge density: ρ = div D; Magnetic permeability µ; Electric permittivity e.
Most of the particles of the finite cell structures are distinguishable, corresponding to a discrete Hausdorff T2 topology which can be partitioned into a qualitative T0 component. With T × 0 as topological dual, (compliment) of T0, Kiehn, (2010, [90,91]), demonstrates that the power set topology, T2, can be partitioned as:
T2 = T0 union T × 0 union R;
(or: T2 = T0 È T × 0 È R), (3)
where R is a Not-T0 residue that can have indistinguishable singletons (possibly representing bosons.) It is conjectured that the living cell medium (represented by a T2 discrete topology) can be altered by the modulation of a KCT0 electromagnetic component of a T2 partition, thereby inducing an evolutionary thermodynamic change in the living cell medium, T2.
The evolutionary process may be accompanied by a Not T0 residue component, R, representing indistinguishable quantum collective states of bosons. This way, after a magnetic field exposure, any modulation of the medium qualitative topology T0 may affect the topology T2 of the cell molecules make up and possibly affect the associated topology R about quantum collective states. Now Binhi, (2007, [38]), indicated bosons collective quantum states are the key to get top bioeffects after a magnetic field, MF, exposure.
REFERENCES
- J. F. Schenck. C. L. Dumoulin and P. Souza, “Health and Physiological Effects of Human Exposure to Whole-Body 4 Tesla Magnetic Fields during Magnetic Resonance Scanning, Book of Abstracts,” Society of Resonance Imaging in Medicine, Berkeley, 1990, p. 277.
- L. Teodoni, W. Göhde, Mg. Valente, F. Tagliaferri, et al., “Static Magnetic Fields Affect Calcium Fluxes and Inhibit Stress-Induced Apoptosis in Human Glioblastoma Cells,” Cytometry Part A, Vol. 49, No. 4, 2002, pp. 143- 149.
- P. Nagy, “The Effect of Low Inductivity Static Magnetic Field on Some Plant Pathogen Fungi,” Journal of Central European Agriculture, Vol. 6, No. 2, 2005, pp. 167-171.
- P. Lazarekis, “Effects of a Static Magnetic Field on Biological Samples,” Thesis, University of Wollongong, Australia, 2009. http://trove.nla.gov.au/work/359668578?selec-tedvesion=NBD44884397
- A. R. Liboff, “Towards an Electromagnetic Paradigm for Biology and Medicine,” The Journal of Complementary and Alternative Medicine, Vol. 10, No. 1, 2004, pp. 41- 47.
- R. E. Jacobs, “MRI: Volumetric Imaging for Vital Imaging and Atlas Construction,” Imaging in Cell Biology, September 2003, pp. SS10-SS16. http://www.nature.com/focus/cell-bioimaging
- R. Thom, “Modèles Mathématiques de la Morphogenèse,” Christian Bourgeois, Paris, 1980, noté MMM.
- H. Poincaré, “Analysis Situs,” Journal de l’Ecole Polytechnique, 2e ser., Paris, 1895, Cahier I.
- E. Chautard, N. Thierry-Mieg and S. Ricard-Blum, “Interactions Networks: From Proteins Functions to Drug Discovery. A Review,” Pathologie Biologie, Vol. 57, No. 4, 2009, pp. 324-333. doi:10.1016/J.patbio.2008.10.004
- A. Toscano, “Merleau-Ponty, Whitehead and the Politics of Nature, Merleau-Ponty and the Philosophy of Nature Conference,” University of Warwick, Warwick, May 2003.
- R. Thom, “La Théorie du Cobordisme, Fields Medal Conference,” Edimbourg, 1958.
- M. Merleau-Ponty, “La Phénoménologie de la Perception, Gallimard,” Paris, 1945.
- J. Largeault, “Réductionnisme et Holisme,” Encyclopedia Universalis, Paris, 2006.
- J. R. Munson and R. C. York, “Nature, Philosophy of, Encyclopaedia Britannica,” Chicago, 2007.
- C. Gillain, “La Théorie Qualitative de Poincaré et le Problème de l'intégration des Équations Différentielles,” In: H. Gispert, Ed., La France Mathématique: La Société Mathématique de France, 1872-1914, SFHST-SMF, Paris, 1991, pp. 215-242.
- J. Mawhin, “The Early Reception in France of the Work of Poincaré and Lyapunov in the Qualitative Theory of Differential Equations,” Philosophia Scientiae, Vol. 1, No. 4, 1996, pp. 119-133.
- A. N. Whitehead, “The Concept of Nature,” Cambridge University Press, Cambridge, 1920.
- A. N. Whitehead, “Science and the Modern World,” The MacMillan Company, New-York, 1925.
- A. N. Whitehead, “Symbolism, Its Meaning and Effects,” The MacMillan Company, New York, 1927.
- A. N. Whitehead, “Process and Reality,” The MacMillan Company, New-York, 1929.
- M. Merleau-Ponty, “La Nature-Notes,” Cours au Collège de France, Seuil, Paris, 1995.
- R. Barbaras, “Merleau-Ponty et la Nature,” Atelier Merleau-Ponty, U de Toulouse, 9 Septembre 2000.
- P. A. Miquel, “Qu’y-a-t-il de Vital dans un Organisme Vivant?” Bulletin d’Analyse Phénoménologique, Vol. 2, 2010, pp. 313-337.
- M. Blank and R. Goodman, “Electromagnetic Fields Stress Living Cells,” Pathophysiology, Vol. 16, No. 2-3, 2009, pp. 71-78. doi:10.1016/j.pathophys.2009.01.006
- M. J. Berridge, “Cell Signaling Biology,” Portland Press Limited, Portland, 2008. http://www.cellsignallingbiology.org
- A. Z. Fire and C. C. Mello, “RNA Interference, Advanced Information,” The Nobel Prize in Physiology or Medicine, 2006. http://nobelprize.org/nobel_prizes/medicine/laureates/2006/adv.html
- M. Radman, “Enzymes of Evolutionary Change,” Nature, Vol. 401, No. 6756, 1999, pp. 868-869.
- S. L. Berger, T. Kouzadires, R. Shiekhattar and A. Shilatifard, “An Operational Definition of Epigenetics,” Genes & Development, Vol. 23, No. 7, 2009, pp. 781-783. http://genesdev.cshlp.org/con-tent/23/7/781.full doi:10.1101/gad.1787609,
- M. W. Ho, “How Development Directs Evolution Epigenetics & Generative Dynamics, Evolution and the Future Conference,” Hotel Continental-Beograd, Belgrade, 14- 18 October 2009.
- D. Baulcombe, “Of Maize and Men, or Peas and People: Case Histories to Justify Plants and Other Model Systems,” Nature Medicine, Vol. 14, No. 10, 2008, pp. 20-28. doi:10.1038/nm1008-1046
- M. Mott, “Did Animals Sense Tsunami Was Coming?” National Geographic News, January 4, 2005
- M. Heidegger, “Lettre sur l’humanisme, tr. R. Munier in Questions III,” Collection Tel, Gallimard, Paris, 1959 (in French, from German).
- A. Tan, A. Jimeno, B. Rubio-Viqueira and M. Hidalgo, “NCI-60 Gene Set Connectivity Map, GS-Cmap: Connecting Pathway-Based Gene Expression Profiles for Therapeutic Efficacy Determination,” Journal of clinical oncology, ASCO Annual Meeting Proceedings, Vol. 26, 20 May 2008, p. 3588.
- Z. Wang, A. Sarje, P. L. Che and K. J. Yarema, “Moderate Strength (0.23-0.28 T) Static Magnetic Fields (SMF) Modulate Signaling and Differentiation in Human Embryonic Cells,” BMC Genomics, Vol. 10, No. 356, 2009, pp. 1-23. www.biomedcentral.com/1471-2164/10/356
- K. Kull, “Jakob von Uexküll: An Introduction,” Semiotica, Vol. 134, No. 1-4, 2001, pp. 1-59, [Includes complete bibliography of Uexküll].
- S. N. Ayrapetyan, “Cell Aqua Medium as a Primary Target for the Effect of Electromagnetic Fields,” Bioelectromagnetics Current Concepts, NATO Security through Science Services, Springer, 2006, pp. 31-63. doi:10.1007/1-4020-4276-8
- R. M. Kiehn, “Non-Equilibrium Systems and Irreversible Processes,” Topological Torsion and Macroscopic Spinors, Lulu, Vol. 5, 2008. http://www.lulu.com/kiehn
- V. N. Binhi and A. B. Rubin, “Magnetobiology: The kT Paradox and Possible Solutions,” Electromagnetic biology and Medicine, Vol. 26, No. 1, 2007, pp. 45-62.
- G. Andocs, G. Y. Vincze, O. Szasz, P. Szendro and A. Szasz, “Effects of Curl-Free Potentials on Water,” Electromagnetic Biology and Medicine, Vol. 28, No. 2, 2009 , pp. 166-181.
- A. van Oudennaarden, M. H. Devoret, Y. V. Nazarov and J. E. Mooij, “Magneto-Electric Aharonov-Bohm Effet in Metal Rings, Letters to Nature,” Nature, Vol. 391, 1998, pp. 768-770.
- P. Mentré, “L’eau dans la Cellule, Masson,” Paris, 1995, ISBN 2-225-84608-1 (in French).
- V. E. Petrenko, et al., “The Influence of Inclusion of Non-Electrostatic OH interaction in the Empirical Pair Potential for Water on the Mutual Orientation of Molecules in the Nearest Surround,” Saint-Petersbourg, Russia, 12-16 July 2004.
- V. I. Tikhonov and A. A. Volkov, “Separation of Water in Its Ortho and Para Isomers,” Science, Vol. 296, No. 5577, 2002, p. 2363
- M. Colic and D. Morse, “The Elusive Mechanism of the Magnetic Memory of Water,” Colloids and Surfaces A, Vol. 154, No. 1-2, 1999, pp. 167-174.
- C. Gabrielli, R. Jaouhari, G. Maurin and M. Keddam, “Magnetic Water Treatment for Scale Prevention,” Water Research, Vol. 35, No. 13, 2001, pp. 3249-3259. doi:10.1016/S0043-1354(01)00010-0
- K. Higashitani, H. Iseri, K. Okuhara, A. Kage and S. Hatade, “Magnetic Effects on Zeta Potential and Diffusivity of Nonmagnetic Colloidal Particles,” Journal of Colloid and Interface Science, Vol. 172, No. 2, 1995, pp. 383-388. doi:10.1006/jcis.1995.1268
- L. C. Lipus, J. Krope and L. Crepinsek, “Dispersion Destabilization in Magnetic Water Treatment,” Journal of Colloid and Interface Science, Vol. 236, No. 1, 2001, pp. 60-66. doi:10.1006/jcis.2000.7392
- P. Vallée, “Etude de l’effet de Champs Électromagnétiques Basse Fréquence Sur les Proprieties PhysicoChimi-ques de L’Eau,” Ph.D. Thesis, Thèse de Doctorat de l’Université Pierre et Marie Curie, Paris VI, 2004.
- P. Le Chapellier, “L’Humidité des Lieux et la Santé,” Journées Toulousaines de l’Eau 2004, Toulouse, 19-20 Septembre 2004.
- O. Devos, A. Oliver, J. P. Chopart, O. Aaboubi and G. Maurin, “Magnetic Field Effects on Nickel Electrodeposition,” Journal of The Electrochemical Society, Vol. 145, No. 2, 1998, pp. 401-405. doi:10.1149/1.1838276
- N. Gavrilov-Yusim, E. Hahiashvili, M. Tashker, V. Yavelsky, O. Karnieli and L. Lobel, “Enhancement of Hybridoma Formation, Clonability and Cell Proliferation in a Nanoparticle-Doped Aqueous Environment,” BMC Biotechnology, Vol. 8, No. 3, 2008, p. 3. http://www.biomedcentral.com/1472-6750/8/3
- E. E. Fesenko, V. I. Geletyuk, V. N. Kazachenko and N. K. Chemeris, “Preliminary Microwave Irradiation of Water Solutions Changes Their Channel-Modifying Activity,” FEBS Letters, Vol. 386, No. 1, 1995, pp. 49-52. doi:10.160014-5793(95)
- V. S. J. Craig, B. W. Ninham and R. M. Pashley, “The Effect of Electrolytes on Bubble Coalescence in Water,” The Journal of Physical Chemistry (ACS Publications), Vol. 97, No. 39, 1993, pp. 10192-10197. doi:10.1021/j100141a047
- A. V. Botelho, T. Huber, T. P. Sakmar and M. F. Brown, “Curvature and Hydrophobic Forces Drive Oligomerization and Modulate Activity of Rhodopsin in Membranes,” Biophysical Journal, Vol. 91, No. 12, 2006, pp. 4464- 4477. doi:10.1529biophysj.106.082776
- A. D. Rosen, “Studies on the Effect of Static Magnetic Fields on Biological Systems,” PIERS Online, Vol. 6, No. 2, 2010, pp. 133-136.
- F. Marinelli, F. Bersani, S. Santi, M. Riccio, S. Petrini, A. Valmori and N. M. Maraldi, “50 Hz Pulsed Magnetic Fields Affect Intramembrane Proteins Distribution in Cultured Cells, Electricity and Magnetism in Biology and Medicine,” Proceedings of the Second World Congress, Bologna, 8-13 June 1997, pp. 31-32.
- P. Le Chapellier and B. Matta, “Cellular Perception and Static Magnetic Fields Active Penetration Depth for Pain Magnetotherapy,” PIERS Online, Vol. 6, No. 3, 2010, pp. 287-292. doi:10.2529/PIERS090930101455
- A. Bankura and A. Chandra, “Hydration and Translocation of an Excess Proton in Water Clusters: An ab Initio Molecular Dynamics Study,” Pranam Journal of Physics, Vol. 65, No. 4, 2005, pp. 763-768.
- M. Wikström, M. I. Verkhosky and G Hummer, “Water-Gated Mechanism of Proton Translocation by Cytochrome C Oxidase,” Biochimica & Biophysica Acta, Bioenergetics, Vol. 1604, No. 2-5, 2003, pp. 61-65.
- P. Mentré, “Interfacial Water: A Modulator of Biological Activity,” Journal of Biological Physics and Chemistry, Vol. 4, 2004, pp. 115-123.
- M. de Mattei, M. Fini, S. Setti, A. Ongaro, D. Gemmati, G. Stabellini, A. Pellati and A. Caruso, “Proteoglycan Synthesis in Bovine-Articular Cartilage Explants Exposed to Different Low-Frequency Low-Energy Pulsed Electromagnetic Fields,” Osteoarthritis and Cartilage, Vol. 15, No. 2, 2007, pp. 163-168.
- F. Pezetti, M. De Mattei, A. Caruso, R. Cadossi, P. Zucchini, F. Carinci, G. C. Traina and V. Sollazo, “Effects of Pulsed Electromagnetic Fields on Human Chondrocytes, an in-Vitro Study,” Calcified Tissue International, Vol. 65, No. 5, 1999, pp. 396-401.doi:10.1007/s002239900720
- M. De Mattei, A. Pellati, M. Pasello, A. Ongaro, S. Setti, L. Massari, et al., “Effects of Physical Stimulation With Electromagnetic Field and Insulin Growth Factor-1 Treatment on Proteoglycan Synthesis of Bovine Articular Carilage,” Osteoarthritis and Cartilage, Vol. 12, No. 10, 2004, pp. 793-800.
- M. De Mattei, A. Caruso, F. Pezetti, A. Pellati, G. Stabellini and V. Sollazo, “Effects of Pulsed Electromagnetic Fields on Human Articular Chondrocyte Proliferation,” Connective Tissue Research, Vol. 42, No. 2, 2001, pp. 1- 11.
- M. De Mattei, A. Caruso, F. Pezetti, A. Pellati, G. Stabellini, V. Sollazo and G. C. Traina, “Electromagnetic Fields on Human Articular Chondrocyte Proliferation,” Connective Tissue Research, Vol. 42, No. 4, 2001, pp. 269-279.
- M. E. Jahns, E. Lou, N. G. Durdie, et al., “The Effect of Pulsed Electromagnetic Fields on Chondrocyte Morphology,” Medical & Biological Engineering & Computing, Vol. 45 , No. 10, 2007, pp. 917-925.
- R. Cadossi, F. Bersani, A. Cossarizza, P. Zucchini, G. Emilia, G. Torelli and C Francheschi, “Lymphocytes and Low-Frequency Electromagnetic Fields,” The FASEB Journal, Vol. 6, No. 9, 1992, pp. 2667-2675.
- J. Zhou, G. Gao, J. Zhang and Z. Chang, “CREB DNA Binding Activation by a 50-Hz Magnetic Field in HL-60 Cells is Dependent on Extraand intracellular Ca2+ but Not PKA, PKC, ERK or p38 MAPK,” Biochemical and Biophysical Research Communications, Vol. 296, No. 4, 2002, pp. 1013-1018. doi:10.1016/S0006-291X(02)02022-3
- S. Mac Neil, R. A. Dawson, et al., “Extracellular CalmoDulin and Its Association with Epidermal Growth Factor in Normal Human Body Fluids,” Journal of Endocrinology, Vol. 118, No. 3, 1988, pp. 501-509.
- T. WenQiang, G. Yi, S. Yu, T. Jung and S. DaYe, “Extracellular Calmodulin-Binding Proteins in Body Fluids of Animals,” Journal of Endocrinology, Vol. 155, No. 1, 1997, pp. 13-17.
- E. Onuma and S. Hu, “Electric-Field-Directed Cell Shape Changes, Displacement and Cytoskeletal Reorganization Are Calcium Dependent,” The Journal of Cell Biology, Vol. 106, No. 6, 1988, pp. 2067-2075.
- M. Cho, I. Titushkin and S. Shun, “Altered Calcium Dynamics and Cellular Mechanisms Mediate Electrically Enhanced Stem Cell Differentiation, Session 5, Mechanisms of Cell Interactions with EMF,” Proceedings of the 29th Biolectromagnetic Society Annual Meeting, Kanazawa, 10-15 June 2007, pp. 84-85.
- A. Schuster, “Intracellular Calcium and Diameter Dynamics in Arteries Perfused in Vitro,” Thèse de Doctorat de Physique, EPFL, Lausanne, 2001.
- J. M. Bathon, “Bradikinine Is a Potent and Relatively Selective Stimulus for Cytosolic Calcium Elevation In Human Synovial Cells,” The Journal of Immunology, Vol. 153, No. 6, 1994, pp. 2600-2608.
- D. Gelperin, “Bradikinine (Bk) Increases Cytosolic Calcium in Cultured Rat Myenteric Neurons,” Pharmacology, Vol. 271, No. 1, 1994, pp. 507-514.
- N. Voitenko, “Calcium Disorders as A Common Denominator of Different Types of Pain,” Physiology News, Vol. 65, 2006, pp. 33-34. http://www.physoc.org
- S. Hanada and S. Yamada, “Extremely Low Frequency Magnetic Fields Affect Transcript Levels os Neuronal Genes in Caenorhabditis Elegans,” Proceedings of BEMS the 29th Annual Meeting, Kanazawa, 10-15 June 2007, pp. 436-437.
- L. Potenza, L. Ubaldi, R. De Sanctis, R. de Bellis, R. Cucchianini and M. Dacha, “Effects of a Static Magnetic Field on Cell Growth and Gene Expression in Escherichia Coli,” Mutation Research, Vol. 561, No. 1-2, 11 July 2005, pp. 53-62.
- Y. Hashimoto, “Effect of Static Magnetic Field on Cell Migration,” Electrical Engineering in Japan, Vol. 160, No. 2, 2007, pp. 46-52. doi:10.1002/eej.20203
- S. Xu, N. Tomita, K. Ikeuchi and Y. Ikada, “Recovery of Small-Sized Blood Vessels in Ischemic Bone under Static Magnetic Field,” ECAM, Vol. 4, No. 1, 2007, pp. 59-63.
- H. Okano, K. Gmitrov and C. Ohkubo, “Biphasic Effects of Static Magnetic Fields on Cutaneous Microcirculation in Rabbits,” Bioelectromagnetics, Vol. 20, No. 3, 1999, pp. 161-171.
- H. N. Mayrowitz and E. E. Groseclose, “Effects of a Static Magnetic Field of Either Polarity on Skin Microcirculation,” Microvascular Research, Vol. 69, No. 1-2, 2005, pp. 24-27. doi:10.1016/j.mvr.2004.11.002
- P. Le Chapellier and B. Matta, “Pain Relief by Magnetic Fields and Theories of Radicals Pair and Methionine Oxidation,” Procedeings of the 3rd International Forum on Pain Medicine, Montreal, 28 June-1 July 2007, p. 128.
- J. M. P. Le Chapellier, “Les eaux de l’âme et de l’Esprit, Journées Toulousaines de l’Eau 2010,” Institut Catholique de Toulouse, Toulouse, 11-12 Septembre 2010, (in French).
- C. G. Jung, “Mysterium Conjonctionis,” T1, Albin Michel, Paris, (in French).
- J. Dieudonné, “Eléments d’Analyse,” Vol. IX, Topologie Algébrique et Différentielle, Gauthier-Villars, Paris, 1982, (in French).
- J. Bard, “The Cellular and Molecular Processes of Developmental Anatomy,” In: C H. Waddington, Ed., Biological Development, Enc. Britannica, Chicago, 2007.
- R. M. Kiehn “Non Equilibrium Systems and Irreversible Processes,” Non Equilibrium Thermodynamics, Lulu, Vol. 1, 2009. http://www.lulu.com/kiehn
- R. M. Kiehn, “Kolmogorov-Cartan T0 Spaces of Exterior Differential Forms with Applications to Non-Equilibrium Thermodynamic Systems, and the Emergence of Morphogenic Germs via Dissipative Irreversible Processes,” International Conference on Topology and its Applications, Ankara, 6-11 July 2009.
- R. M. Kiehn, “Private Communication to the Authors,” February 2010.
- R. M. Kiehn “T0 or Not-T0, That Is the Topological Question, Algebra meets Topology: Advances and Applications,” A Conference in Honour of Dikran Dikranjan’s 60th Birthday, UPC-Barcelona Tech., Barcelona, 19-23 July 2010.
NOTES
14T = 4 Teslas. It is a value of magnetic field induction, 80 000 times the average value of the present earth magnetic field induction.
2The aporia is an insurmountable logical difficulty. The aporia of Cartesian dualism is beyond the fact that the Cartesian operation has a residue which is that the human body matter makes exception and presents a body-mind interaction:
3The Cartesian dualism aporia stems from absurdity that, in natural perception, sense order be dissociated from causal order. It is absurd that sense order be ignored in the comprehension of the biological process and consequently excluded from scientific construction. 3The cybernetic aporia stems from absurdity that an abstraction representing a functional diagram often buckled, i.e. implying a feedback going from end towards beginning of a process in the living matter, can emerge from a substrate made up of simple biochemical molecules aggregate. A buckling factor is needed.
4Aqueous physical vacuum extends in liquid water between H2O molecules always separated with a minimum vacuum distance d, around 2 Ä. Forces structured it.
5Topology was first a qualitative geometry. It applies to an object properties which are retained under deformation, so as a triangle be topologically equivalent to a circle.
6Life, according to Philosophy of Nature, is on the contrary regarded as a structure or an envelope for microphenomenons. According to Toscano, (2003, [10]), life of unicellular or complex organisms is marked as being a relational and unifying total structure, (an architectonic.) This envelope-structure sets constraints on smaller scale phenomenons, such as those relating to biomolecules and their biochemical reactions. That “enveloping” which is associated to organic life, gives rise to the existence of bio-regulation models.
7Ontology studies the fundamental characters of any reality as such.
8The Holism is sophisticated cybernetic doctrines which consider that the determining biological factors of an organism are its irreducible totalities: Dependent on organization levels, these totalities justify the cybernetic mechanisms of control and regulation. However the holism doctrine considers that totalities of an organic nature are made up of a non causal principle which connects and assembles its parts.
9The delocalized word has here with the chemical significance of: stripped of location according to a simple location with 4 dimensions, but however present within totality.
10The first law of thermodynamics is the law of energy E conservation, through an equivalence between heat Q and work W, according to: W + dE = Q.
11The Aharonov-Bohm effect is that the phase coherence of electrons is changed by a magnetic A or electrostatic Φ potential, as indicated by Van Oudennaarden, (1998, [40]).
12Surface tension is a property of a liquid surface to be like a stretched membrane. It results that a water drop tends to take a spherical form, in order to minimize its surface. This tension ensures a rise of water in capillary tubes, of whose height is a function opposite to the diameter of the capillary tube, as following the Jurin law.
13The bosons are subatomic particles with a whole spin, (it is to say with an angular momentum noted 0, 1, etc,) controled by the BoseEinstein statistics. Bosons embody the force fields; for example, the photons transport the energy of EM waves.
14The wild ground worm lives in a natural geomagnetism whose force of induction, or intensity, is about 0, 5 Gauss, - and comprises nuances and fluctuations, useful to its existence in the world.
15Pain relief by SMF: In vivo try with laboratory animals or human patients showed the antinociceptive effect is quite frequent but not systematic, probably because cells sense order can resound also on other unknown events.